
Analyses of Pepper Cinnamoyl-CoA Reductase Gene Family and Cloning of CcCCR1/2 and Their Function Identification in the Formation of Pungency


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Reagents
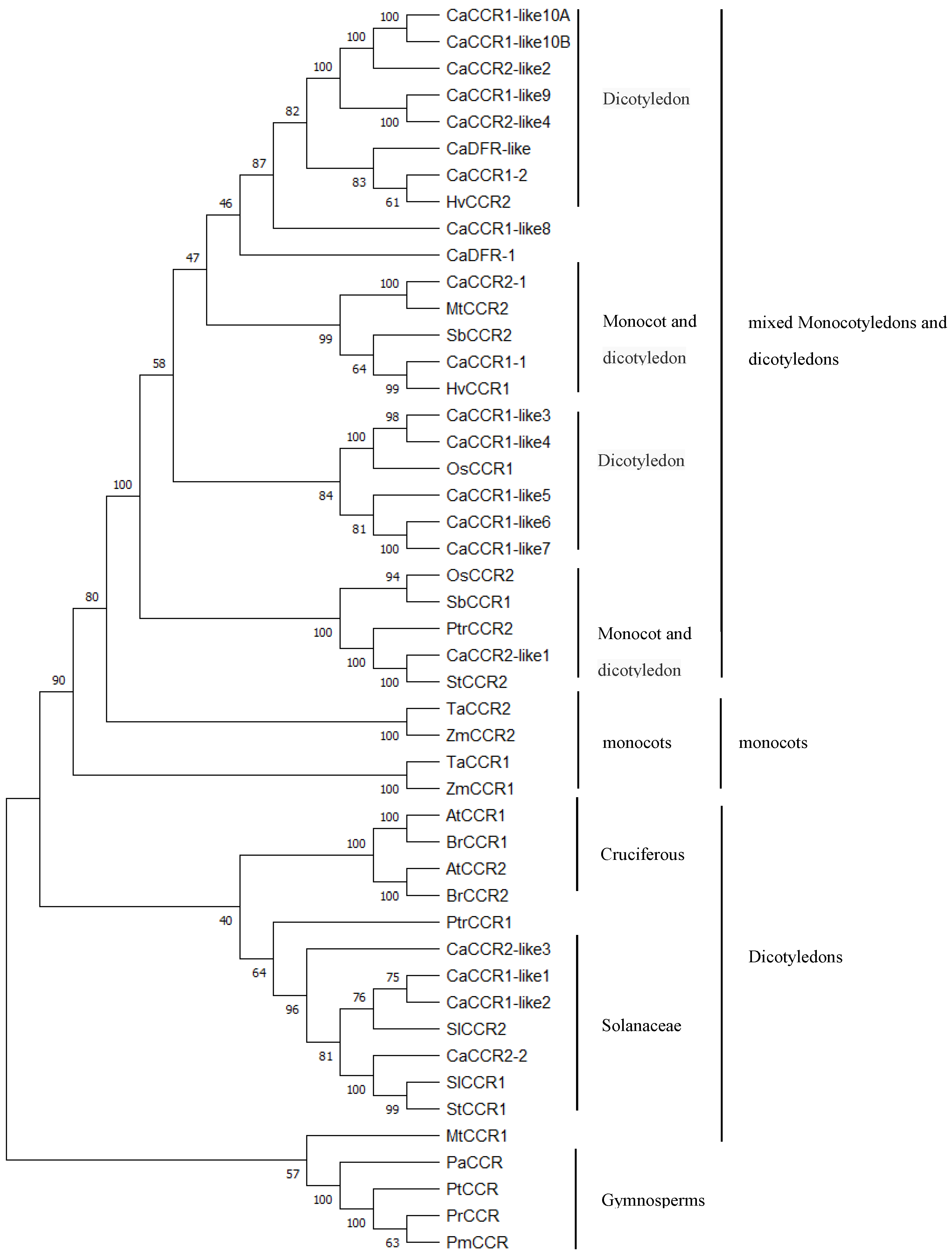
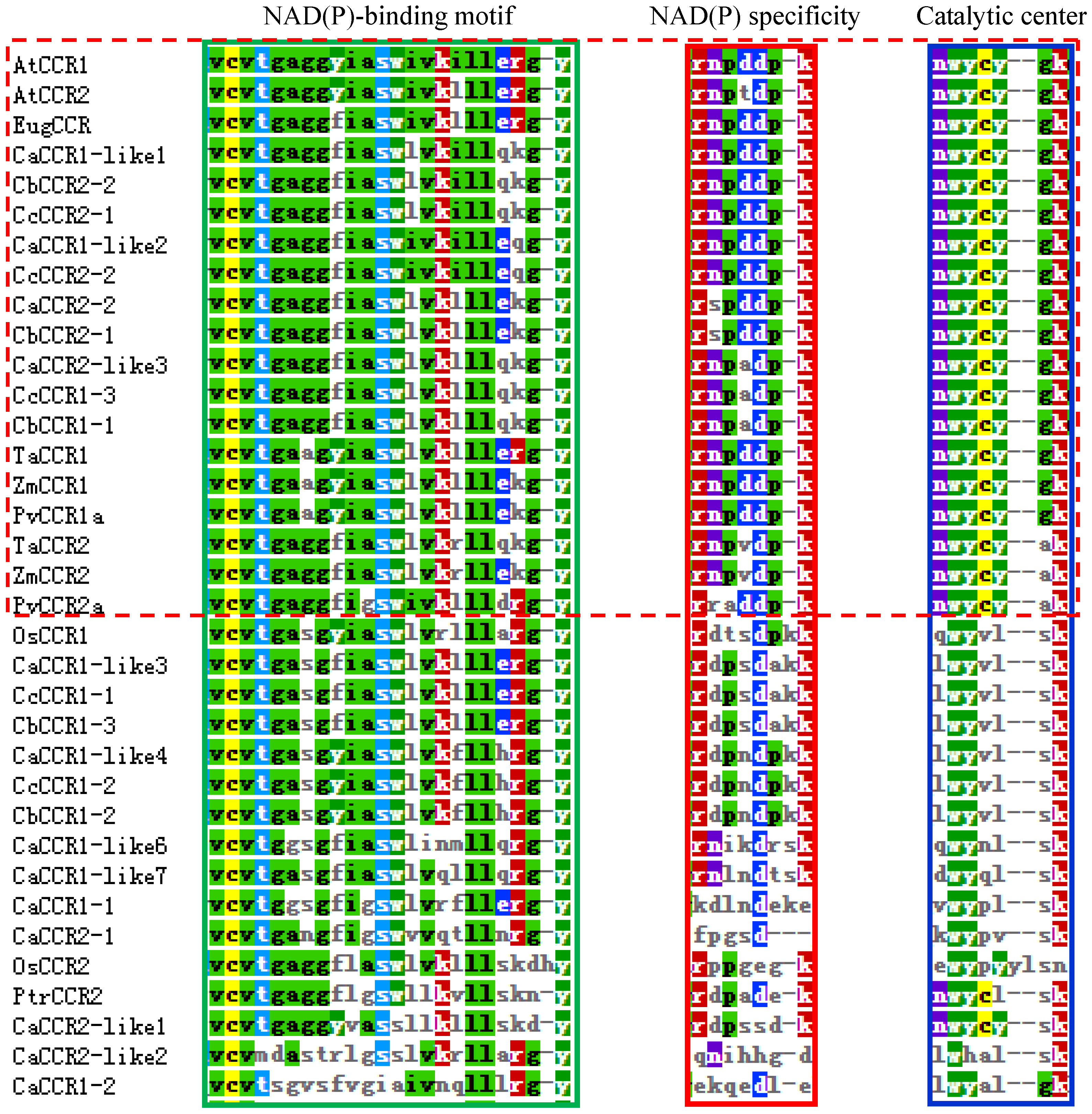
2.2. Multiple Sequence Alignments, Phylogenetic Analysis of Pepper CCRs and CCR-like Gene Family
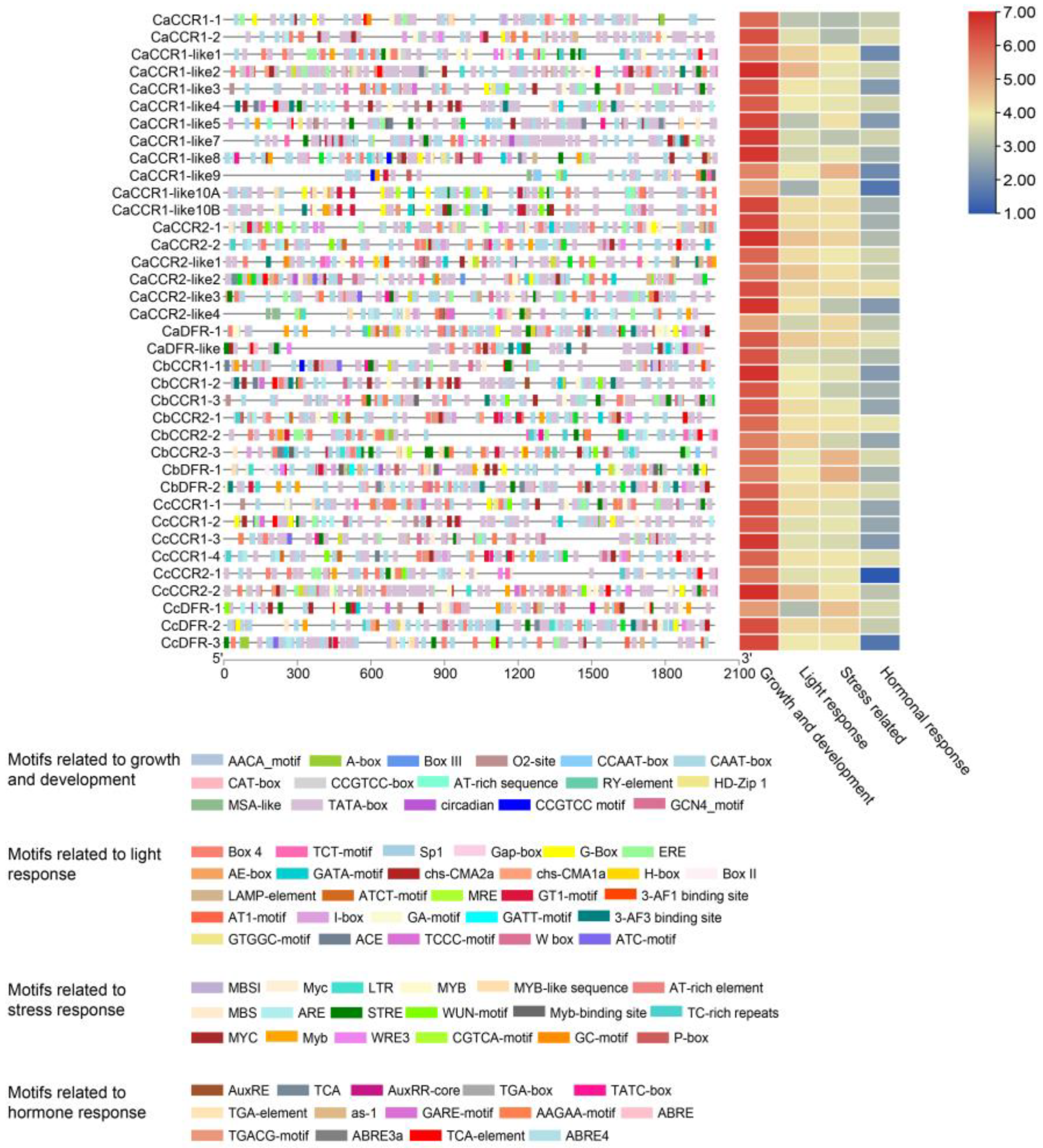
2.3. Analysis of Cis-Regulatory Elements in Pepper CCR and CCR-like Gene Family
2.4. Cloning and Bioinformatics Analysis of CcCCR1 and CcCCR2 from Huangdenglong Pepper
2.5. Expression of CCR1 and CCR2 in E. coli and Their Enzyme Kinetics Parameters
2.6. Heterologous Expression of CcCCR1 and CcCCR2 Genes in Arabidopsis thaliana
2.7. Virus-Induced Gene Silencing of CcCCR1 and CcCCR2 in Hainan Huangdenglong Pepper
2.8. Determination of Relative Expression of CCR Genes and Other Genes Involving in Capsaicin Biosynthesis
2.9. Determination of Capsaicin, Lignin, and Flavonoids in Peppers
3. Results
3.1. The CCR Gene Family and Their Intron–Exon Structures in Capsicum L.
3.2. Analysis of Cis-Regulatory Elements in Pepper CCR and CCR-like Gene Family
3.3. Sequence Homology and Phylogenetic Analysis of Pepper CCRs and CCR-like Protein Family
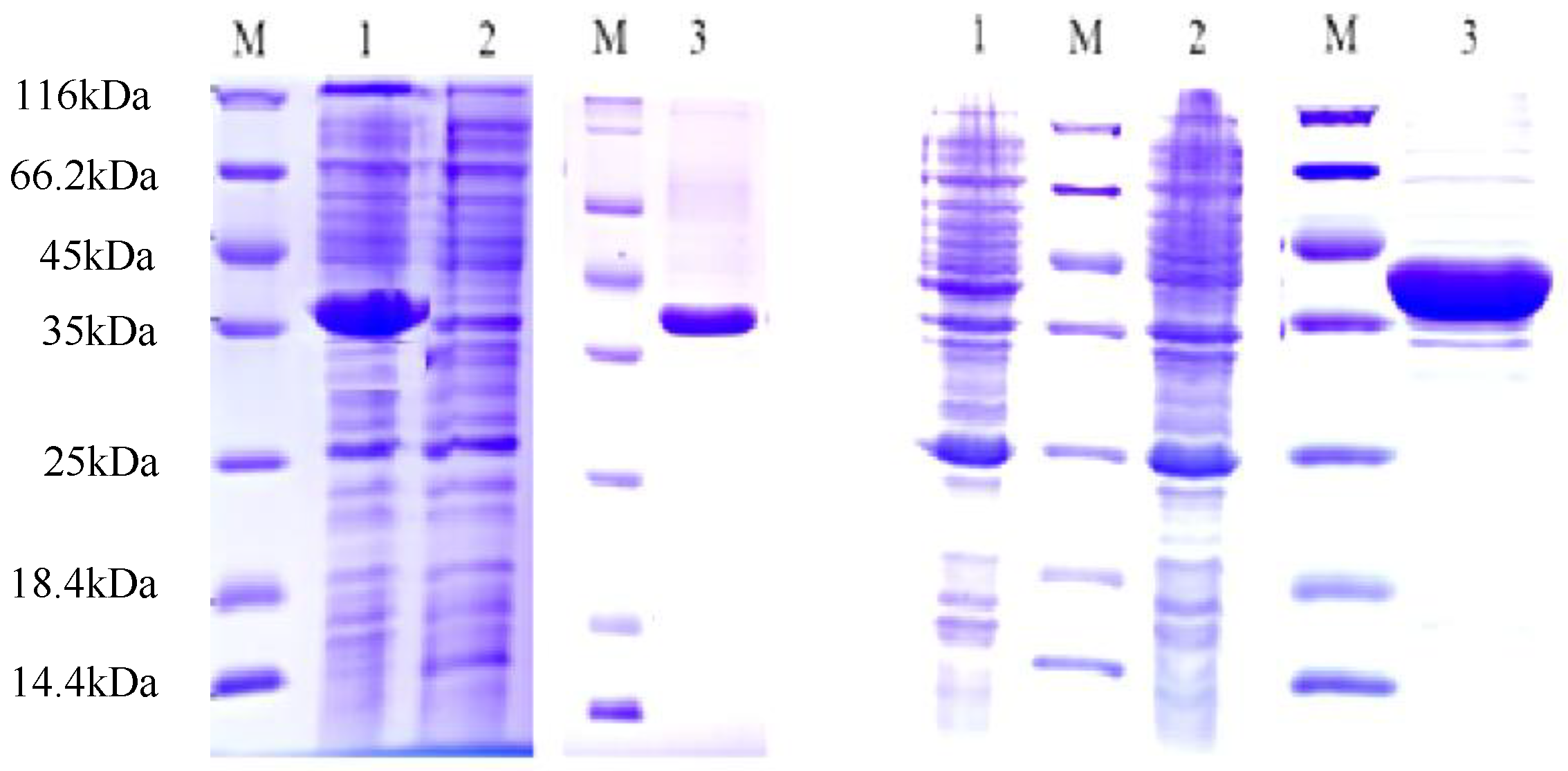
3.4. Cloning and Heterologous Expression of Two CCR Genes from Capsicum chinense
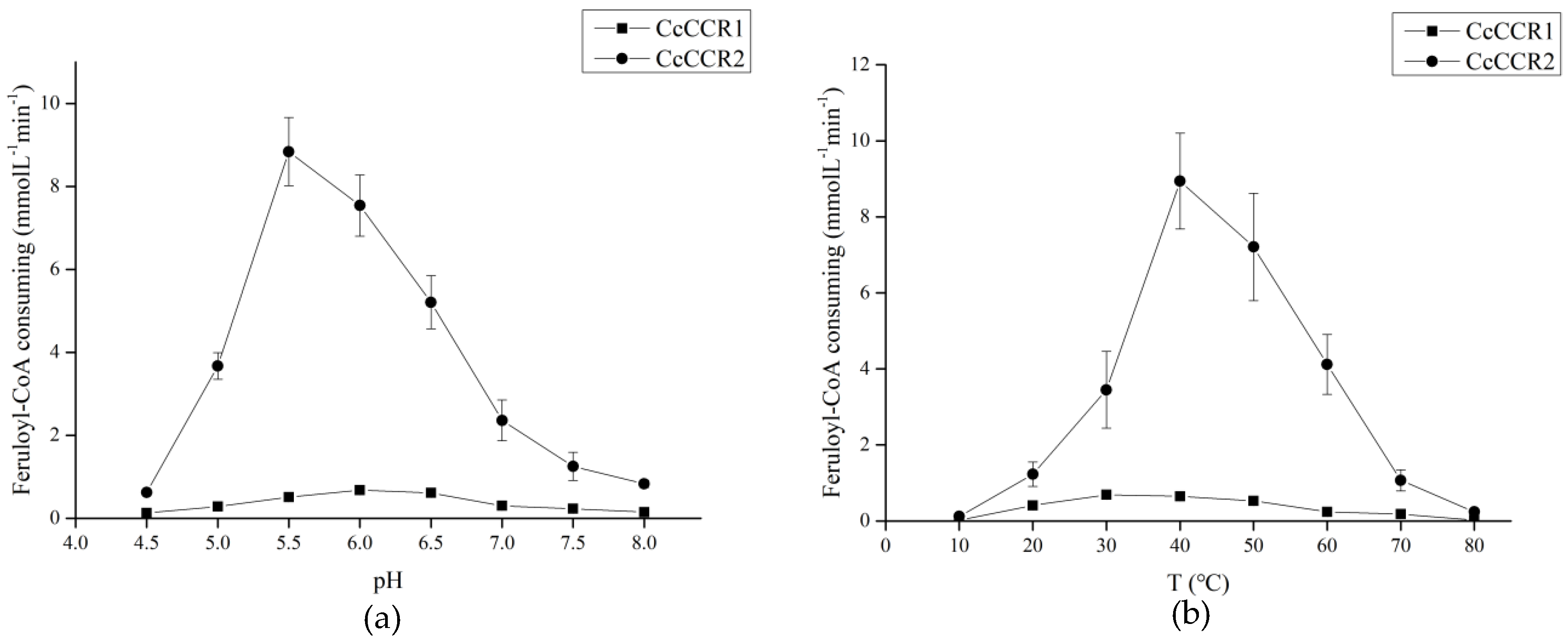
3.5. CCR Activity and Kinetic Parameters of the Recombinant CcCCR1 and CcCCR2
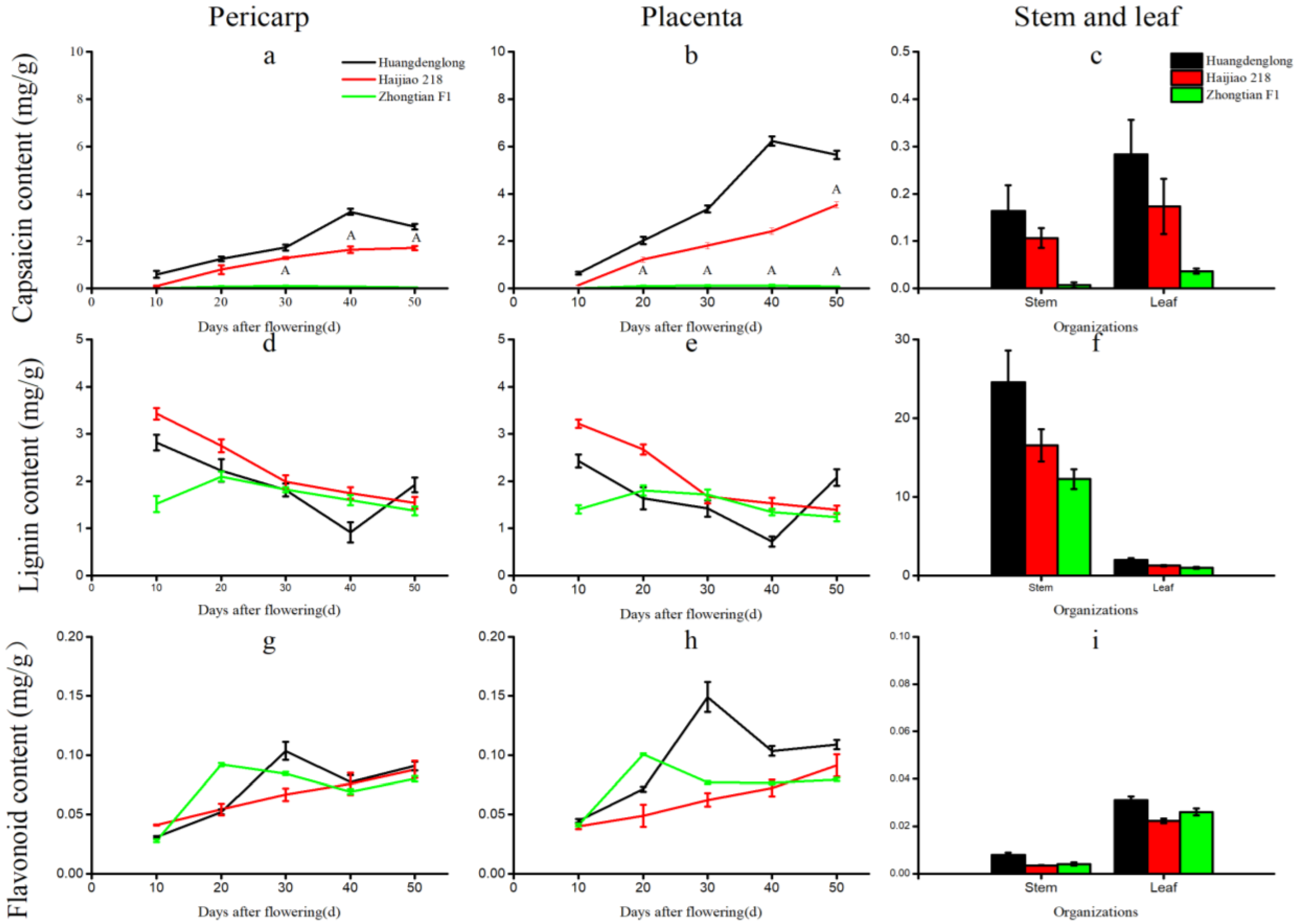
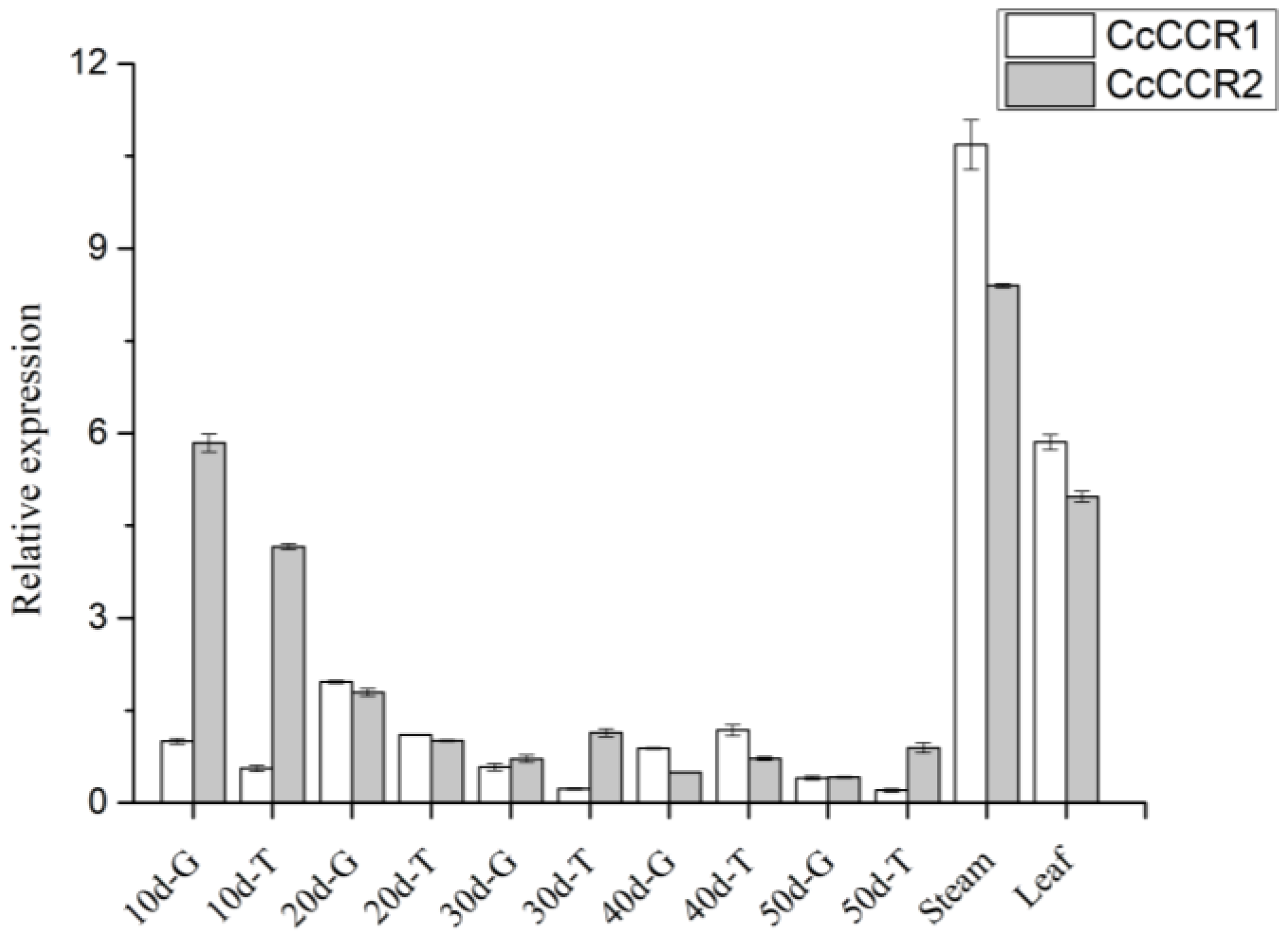
3.6. The Correlation between Expression of CcCCRs and Contents of Capsaicin, Lignin, and Flavonoids
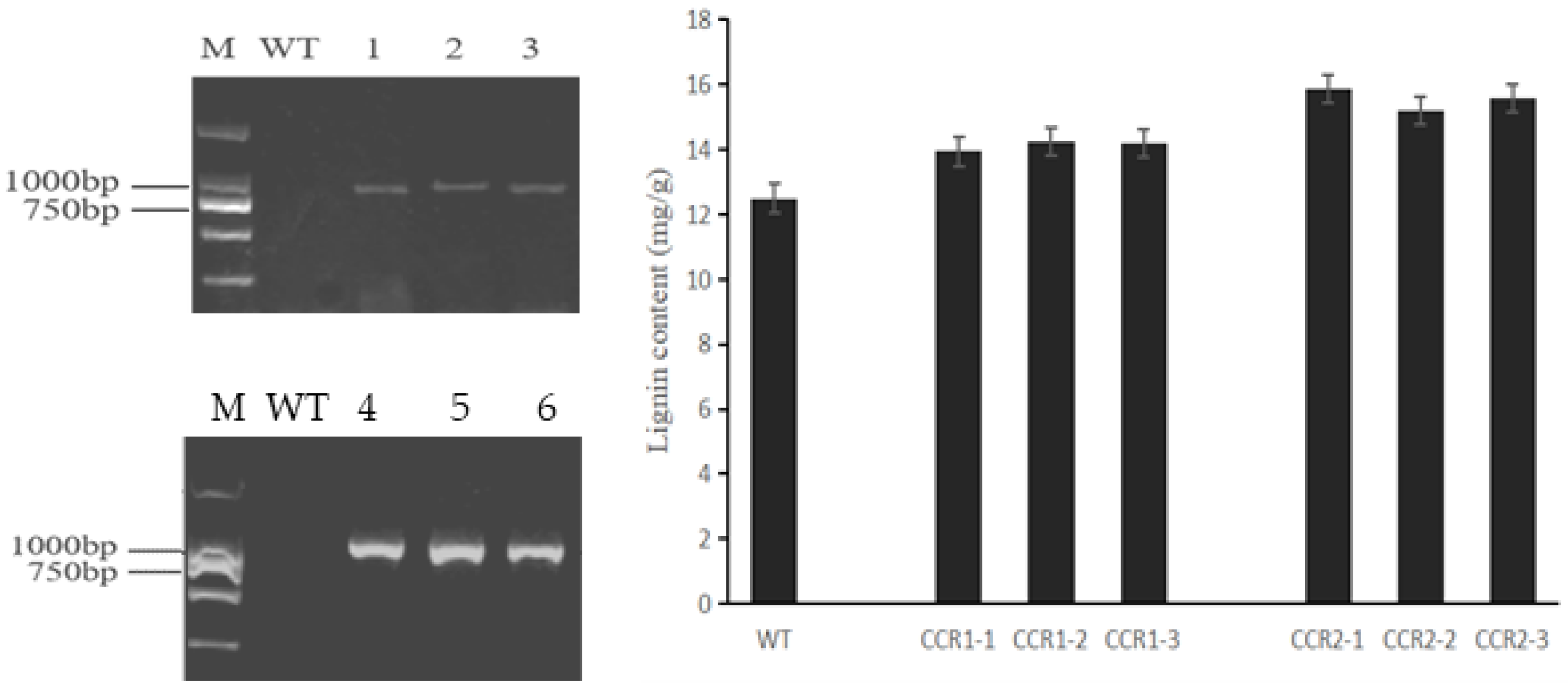
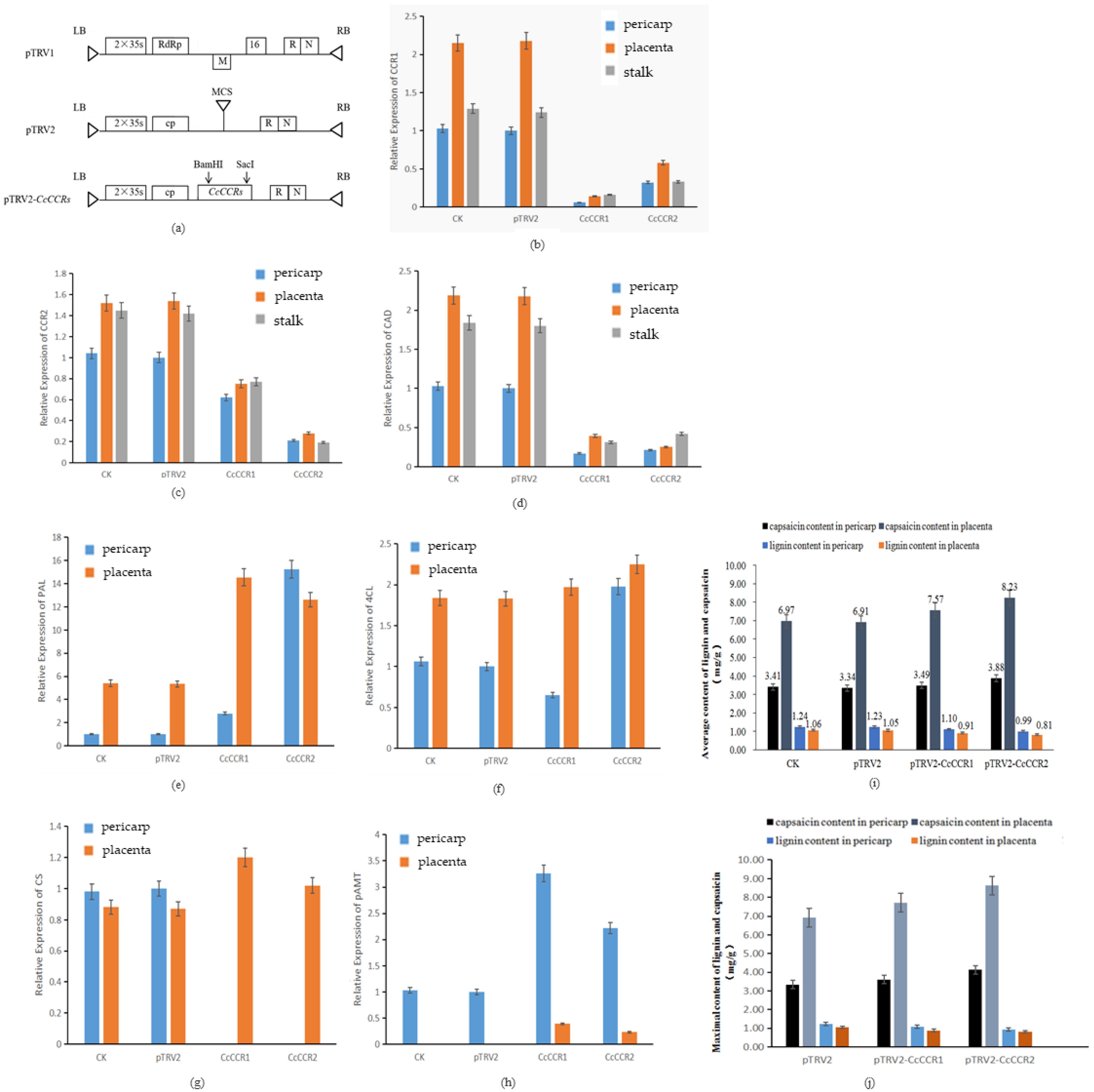
3.7. Functional Confirmation in the Regulation of Pungency in Pepper
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QC (accessed on 25 April 2022).
- Barbero, G.F.; de Aguiar, A.C.; Carrera, C.; Olachea, Á.; Ferreiro-González, M.; Martínez, J.; Palma, M.; Barroso, C.G. Evolution of Capsaicinoids in Peter Pepper (Capsicum Annuum Var. Annuum) During Fruit Ripening. Chem. Biodivers. 2016, 13, 1068–1075. [Google Scholar] [CrossRef]
- Sharma, S.K.; Vij, A.S.; Sharma, M. Mechanisms and Clinical Uses of Capsaicin. Eur. J. Pharmacol. 2013, 720, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Ma, Y.; Dai, X.; Li, X.; Yang, S. Spread and industry development of pepper in China. Acta Hortic. Sin. 2020, 47, 1715–1716. [Google Scholar]
- Aza-González, C.; Núñez-Palenius, H.G.; Ochoa-Alejo, N. Molecular Biology of Capsaicinoid Biosynthesis in Chili Pepper (Capsicum Spp.). Plant Cell Rep. 2011, 30, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Bosland, P.W.; Coon, D.; Cooke, P.H. Novel Formation of Ectopic (Nonplacental) Capsaicinoid Secreting Vesicles on Fruit Walls Explains the Morphological Mechanism for Super-Hot Chile Peppers. J. Amer. Soc. Hort. Sci. 2015, 140, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Fujiwake, H.; Suzuki, T.; Iwai, K. Capsaicinoid Formation in the Protoplast from the Placenta of Capsicum Fruits. Chem. Biol. Technol. Agric. 1982, 46, 2591–2592. [Google Scholar] [CrossRef] [Green Version]
- Mazourek, M.; Pujar, A.; Borovsky, Y.; Paran, I.; Mueller, L.; Jahn, M.M. A Dynamic Interface for Capsaicinoid Systems Biology. Plant Physiol. 2009, 150, 1806–1821. [Google Scholar] [CrossRef] [Green Version]
- Urrutia, M.; Schwab, W.; Hoffmann, T.; Monfort, A. Genetic Dissection of the (Poly)Phenol Profile of Diploid Strawberry (Fragaria Vesca) Fruits Using a NIL Collection. Plant Sci. 2016, 242, 151–168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Butelli, E.; Alseekh, S.; Tohge, T.; Rallapalli, G.; Luo, J.; Kawar, P.G.; Hill, L.; Santino, A.; Fernie, A.R.; et al. Multi-Level Engineering Facilitates the Production of Phenylpropanoid Compounds in Tomato. Nat. Commun. 2015, 6, 8635. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; He, L.; Chen, J.; Gao, Y. Effects of Light Intensity on Contents of Capsaicin and Its Metabolizing Competition Matters in Pepper Fruits. J. Henan Agric. Sci. 2008, 4, 83–86. [Google Scholar]
- Wang, S.; He, L.; Chen, J.; Geng, L.; Tong, X. Effects of nitrogen on contents of aapsaicin and its same substrates in pepper fruit. J. Shenyang Agric. Univ. 2009, 40, 144–147. [Google Scholar]
- He, L.L.; Jia, Q.; Wang, S.J.; Yang, J.; Gao, S. Effects of PUT and SPD on Capsaicin and Its Metabolism-related Substances in Pepper. Acta Agr. Bor.-Occid. Sin. 2010, 19, 153–156+175. [Google Scholar]
- Estrada, B.; Pomar, F.; Díaz, J.; Merino, F.; Bernal, M.A. Effects of Mineral Fertilizer Supplementation on Fruit Development and Pungency in ‘Padrón’ Peppers. J. Hortic. Sci. Biotech. 1998, 73, 493–497. [Google Scholar] [CrossRef]
- Nuñez-Palenius, H.G.; Ochoa-Alejo, N. Effect of Phenylalanine and Phenylpropanoids on the Accumulation of Capsaicinoids and Lignin in Cell Cultures of Chili Pepper (Capsicum Annuum L.). Vitr. Cell Dev. Plant 2005, 41, 801–805. [Google Scholar] [CrossRef]
- Lacombe, E.; Hawkins, S.; Van Doorsselaere, J.; Piquemal, J.; Goffner, D.; Poeydomenge, O.; Boudet, A.-M.; Grima-Pettenati, J. Cinnamoyl CoA Reductase, the First Committed Enzyme of the Lignin Branch Biosynthetic Pathway: Cloning, Expression and Phylogenetic Relationships. Plant J. 1997, 11, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Goujon, T.; Ferret, V.; Mila, I.; Pollet, B.; Ruel, K.; Burlat, V.; Joseleau, J.-P.; Barrière, Y.; Lapierre, C.; Jouanin, L. Down-Regulation of the AtCCR1 Gene in Arabidopsis Thaliana: Effects on Phenotype, Lignins and Cell Wall Degradability. Planta 2003, 217, 218–228. [Google Scholar] [CrossRef]
- Leplé, J.-C.; Dauwe, R.; Morreel, K.; Storme, V.; Lapierre, C.; Pollet, B.; Naumann, A.; Kang, K.-Y.; Kim, H.; Ruel, K.; et al. Downregulation of Cinnamoyl-Coenzyme A Reductase in Poplar: Multiple-Level Phenotyping Reveals Effects on Cell Wall Polymer Metabolism and Structure. Plant Cell 2007, 19, 3669–3691. [Google Scholar] [CrossRef] [Green Version]
- Escamilla-Treviño, L.L.; Shen, H.; Uppalapati, S.R.; Ray, T.; Tang, Y.; Hernandez, T.; Yin, Y.; Xu, Y.; Dixon, R.A. Switchgrass (Panicum Virgatum) Possesses a Divergent Family of Cinnamoyl CoA Reductases with Distinct Biochemical Properties. New Phytol. 2010, 185, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Giordano, A.; Liu, Z.; Panter, S.N.; Dimech, A.M.; Shang, Y.; Wijesinghe, H.; Fulgueras, K.; Ran, Y.; Mouradov, A.; Rochfort, S.; et al. Reduced Lignin Content and Altered Lignin Composition in the Warm Season Forage Grass Paspalum Dilatatum by Down-Regulation of a Cinnamoyl CoA Reductase Gene. Transgenic. Res. 2014, 23, 503–517. [Google Scholar] [CrossRef] [Green Version]
- Lauvergeat, V.; Lacomme, C.; Lacombe, E.; Lasserre, E.; Roby, D.; Grima-Pettenati, J. Two Cinnamoyl-CoA Reductase (CCR) Genes from Arabidopsis Thaliana Are Differentially Expressed during Development and in Response to Infection with Pathogenic Bacteria. Phytochemistry 2001, 57, 1187–1195. [Google Scholar] [CrossRef]
- Ma, Q.-H.; Wang, C.; Zhu, H.-H. TaMYB4 Cloned from Wheat Regulates Lignin Biosynthesis through Negatively Controlling the Transcripts of Both Cinnamyl Alcohol Dehydrogenase and Cinnamoyl-CoA Reductase Genes. Biochimie 2011, 93, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Zhou, R.; Louie, G.V.; Mühlemann, J.K.; Bomati, E.K.; Bowman, M.E.; Dudareva, N.; Dixon, R.A.; Noel, J.P.; Wang, X. Structural Studies of Cinnamoyl-CoA Reductase and Cinnamyl-Alcohol Dehydrogenase, Key Enzymes of Monolignol Biosynthesis. Plant Cell 2014, 26, 3709–3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamasloukht, B.; Wong Quai Lam, M.S.-J.; Martinez, Y.; Tozo, K.; Barbier, O.; Jourda, C.; Jauneau, A.; Borderies, G.; Balzergue, S.; Renou, J.-P.; et al. Characterization of a Cinnamoyl-CoA Reductase 1 (CCR1) Mutant in Maize: Effects on Lignification, Fibre Development, and Global Gene Expression. J. Exp. Bot. 2011, 62, 3837–3848. [Google Scholar] [CrossRef] [PubMed]
- Van Acker, R.; Leplé, J.-C.; Aerts, D.; Storme, V.; Goeminne, G.; Ivens, B.; Légée, F.; Lapierre, C.; Piens, K.; Van Montagu, M.C.E.; et al. Improved Saccharification and Ethanol Yield from Field-Grown Transgenic Poplar Deficient in Cinnamoyl-CoA Reductase. Proc. Natl. Acad. Sci. USA 2014, 111, 845–850. [Google Scholar] [CrossRef] [Green Version]
- Barakat, A.; Yassin, N.B.M.; Park, J.S.; Choi, A.; Herr, J.; Carlson, J.E. Comparative and Phylogenomic Analyses of Cinnamoyl-CoA Reductase and Cinnamoyl-CoA-Reductase-like Gene Family in Land Plants. Plant Sci. 2011, 181, 249–257. [Google Scholar] [CrossRef]
- Zhou, R.; Jackson, L.; Shadle, G.; Nakashima, J.; Temple, S.; Chen, F.; Dixon, R.A. Distinct Cinnamoyl CoA Reductases Involved in Parallel Routes to Lignin in Medicago Truncatula. Proc. Natl. Acad. Sci. USA 2010, 107, 17803–17808. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-X.; Zhao, S.-N.; Liu, G.-F.; Huang, Z.-M.; Cao, Z.-M.; Cheng, S.-H.; Lin, S.-S. Discovery of Putative Capsaicin Biosynthetic Genes by RNA-Seq and Digital Gene Expression Analysis of Pepper. Sci. Rep. 2016, 6, 34121. [Google Scholar] [CrossRef] [Green Version]
- Sarni, F.; Grand, C.; Boudet, A.M. Purification and Properties of Cinnamoyl-CoA Reductase and Cinnamyl Alcohol Dehydrogenase from Poplar Stems (Populus X Euramericana). Eur. J. Biochem. 1984, 139, 259–265. [Google Scholar] [CrossRef]
- Goffner, D.; Campbell, M.M.; Campargue, C.; Clastre, M.; Borderies, G.; Boudet, A.; Boudet, A.M. Purification and Characterization of Cinnamoyl-Coenzyme A:NADP Oxidoreductase in Eucalyptus Gunnii. Plant Physiol. 1994, 106, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Lüderitz, T.; Grisebach, H. Enzymic Synthesis of Lignin Precursors Comparison of Cinnamoyl-CoA Reductase and Cinnamyl Alcohol: NADP+ Dehydrogenase from Spruce (Picea Abies L.) and Soybean (Glycine Max L.). Eur. J. Bio. 1981, 119, 115–124. [Google Scholar] [CrossRef]
- Wengenmayer, H.; Ebel, J.; Grisebach, H. Enzymic Synthesis of Lignin Precursors. Purification and Properties of a Cinnamoyl-CoA: NADPH Reductase from Cell Suspension Cultures of Soybean (Glycine Max). Eur. J. Biochem. 1976, 65, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Pichon, M.; Courbou, I.; Beckert, M.; Boudet, A.M.; Grima-Pettenati, J. Cloning and characterization of two maize cDNAs encoding Cinnamoyl-CoA Reductase (CCR) and differential expression of the corresponding genes. Plant Mol. Biol. 1998, 38, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.-H. Characterization of a Cinnamoyl-CoA Reductase That Is Associated with Stem Development in Wheat. J. Exp. Bot. 2007, 58, 2011–2021. [Google Scholar] [CrossRef] [PubMed]
- Park, H.L.; Bhoo, S.H.; Kwon, M.; Lee, S.-W.; Cho, M.-H. Biochemical and Expression Analyses of the Rice Cinnamoyl-CoA Reductase Gene Family. Front. Plant Sci. 2017, 8, 2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, M.E.; Blasco, R. Expansion of the Mammalian 3β-Hydroxysteroid Dehydrogenase/Plant Dihydroflavonol Reductase Superfamily to Include a Bacterial Cholesterol Dehydrogenase, a Bacterial UDP-Galactose-4-Epimerase, and Open Reading Frames in Vaccinia Virus and Fish Lymphocystis. FEBS Lett. 1992, 301, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Chao, N.; Li, N.; Qi, Q.; Li, S.; Lv, T.; Jiang, X.-N.; Gai, Y. Characterization of the Cinnamoyl-CoA Reductase (CCR) Gene Family in Populus Tomentosa Reveals the Enzymatic Active Sites and Evolution of CCR. Planta 2017, 245, 61–75. [Google Scholar] [CrossRef]
- Shi, R.; Sun, Y.-H.; Li, Q.; Heber, S.; Sederoff, R.; Chiang, V.L. Towards a Systems Approach for Lignin Biosynthesis in Populus Trichocarpa: Transcript Abundance and Specificity of the Monolignol Biosynthetic Genes. Plant Cell Physiol. 2010, 51, 144–163. [Google Scholar] [CrossRef] [Green Version]
- Carocha, V.; Soler, M.; Hefer, C.; Cassan-Wang, H.; Fevereiro, P.; Myburg, A.A.; Paiva, J.A.P.; Grima-Pettenati, J. Genome-wide Analysis of the Lignin Toolbox of E Ucalyptus Grandis. New Phytol. 2015, 206, 1297–1313. [Google Scholar] [CrossRef] [Green Version]
- Van Parijs, F.R.D.; Ruttink, T.; Boerjan, W.; Haesaert, G.; Byrne, S.L.; Asp, T.; Roldán-Ruiz, I.; Muylle, H. Clade Classification of Monolignol Biosynthesis Gene Family Members Reveals Target Genes to Decrease Lignin in Lolium Perenne. Plant Biol. J. 2015, 17, 877–892. [Google Scholar] [CrossRef]
- Cheng, X.; Li, M.; Li, D.; Zhang, J.; Jin, Q.; Sheng, L.; Cai, Y.; Lin, Y. Characterization and Analysis of CCR and CAD Gene Families at the Whole-Genome Level for Lignin Synthesis of Stone Cells in Pear (Pyrus Bretschneideri). Fruit. Biol. Open 2017, 6, 1602–1613. [Google Scholar] [CrossRef] [Green Version]
- Yin, N.; Li, B.; Liu, X.; Liang, Y.; Lian, J.; Xue, Y.; Qu, C.; Lu, K.; Wei, L.; Wang, R.; et al. Two Types of Cinnamoyl-CoA Reductase Function Divergently in Accumulation of Lignins, Flavonoids and Glucosinolates and Enhance Lodging Resistance in Brassica Napus. Crop J. 2021, in press. [CrossRef]
- Mir Derikvand, M.; Sierra, J.B.; Ruel, K.; Pollet, B.; Do, C.-T.; Thévenin, J.; Buffard, D.; Jouanin, L.; Lapierre, C. Redirection of the Phenylpropanoid Pathway to Feruloyl Malate in Arabidopsis Mutants Deficient for Cinnamoyl-CoA Reductase 1. Planta 2008, 227, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Ruel, K.; Berrio-Sierra, J.; Derikvand, M.M.; Pollet, B.; Thévenin, J.; Lapierre, C.; Jouanin, L.; Joseleau, J. Impact of CCR1 Silencing on the Assembly of Lignified Secondary Walls in Arabidopsis Thaliana. New Phytol. 2009, 184, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fan, F.; Wang, L.; Zhan, Q.; Wu, P.; Du, J.; Yang, X.; Liu, Y. Cloning and Expression Analysis of Cinnamoyl-CoA Reductase (CCR) Genes in Sorghum. PeerJ. 2016, 4, e2005. [Google Scholar] [CrossRef]
- Li, L.; Cheng, X.; Lu, S.; Nakatsubo, T.; Umezawa, T.; Chiang, V.L. Clarification of Cinnamoyl Co-Enzyme A Reductase Catalysis in Monolignol Biosynthesis of Aspen. Plant Cell Physiol. 2005, 46, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Larsen, K. Molecular Cloning and Characterization of CDNAs Encoding Cinnamoyl CoA Reductase (CCR) from Barley (Hordeum Vulgare) and Potato (Solanum Tuberosum). J. Plant Physiol. 2004, 161, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, P.; Vishwakarma, R.K.; Khan, B.M. Biochemical Characterization of Recombinant Cinnamoyl CoA Reductase 1 (Ll-CCRH1) from Leucaena Leucocephala. Int. J. Biol. Macromol. 2013, 58, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Dauwe, R.; Morreel, K.; Goeminne, G.; Gielen, B.; Rohde, A.; Van Beeumen, J.; Ralph, J.; Boudet, A.-M.; Kopka, J.; Rochange, S.F.; et al. Molecular Phenotyping of Lignin-Modified Tobacco Reveals Associated Changes in Cell-Wall Metabolism, Primary Metabolism, Stress Metabolism and Photorespiration: Molecular Phenotyping of Lignin-Modified Tobacco. Plant J. 2007, 52, 263–285. [Google Scholar] [CrossRef]
- Van der Rest, B.; Danoun, S.; Boudet, A.-M.; Rochange, S.F. Down-Regulation of Cinnamoyl-CoA Reductase in Tomato (Solanum Lycopersicum L.) Induces Dramatic Changes in Soluble Phenolic Pools. J. Exp. Bot. 2006, 57, 1399–1411. [Google Scholar] [CrossRef] [Green Version]
- Arce-Rodríguez, M.L.; Ochoa-Alejo, N. Silencing AT3 Gene Reduces the Expression of PAmt, BCAT, Kas, and Acl Genes Involved in Capsaicinoid Biosynthesis in Chili Pepper Fruits. Biol. Plant. 2015, 59, 477–484. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.-T.; et al. Genome Sequence of the Hot Pepper Provides Insights into the Evolution of Pungency in Capsicum Species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Name | Gene Description | ORF b | Protein Size c | Theoretical MW c (kDa) | pI | Chromosome |
|---|---|---|---|---|---|---|---|
| XP_016548138.1 | CaCCR1-1 | PREDICTED: cinnamoyl-CoA reductase 1 | 975 | 324 | 35.8 | 5.36 | 11 |
| XP_016561561.1 | CaCCR1-2 | PREDICTED: cinnamoyl-CoA reductase 1 | 1089 | 362 | 40.1 | 6.90 | 2 |
| XP_016562886.1 | CaCCR1-like1 | PREDICTED: cinnamoyl-CoA reductase 1-like | 1005 | 334 | 36.9 | 6.41 | 3 |
| XP_016562887.1 | CaCCR1-like2 | PREDICTED: cinnamoyl-CoA reductase 1-like | 1005 | 334 | 37.1 | 6.46 | 3 |
| XP_016576987.1 | CaCCR1-like3 | PREDICTED: cinnamoyl-CoA reductase 1-like | 963 | 320 | 35.3 | 6.61 | 1 |
| XP_016577001.1 | CaCCR1-like4 | PREDICTED: cinnamoyl-CoA reductase 1-like | 990 | 329 | 36.0 | 6.33 | 1 |
| XP_016558919.1 | CaCCR1-like5 | PREDICTED: cinnamoyl-CoA reductase 1-like | 453 | 150 | 16.7 | 8.54 | 1 |
| XP_016556899.1 | CaCCR1-like6 | PREDICTED: cinnamoyl-CoA reductase 1-like | 987 | 328 | 35.9 | 7.60 | 1 |
| XP_016538689.1 | CaCCR1-like7 | PREDICTED: cinnamoyl-CoA reductase 1-like | 987 | 328 | 35.8 | 5.86 | 1 |
| XP_016562803.1 | CaCCR1-like8 | PREDICTED: cinnamoyl-CoA reductase 1-like | 276 | 91 | 10.1 | 9.78 | 1 |
| XP_016564118.1 | CaCCR1-like9 | PREDICTED: cinnamoyl-CoA reductase 1-like isoform X1 | 990 | 329 | 36.8 | 5.19 | 3 |
| XP_016575015.1 | CaCCR1-like10A | PREDICTED: cinnamoyl-CoA reductase 1-like isoform X1 | 945 | 314 | 35.3 | 4.97 | 6 |
| XP_016575016.1 | CaCCR1-like10B | PREDICTED: cinnamoyl-CoA reductase 1-like isoform X2 | 942 | 313 | 35.3 | 4.97 | 6 |
| XP_016538911.1 | CaCCR2-1 | PREDICTED: cinnamoyl-CoA reductase 2 | 981 | 326 | 35.2 | 6.25 | 8 |
| XP_016578459.1 | CaCCR2-2 | PREDICTED: cinnamoyl-CoA reductase 2 | 999 | 332 | 36.7 | 6.02 | 6 |
| XP_016568210.1 | CaCCR2-like1 | PREDICTED: cinnamoyl-CoA reductase 2-like | 966 | 321 | 35.5 | 5.86 | 4 |
| XP_016572918.1 | CaCCR2-like2 | PREDICTED: cinnamoyl-CoA reductase 2-like | 963 | 320 | 35.8 | 5.39 | 1 |
| XP_016581443.1 | CaCCR2-like3 | PREDICTED: cinnamoyl-CoA reductase 2-like | 1002 | 333 | 37.2 | 8.80 | 1 |
| XP_016564119.1 | CaCCR2-like4 | PREDICTED: cinnamoyl-CoA reductase 2-like isoform X2 | 831 | 276 | 31.0 | 5.18 | 3 |
| NP_001311706.1 | CaDFR-1 | PREDICTED: dihydroflavonol-4-reductase | 1149 | 382 | 42.5 | 562 | 2 |
| XP_016556013.1 | CaDFR-like | PREDICTED: dihydroflavonol-4-reductase-like | 1029 | 342 | 38.0 | 5.95 | Un |
| PHT59959.1 | CbCCR1-1 | Cinnamoyl-CoA reductase 1 | 1002 | 333 | 37.2 | 8.46 | 1 |
| PHT59278.1 | CbCCR1-2 | Cinnamoyl-CoA reductase 1 | 990 | 329 | 36.0 | 6.61 | 1 |
| PHT59279.1 | CbCCR1-3 | Cinnamoyl-CoA reductase 1 | 963 | 320 | 35.3 | 6.61 | 1 |
| PHT46015.1 | CbCCR2-1 | Cinnamoyl-CoA reductase 2 | 999 | 332 | 36.8 | 6.02 | 6 |
| PHT47585.1 | CbCCR2-2 | Cinnamoyl-CoA reductase 2 | 1005 | 334 | 36.9 | 7.03 | 5 |
| PHT47586.1 | CbCCR2-3 | Cinnamoyl-CoA reductase 2 | 1185 | 394 | 43.8 | 8.56 | 5 |
| PHT54849.1 | CbDFR-1 | Cinnamoyl CoA reductase family/Dihydroflavonol-4-reductase | 1356 | 451 | 50.3 | 8.49 | 2 |
| PHT56606.1 | CbDFR-2 | Cinnamoyl CoA reductase family/Dihydroflavonol-4-reductase | 1149 | 382 | 42.5 | 5.62 | 2 |
| PHU29451.1 | CcCCR1-1 | Cinnamoyl-CoA reductase 1 | 963 | 320 | 35.3 | 6.61 | 1 |
| PHU29452.1 | CcCCR1-2 | Cinnamoyl-CoA reductase 1 | 990 | 329 | 36.0 | 6.33 | 1 |
| PHU30004.1 | CcCCR1-3 | Cinnamoyl-CoA reductase 1 | 1002 | 333 | 37.2 | 8.68 | 1 |
| PHU15306.1 | CcCCR1-4 | Cinnamoyl-CoA reductase 1 | 1929 | 642 | 72.3 | 5.98 | 6 |
| PHU02385.1 | CcCCR2-1 | Cinnamoyl-CoA reductase 2 | 1005 | 334 | 36.9 | 7.03 | 11 |
| PHU02386.1 | CcCCR2-2 | Cinnamoyl-CoA reductase 2 | 1005 | 334 | 37.0 | 6.71 | 11 |
| PHU29448.1 | CcDFR-1 | Cinnamoyl CoA reductase family/Dihydroflavonol-4-reductase | 255 | 84 | 9.3 | 5.16 | 1 |
| PHU27058.1 | CcDFR-2 | Cinnamoyl CoA reductase family/Dihydroflavonol-4-reductase | 1149 | 382 | 42.5 | 5.45 | 2 |
| PHU07366.1 | CcDFR-3 | Cinnamoyl CoA reductase family/Dihydroflavonol-4-reductase | 192 | 63 | 6.9 | 9.03 | 9 |
| Eyzyme | Substrate | Km (μM) | Vmax (mmolL−1min−1) | Kcat (min−1) | Kcat/Km (μM−1min−1) |
|---|---|---|---|---|---|
| CcCCR1 | Feruloyl-CoA | 23.99 ± 3.32 | 0.66 ± 0.10 | 1.14 ± 016 | 0.048 |
| Sinapoyl-COA | 32.35 ± 5.96 | 0.45 ± 0.09 | 0.77 ± 0.16 | 0.024 | |
| p-Coumaroyl-CoA | 34.52 ± 3.69 | 0.78 ± 0.08 | 1.34 ± 0.14 | 0.022 | |
| CcCCR2 | Feruloyl-CoA | 16.53 ± 1.09 | 9.20 ± 1.14 | 13.53 + 1.67 | 0.819 |
| Sinapoyl-COA | 95.87 ± 9.58 | 1.06 ± 0.11 | 1.56 ± 0.16 | 0.016 | |
| p-Coumaroyl-CoA | 107.21 ± 3.13 | 6.92 ± 1.21 | 10.17 ± 1.78 | 0.095 |
| Factor | Capsaicin | Lignin | Flavonoid | CcCCR1 | CcCCR2 |
|---|---|---|---|---|---|
| Capsaicin | 1 | ||||
| Lignin | −0.387 * | 1 | |||
| Flavonoid | 0.746 ** | −0.534 ** | 1 | ||
| CcCCR1 | −0.414 * | 0.267 | −0.252 | 1 | |
| CcCCR2 | −0.664 ** | 0.326 * | −0.719 ** | 0.590 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, D.; Ni, M.; Lei, X.; Zhang, L.; Zhang, W.; Shu, H.; Wang, Z.; Zhu, J.; Cheng, S.; Liu, P.; et al. Analyses of Pepper Cinnamoyl-CoA Reductase Gene Family and Cloning of CcCCR1/2 and Their Function Identification in the Formation of Pungency. Horticulturae 2022, 8, 537. https://doi.org/10.3390/horticulturae8060537
Wu D, Ni M, Lei X, Zhang L, Zhang W, Shu H, Wang Z, Zhu J, Cheng S, Liu P, et al. Analyses of Pepper Cinnamoyl-CoA Reductase Gene Family and Cloning of CcCCR1/2 and Their Function Identification in the Formation of Pungency. Horticulturae. 2022; 8(6):537. https://doi.org/10.3390/horticulturae8060537
Chicago/Turabian StyleWu, Dan, Miao Ni, Xin Lei, Liping Zhang, Wei Zhang, Huangying Shu, Zhiwei Wang, Jie Zhu, Shanhan Cheng, Pingwu Liu, and et al. 2022. "Analyses of Pepper Cinnamoyl-CoA Reductase Gene Family and Cloning of CcCCR1/2 and Their Function Identification in the Formation of Pungency" Horticulturae 8, no. 6: 537. https://doi.org/10.3390/horticulturae8060537
APA StyleWu, D., Ni, M., Lei, X., Zhang, L., Zhang, W., Shu, H., Wang, Z., Zhu, J., Cheng, S., Liu, P., Lv, H., & Yang, L. (2022). Analyses of Pepper Cinnamoyl-CoA Reductase Gene Family and Cloning of CcCCR1/2 and Their Function Identification in the Formation of Pungency. Horticulturae, 8(6), 537. https://doi.org/10.3390/horticulturae8060537