
Effect of Biostimulant Application on Plant Growth, Chlorophylls and Hydrophilic Antioxidant Activity of Spinach (Spinacia oleracea L.) Grown under Saline Stress

 ,
,  , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Crop Management, Saline Irrigation and Biostimulant Application
2.2. Soil Electrical Conductivity Measurements, and Temperature Monitoring
2.3. Yield, Growth Parameters Measurements, and Nitrate Determination
2.4. SPAD Index, and Leaves Color Determination
2.5. Determination of Chlorophylls, Carotenoids, and Antioxidant Compounds and Activity
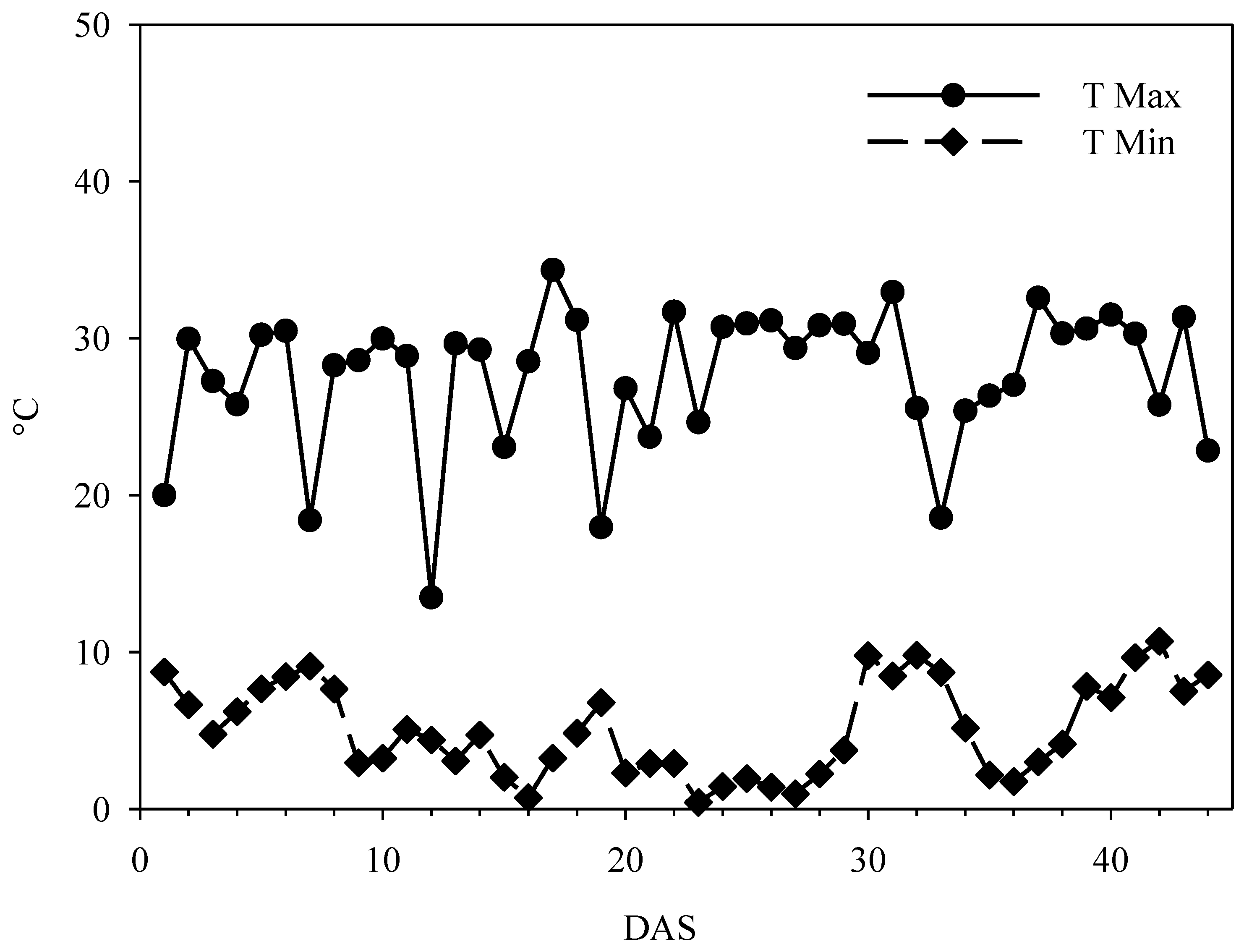
2.6. Climate Characteristics of the Greenhouse
2.7. Statistical Analysis
3. Results and Discussion
3.1. Electrical Conductivity of Soil
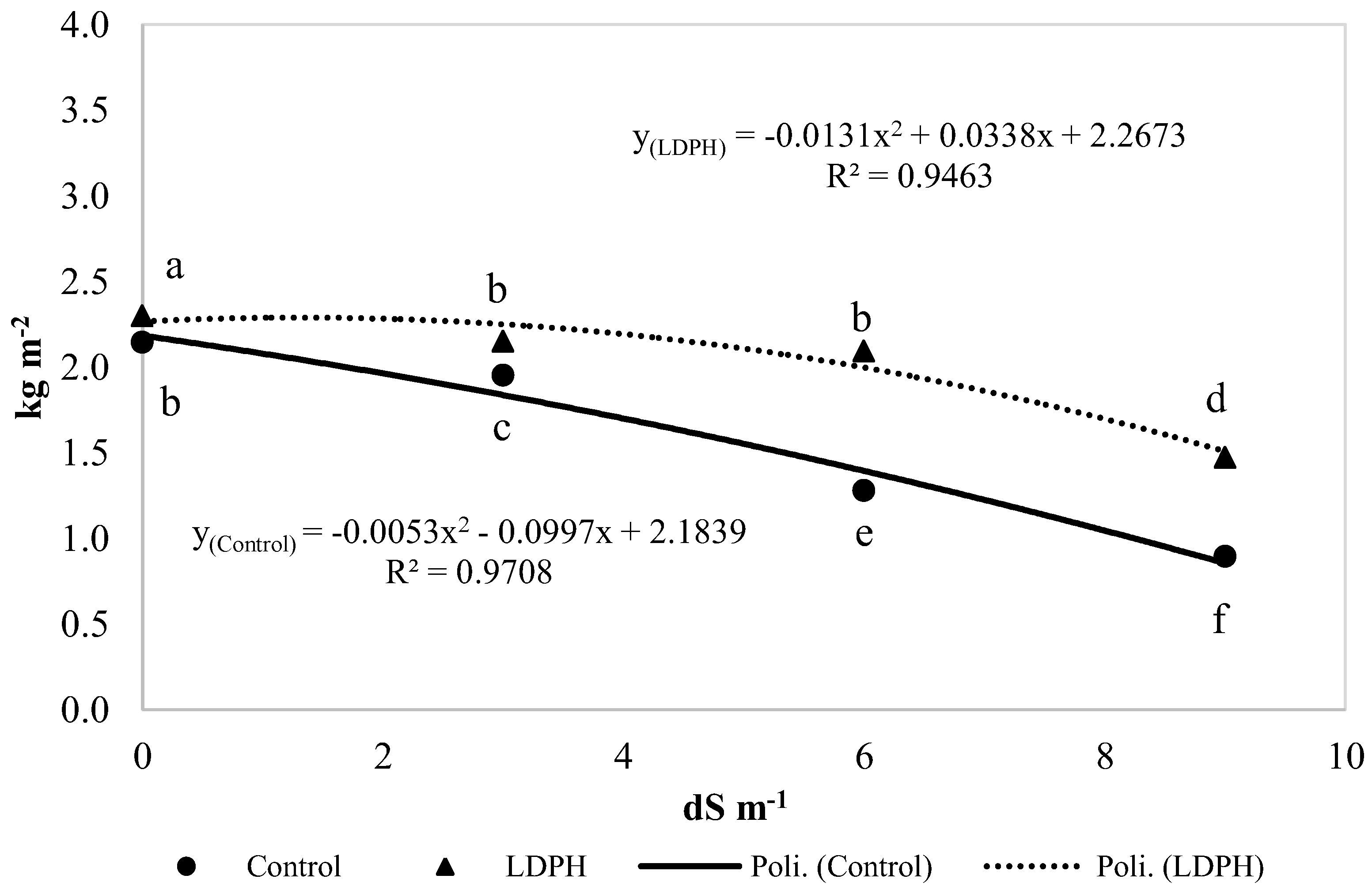
3.2. Yield and Growth Parameters
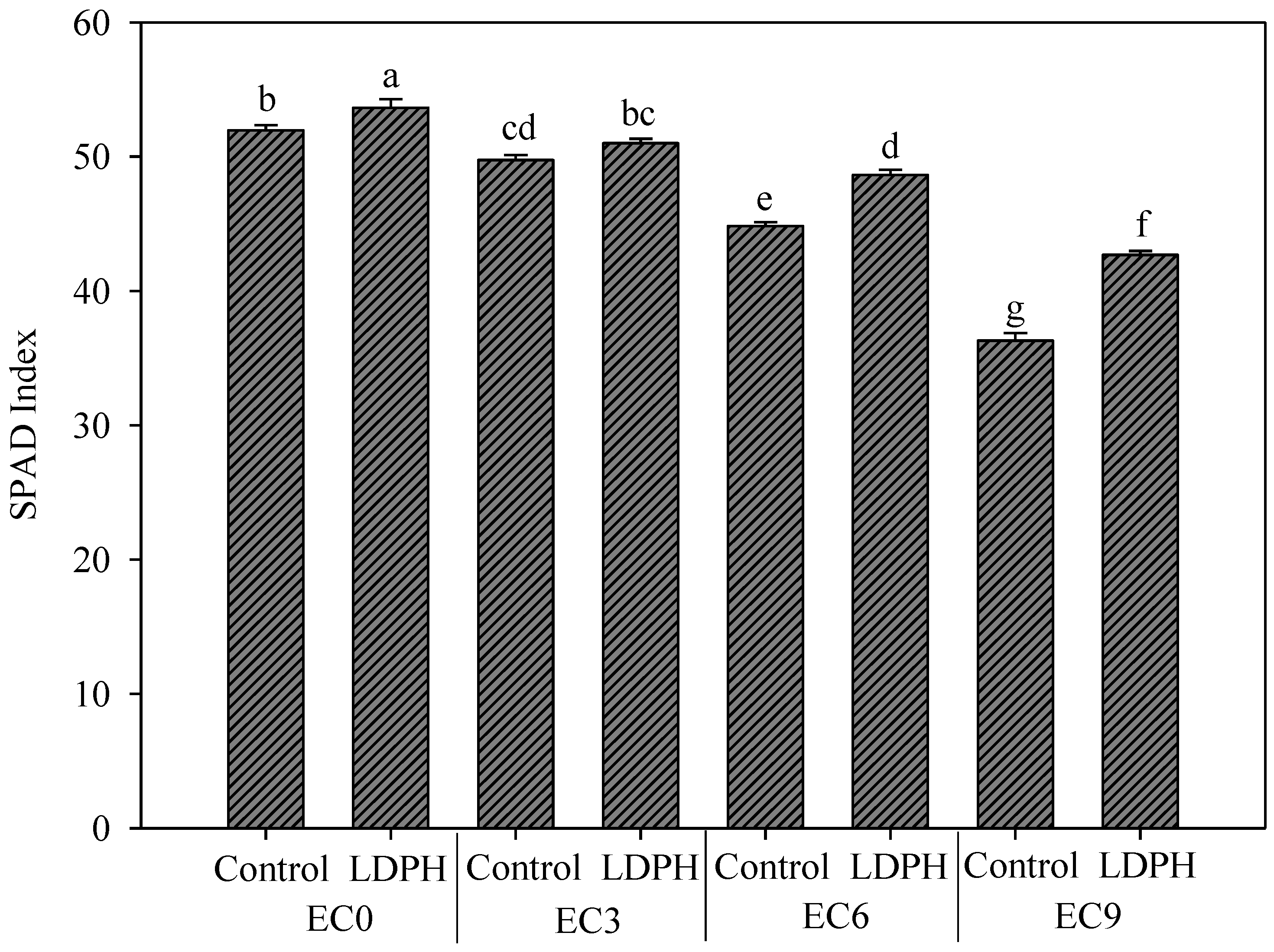
3.3. SPAD Index and Color Parameters
3.4. Chlorophylls, Carotenoids, and Nitrate Content of Spinach Leaves
3.5. Antioxidant Compounds and Activity of Spinach Leaves
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, H.J.; Fonseca, J.M.; Choi, J.H.; Kubota, C.; Dae, Y.K. Salt in irrigation water affects the nutritional and visual properties of romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Zaman, M.; Shahid, S.A.; Heng, L. Introduction to Soil Salinity, Sodicity and Diagnostics Techniques; Springer: Cham, Switzerland, 2018; ISBN 9783319961897. [Google Scholar]
- Ferreira, J.F.S.; Sandhu, D.; Liu, X.; Halvorson, J.J. Spinach (Spinacea oleracea L.) response to salinity: Nutritional value, physiological parameters, antioxidant capacity, and gene expression. Agriculture 2018, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Caparrotta, S.; Masi, E.; Atzori, G.; Diamanti, I.; Azzarello, E.; Mancuso, S.; Pandolfi, C. Growing spinach (Spinacia oleracea) with different seawater concentrations: Effects on fresh, boiled and steamed leaves. Sci. Hortic. 2019, 256, 108540. [Google Scholar] [CrossRef]
- Rouphael, Y.; Carillo, P.; Garcia-Perez, P.; Cardarelli, M.; Senizza, B.; Miras-Moreno, B.; Colla, G.; Lucini, L. Plant biostimulants from seaweeds or vegetal proteins enhance the salinity tolerance in greenhouse lettuce by modulating plant metabolism in a distinctive manner. Sci. Hortic. 2022, 305, 111368. [Google Scholar] [CrossRef]
- Shannon, M.C.; Grieve, C.M. Tolerance of vegetable crops to salinity. HortScience 1999, 78, 5–38. [Google Scholar] [CrossRef]
- Turhan, A.; Kuscu, H.; Ozmen, N.; Asik, B.B.; Serbeci, M.S.; Seniz, V. Alleviation of deleterious effects of salt stress by applications of supplementary potassium-calcium on spinach. Acta Agric. Scand. Sect. B Soil Plant Sci. 2013, 63, 184–192. [Google Scholar] [CrossRef]
- Lim, J.H.; Park, K.J.; Kim, B.K.; Jeong, J.W.; Kim, H.J. Effect of salinity stress on phenolic compounds and carotenoids in buckwheat (Fagopyrum esculentum M.) sprout. Food Chem. 2012, 135, 1065–1070. [Google Scholar] [CrossRef]
- Bonasia, A.; Conversa, G.; Lazzizera, C.; Elia, A. Foliar application of protein hydrolysates on baby-leaf spinach grown at different n levels. Agronomy 2022, 12, 36. [Google Scholar] [CrossRef]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; De Pasquale, C.; Iapichino, G.; D’anna, F.; La Bella, S. Protein hydrolysates and mo-biofortification interactively modulate plant performance and quality of ‘canasta’ lettuce grown in a protected environment. Agronomy 2021, 11, 1023. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Tsouvaltzis, P.; Koukounaras, A.; Siomos, A.S. Application of amino acids improves lettuce crop uniformity and inhibits nitrate accumulation induced by the supplemental inorganic nitrogen fertilization. Int. J. Agric. Biol. 2014, 16, 951–955. [Google Scholar]
- Srivastava, N. Biostimulants for Plant Abiotic Stress Tolerance. In Biostimulants for Crop Production and Sustainable Agriculture; CABI: Wallingford, UK, 2022. [Google Scholar]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristofano, F.; El-Nakhel, C.; Rouphael, Y. Biostimulant substances for sustainable agriculture: Origin, operating mechanisms and effects on cucurbits, leafy greens, and nightshade vegetables species. Biomolecules 2021, 11, 1103. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Kumar, P.; Colla, G. The effect of a plant-derived biostimulant on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Kunicki, E.; Grabowska, A.; Sękara, A.; Wojciechowska, R. The effect of cultivar type, time of cultivation, and biostimulant treatment on the yield of spinach (Spinacia oleracea L.). Folia Hortic. 2010, 22, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, G.; Samani, Z. Reference crop evapotranspiration from temperature. Appl. Eng. Agric. 1985, 1, 96–99. [Google Scholar] [CrossRef]
- Di Mola, I.; Conti, S.; Cozzolino, E.; Melchionna, G.; Ottaiano, L.; Testa, A.; Sabatino, L.; Rouphael, Y.; Mori, M. Plant-based protein hydrolysate improves salinity tolerance in hemp: Agronomical and physiological aspects. Agronomy 2021, 11, 342. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; El-Nakhel, C.; Rippa, M.; Mormile, P.; Corrado, G.; Rouphael, Y.; Mori, M. Assessment of Yield and Nitrate Content of Wall Rocket Grown under Diffuse-Light-or Clear-Plastic Films and Subjected to Different Nitrogen Fertilization Levels and Biostimulant Application. Horticulturae 2022, 8, 138. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteau Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Louati, D.; Majdoub, R.; Rigane, H.; Abida, H. Effects of Irrigating with Saline Water on Soil Salinization (Eastern Tunisia). Arab. J. Sci. Eng. 2018, 43, 3793–3805. [Google Scholar] [CrossRef]
- Ben Ahmed, C.; Magdich, S.; Ben Rouina, B.; Boukhris, M.; Ben Abdullah, F. Saline water irrigation effects on soil salinity distribution and some physiological responses of field grown Chemlali olive. J. Environ. Manag. 2012, 113, 538–544. [Google Scholar] [CrossRef]
- Kargas, G.; Londra, P.; Sgoubopoulou, A. Comparison of soil EC values from methods based on 1:1 and 1:5 soil to water ratios and ECe from saturated paste extract based method. Water 2020, 12, 1010. [Google Scholar] [CrossRef] [Green Version]
- Ors, S.; Suarez, D.L. Spinach biomass yield and physiological response to interactive salinity and water stress. Agric. Water Manag. 2017, 190, 31–41. [Google Scholar] [CrossRef]
- Yamada, M.; Kuroda, C.; Fujiyama, H. Growth promotion by sodium in amaranthaceous plants. J. Plant Nutr. 2016, 39, 1186–1193. [Google Scholar] [CrossRef]
- Ors, S.; Suarez, D.L. Salt tolerance of spinach as related to seasonal climate. Hortic. Sci. 2016, 43, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Dias, M.I.; Petropoulos, S.A.; Plexida, S.; Chrysargyris, A.; Tzortzakis, N.; Calhelha, R.C.; Ivanov, M.; Stojković, D.; Soković, M.; et al. The effects of biostimulants, biofertilizers and water-stress on nutritional value and chemical composition of two spinach genotypes (Spinacia oleracea L.). Molecules 2019, 24, 4494. [Google Scholar] [CrossRef] [PubMed]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Giordano, M.; Rouphael, Y.; Colla, G.; Mori, M. Effect of Vegetal- and Seaweed Extract-Based Biostimulants on Agronomical and Leaf Quality Traits of Plastic Tunnel-Grown Baby Lettuce under Four Regimes of Nitrogen Fertilization. Agronomy 2019, 9, 571. [Google Scholar] [CrossRef] [Green Version]
- Goñi, O.; Quille, P.; Connell, S.O. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Taofiq, O.; Fernandes, Â.; Tzortzakis, N.; Ciric, A.; Sokovic, M.; Barros, L.; Ferreira, I.C.F.R.; Plexida, S.; Chrysargyris, A.; et al. Bioactive properties of greenhouse-cultivated green beans (Phaseolus vulgaris L.) under biostimulants and water-stress effect. J. Sci. Food Agric. 2019, 99, 6049–6059. [Google Scholar] [CrossRef] [Green Version]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Petropoulos, S.A. Biostimulants Application: A Low Input Cropping Management Tool for Sustainable Farming of Vegetables. Biomolecules 2021, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant-and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Nocerino, S.; Rouphael, Y.; Colla, G.; El-Nakhel, C.; Mori, M. Nitrogen use and uptake efficiency and crop performance of baby spinach (Spinacia oleracea L.) and Lamb’s Lettuce (Valerianella locusta L.) grown under variable sub-optimal N regimes combined with plant-based biostimulant application. Agronomy 2020, 10, 278. [Google Scholar] [CrossRef] [Green Version]
- Rahdari, P.; Tavakoli, S.; Hosseini, S.M. Studying of salinity stress effect on germination, proline, sugar, protein, lipid and chlorophyll content in purslane (Portulaca oleracea L.) leaves. J. Stress Physiol. Biochem. 2012, 8, 182–193. [Google Scholar]
- Corrado, G.; Vitaglione, P.; Giordano, M.; Raimondi, G.; Napolitano, F.; Di Stasio, E.; Di Mola, I.; Mori, M.; Rouphael, Y. Phytochemical responses to salt stress in red and green baby leaf lettuce (Lactuca sativa L.) varieties grown in a floating hydroponic module. Separations 2021, 8, 175. [Google Scholar] [CrossRef]
- Fallovo, C.; Rouphael, Y.; Rea, E.; Battistelli, A.; Colla, G. Nutrient solution concentration and growing season affect yield and quality of Lactuca sativa L. var. acephala in floating raft culture. J. Sci. Food Agric. 2009, 89, 1682–1689. [Google Scholar] [CrossRef]
- Xu, C.; Mou, B. Responses of spinach to salinity and nutrient deficiency in growth, physiology, and nutritional value. J. Am. Soc. Hortic. Sci. 2016, 141, 12–21. [Google Scholar] [CrossRef]
- Mostafa, H. Effects of salinity stress on growth, chlorophyll content and osmotic components of two basil (Ocimum basilicum L.) genotypes. Afr. J. Biotechnol. 2011, 11, 379–384. [Google Scholar] [CrossRef]
- Naz, R.; Zaman, Q.; Nazir, S.; Komal, N.; Chen, Y.; Ashraf, K.; Al-huqail, A.A.; Alfagham, A.; Id, M.H.S. Silicon fertilization counteracts salinity- induced damages associated with changes in physio-biochemical modulations in spinach. PLoS ONE 2022, 17, e0267939. [Google Scholar]
- Aktsoglou, D.C.; Kasampalis, D.S.; Sarrou, E.; Tsouvaltzis, P.; Chatzopoulou, P.; Martens, S.; Siomos, A.S. Protein hydrolysates supplement in the nutrient solution of soilless grown fresh peppermint and spearmint as a tool for improving product quality. Agronomy 2021, 11, 317. [Google Scholar] [CrossRef]
- Francesca, S.; Cirillo, V.; Raimondi, G.; Maggio, A.; Rigano, M.M.; Barone, A. A Novel Protein Hydrolysate-Based Biostimulant Improves Tomato Performances under Drought Stress. Plants 2021, 10, 783. [Google Scholar] [CrossRef]
- Bantis, F.; Fotelli, M.; Ilić, Z.S.; Koukounaras, A. Physiological and phytochemical responses of spinach baby leaves grown in a PFAL system with leds and saline nutrient solution. Agriculture 2020, 10, 574. [Google Scholar] [CrossRef]
- Szerement, J.; Szatanik, A.; Jakub, K.; Monika, M.; Hersztek, M. Agronomic Biofortification with Se, Zn, and Fe: An Effective Strategy to Enhance Crop Nutritional Quality and Stress Defense—A Review. J. Soil Sci. Plant Nutr. 2022, 22, 1129–1159. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Giordano, M.; Rouphael, Y.; El-Nakhel, C.; Leone, V.; Mori, M. Effect of seaweed (Ecklonia maxima) extract and legume-derived protein hydrolysate biostimulants on baby leaf lettuce grown on optimal doses of nitrogen under greenhouse conditions. Aust. J. Crop Sci. 2020, 14, 1456–1464. [Google Scholar] [CrossRef]
- Chung, J.B.; Jin, S.J.; Cho, H.J. Low water potential in saline soils enhances nitrate accumulation of lettuce. Commun. Soil Sci. Plant Anal. 2005, 36, 1773–1785. [Google Scholar] [CrossRef]
- Consentino, B.B.; Virga, G.; la Placa, G.G.; Sabatino, L.; Rouphael, Y.; Ntatsi, G.; Iapichino, G.; la Bella, S.; Mauro, R.P.; D’anna, F.; et al. Celery (Apium graveolens L.) performances as subjected to different sources of protein hydrolysates. Plants 2020, 9, 1633. [Google Scholar] [CrossRef]
- Sogoni, A.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. The impact of salt stress on plant growth, mineral composition, and antioxidant activity in Tetragonia decumbens Mill.: An underutilized edible halophyte in South Africa. Horticulturae 2021, 7, 140. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Dias, M.I.; Pereira, C.; Calhelha, R.C.; Chrysargyris, A.; Tzortzakis, N.; Ivanov, M.; Sokovic, M.D.; Barros, L.; et al. Chemical composition and plant growth of Centaurea raphanina subsp. mixta plants cultivated under saline conditions. Molecules 2020, 25, 2204. [Google Scholar] [CrossRef]
- Petropoulos, S.; Levizou, E.; Ntatsi, G.; Fernandes, Â.; Petrotos, K.; Akoumianakis, K.; Barros, L.; Ferreira, I. Salinity effect on nutritional value, chemical composition and bioactive compounds content of Cichorium spinosum L. Food Chem. 2017, 214, 129–136. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Electrical conductivity (dS m−1) | ||
|---|---|---|---|
| 11 February 2022 | 17 March 2022 | 30 March 2022 | |
| Water Salinity | |||
| EC0 | 0.38 ± 0.02 | 0.65 ± 0.06 c | 0.56 ± 0.07 c |
| EC3 | 0.38 ± 0.02 | 1.81 ± 0.19 b | 1.97 ± 0.07 b |
| EC6 | 0.38 ± 0.02 | 2.34 ± 0.21 ab | 2.71 ± 0.18 a |
| EC9 | 0.38 ± 0.02 | 2.83 ±0.18 a | 3.31 ±0.22 a |
| ns | *** | *** | |
| Treatments | Plants | Leaves | |||
|---|---|---|---|---|---|
| n° m−2 | n° m−2 | n° Plant−1 | ALW (g) | D.M. (%) | |
| Water Salinity | |||||
| EC0 | 1286.7 ± 99.0 a | 6952.9 ± 455.0 a | 5.4 ± 0.4 b | 0.32 ± 0.018 ab | 10.5 ± 0.3 |
| EC3 | 1048.3 ± 91.9 a | 5995.8 ± 393.4 b | 5.5 ± 0.5 b | 0.38 ± 0.021 a | 11.1 ± 0.6 |
| EC6 | 696.7 ± 69.5 b | 4802.3 ± 360.9 bc | 7.0 ± 0.5 a | 0.35 ± 0.023 a | 10.8 ± 0.1 |
| EC9 | 721.7 ±46.8 b | 3850.0 ± 217.9 c | 5.3 ± 0.2 b | 0.30 ± 0.016 b | 11.1 ± 0.4 |
| Biostimulant | |||||
| Control | 848.8 ± 82.5 b | 4640.1 ± 325.6 b | 5.98 ± 0.34 | 0.34 ± 0.016 | 11.1 ± 0.49 a |
| LDPH | 1027.9 ± 71.1 a | 6010.4 ± 388.0 a | 5.62 ± 0.45 | 0.34 ± 0.023 | 10.6 ± 0.19 b |
| Significance | |||||
| Salinity (S) | *** | *** | ** | ** | ns |
| Biostimulant (B) | ** | *** | ns | ns | * |
| S × B | ns | ns | ns | ns | ns |
| Water Salinity | Biostimulant | L* | a* | b* |
|---|---|---|---|---|
| EC0 | Control | 38.28 ± 0.91 bc | −12.17 ± 0.27 b | 18.42 ± 0.56 b |
| LDPH | 36.89 ± 0.50 c | −10.83 ± 0.39 a | 15.12 ± 0.81 c | |
| EC3 | Control | 39.03 ± 0.39 bc | −11.52 ± 0.32 ab | 16.37 ± 0.68 bc |
| LDPH | 38.68 ± 0.41 b | −11.81 ± 0.32 ab | 17.00 ± 0.67 bc | |
| EC6 | Control | 38.48 ± 0.38 bc | −11.19 ± 0.31 ab | 15.97 ± 0.53 c |
| LDPH | 38.59 ± 0.68 bc | −11.42 ± 0.44 ab | 16.92 ± 0.99 bc | |
| EC9 | Control | 43.08 ± 0.73 a | −14.41 ± 0.36 c | 21.93 ± 0.98 a |
| LDPH | 37.69 ± 0.42 bc | −11.38 ± 0.26 ab | 15.10 ± 0.52 c | |
| Significance | ||||
| Salinity (S) | *** | *** | ** | |
| Biostimulant (B) | *** | *** | *** | |
| S × B | *** | *** | *** |
| Treatments | Chlorophyll a | Chlorophyll b | Total Chlorophylls | Carotenoids | Nitrates |
|---|---|---|---|---|---|
| mg g−1 fw | mg g−1 fw | mg g−1 fw | mg g−1 fw | mg kg−1 fw | |
| Water Salinity | |||||
| EC0 | 1.163 ± 0.021 a | 0.743 ± 0.067 a | 1.907 ± 0.087 a | 0.321 ± 0.019 c | 2397.2 ± 225.0 a |
| EC3 | 1.157 ± 0.029 a | 0.680 ± 0.062 a | 1.838 ± 0.090 a | 0.340 ± 0.015 bc | 2389.4 ± 244.1 a |
| EC6 | 1.162 ± 0.019 a | 0.633 ± 0.042 ab | 1.795 ± 0.061 a | 0.360 ± 0. 010 ab | 2095.2 ± 187.5 a |
| EC9 | 1.038 ± 0.054 b | 0.469 ± 0.050 b | 1.507 ± 0.103 b | 0.391 ± 0.006 a | 256.9 ± 95.7 b |
| Biostimulant | |||||
| Control | 1.111 ± 0.321 | 0.614 ± 0.177 | 1.725 ± 0.498 b | 0.354 ± 0.102 | 1442.8 ± 244.7 b |
| LDPH | 1.115 ± 0.332 | 0.649 ± 0.187 | 1.798 ± 0.519 a | 0.352 ± 0.102 | 2126.6 ± 312.3 a |
| Significance | |||||
| Salinity (S) | *** | *** | ** | ** | *** |
| Biostimulant (B) | ns | ns | * | ns | *** |
| S × B | ns | ns | ns | ns | ns |
| Treatments | HAA | ABTS AA | Total Phenols | TAA |
|---|---|---|---|---|
| mmol Ascorbic Acid equ. 100 g−1 dw | Mmol Trolox equ. 100 g−1 dw | mg Gallic Acid g−1 dw | mg g−1 fw | |
| Water Salinity | ||||
| EC0 | 8.08 ± 0.30 a | 9.20 ± 0.99 c | 1.39 ± 0.04 c | 44.87 ± 7.52 |
| EC3 | 8.19 ± 0.38 a | 11.70 ± 0.55 b | 1.55 ± 0.08 bc | 49.57 ± 6.31 |
| EC6 | 8.16 ± 0.24 a | 14.33 ± 0.51 a | 1.77 ± 0.06 ab | 52.50 ± 4.11 |
| EC9 | 5.21 ± 0.62 b | 15.04 ± 0.67 a | 1.89 ± 0.14 a | 58.29 ± 7.33 |
| Biostimulant | ||||
| Control | 7.08 ± 0.58 b | 13.75 ± 0.68 a | 1.77 ± 0.09 a | 58.17 ± 3.32 a |
| LDPH | 7.74 ±0.30 a | 11.38 ± 0.84 b | 1.53 ± 0.06 b | 44.44 ± 4.77 b |
| Significance | ||||
| Salinity (S) | *** | *** | ** | ns |
| Biostimulant (B) | * | *** | * | * |
| S × B | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Nakhel, C.; Cozzolino, E.; Ottaiano, L.; Petropoulos, S.A.; Nocerino, S.; Pelosi, M.E.; Rouphael, Y.; Mori, M.; Di Mola, I. Effect of Biostimulant Application on Plant Growth, Chlorophylls and Hydrophilic Antioxidant Activity of Spinach (Spinacia oleracea L.) Grown under Saline Stress. Horticulturae 2022, 8, 971. https://doi.org/10.3390/horticulturae8100971
El-Nakhel C, Cozzolino E, Ottaiano L, Petropoulos SA, Nocerino S, Pelosi ME, Rouphael Y, Mori M, Di Mola I. Effect of Biostimulant Application on Plant Growth, Chlorophylls and Hydrophilic Antioxidant Activity of Spinach (Spinacia oleracea L.) Grown under Saline Stress. Horticulturae. 2022; 8(10):971. https://doi.org/10.3390/horticulturae8100971
Chicago/Turabian StyleEl-Nakhel, Christophe, Eugenio Cozzolino, Lucia Ottaiano, Spyridon A. Petropoulos, Sabrina Nocerino, Maria Eleonora Pelosi, Youssef Rouphael, Mauro Mori, and Ida Di Mola. 2022. "Effect of Biostimulant Application on Plant Growth, Chlorophylls and Hydrophilic Antioxidant Activity of Spinach (Spinacia oleracea L.) Grown under Saline Stress" Horticulturae 8, no. 10: 971. https://doi.org/10.3390/horticulturae8100971
APA StyleEl-Nakhel, C., Cozzolino, E., Ottaiano, L., Petropoulos, S. A., Nocerino, S., Pelosi, M. E., Rouphael, Y., Mori, M., & Di Mola, I. (2022). Effect of Biostimulant Application on Plant Growth, Chlorophylls and Hydrophilic Antioxidant Activity of Spinach (Spinacia oleracea L.) Grown under Saline Stress. Horticulturae, 8(10), 971. https://doi.org/10.3390/horticulturae8100971