
Characteristics of Meteorological Conditions during a Severe Outbreak of Onion Downy Mildew and Metalaxyl Sensitivity of Peronospora destructor in Saga, Japan, in 2016


Abstract
:1. Introduction
2. Materials and Methods
2.1. Disease Development
2.2. Estimating Infection Date Using Risk Model
2.3. Infection Risk Model Ability to Predict Disease Development
2.4. Effectiveness of Metalaxyl in the Field
2.5. Metalaxyl Sensitivity Assay
3. Results
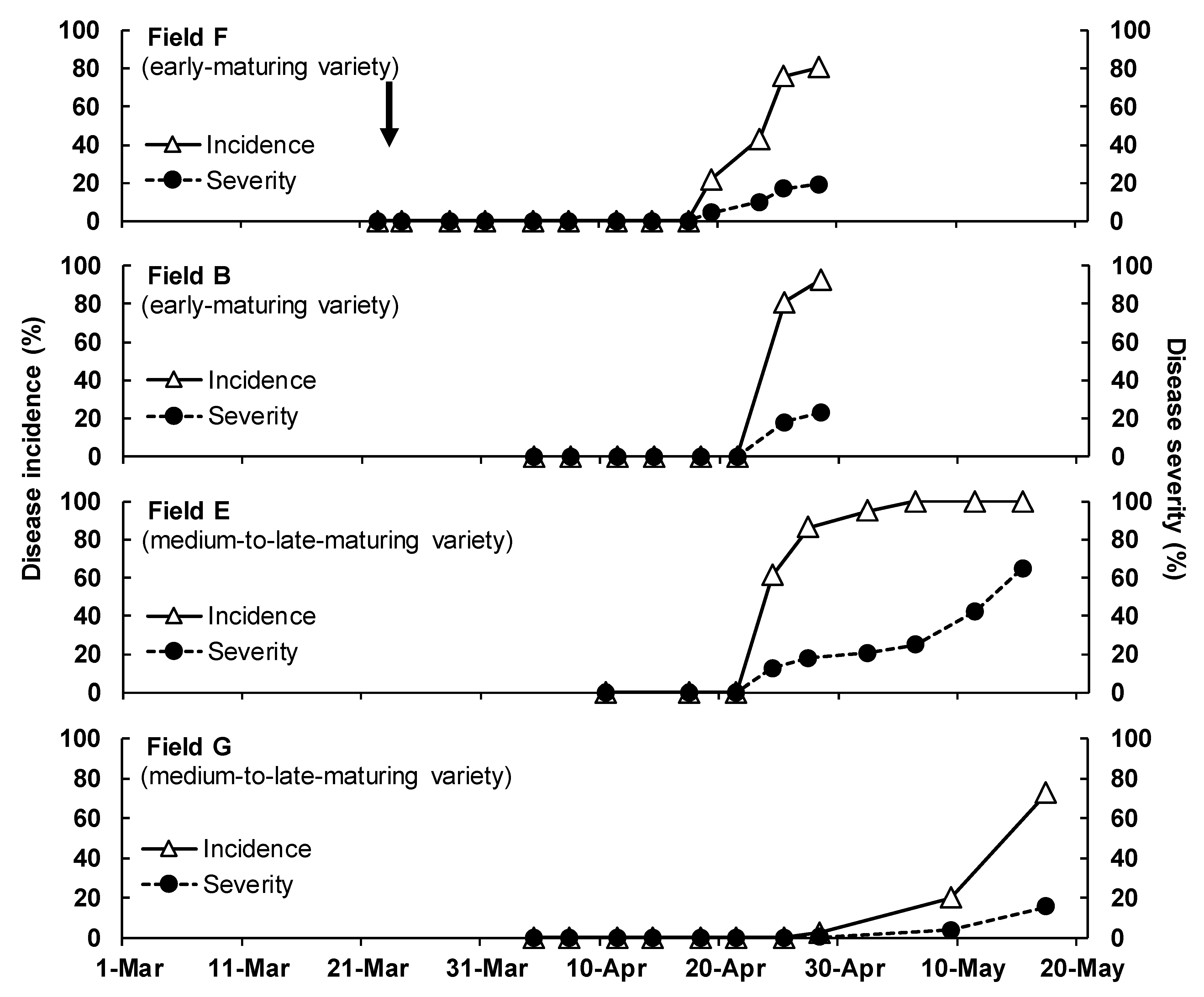
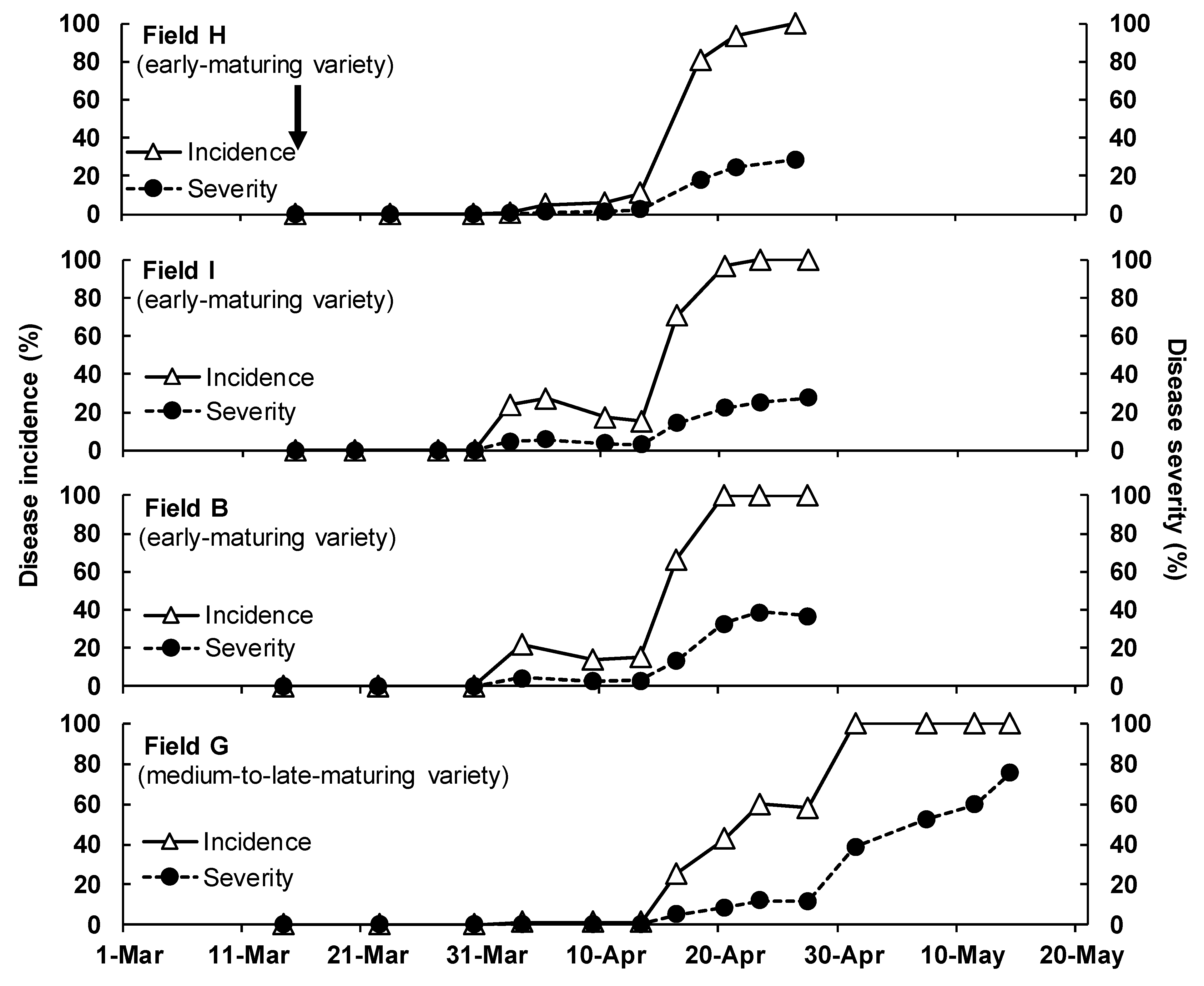
3.1. Disease Development in the Field
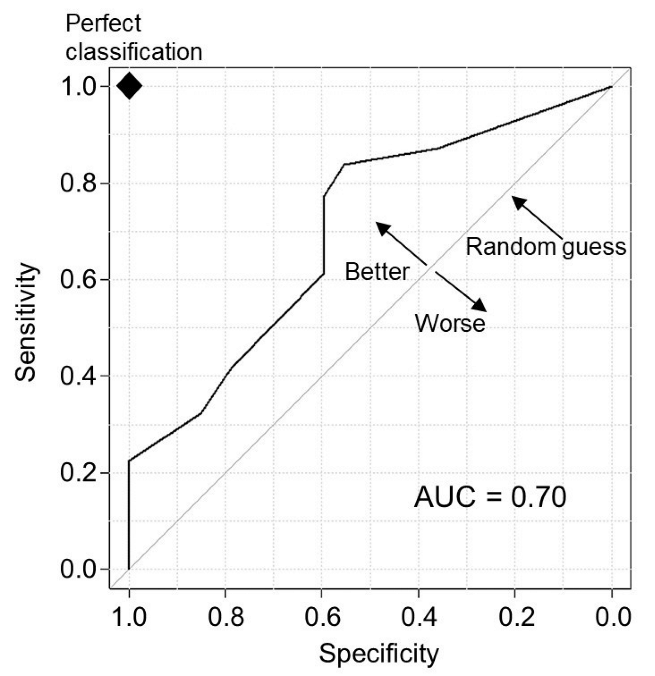
3.2. Prediction Ability of the Infection Risk Model
3.3. Effectiveness of Metalaxyl in the Field
3.4. Metalaxyl Sensitivity Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Survilienė, E.; Valiuškaitė, A.; Raudonis, L. The effect of fungicides on the development of downy mildew of onions. Zemdirbyste 2008, 95, 171–179. [Google Scholar]
- Afifi, M.; Zayan, S. Development of forecasting system for onion downy mildew in Egypt. J. Prod. Dev. 2009, 14, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Parkunan, V.; Gitaitis, R.D.; Dutta, B.; Langston, D.B.; Ji, P. An epidemic of downy mildew caused by Peronospora destructor on Vidalia sweet onions in Georgia in 2012. Plant Health Prog. 2013, 14, 54. [Google Scholar] [CrossRef] [Green Version]
- Van der Heyden, H.; Dutilleul, P.; Charron, J.-B.; Bilodeau, G.J.; Carisse, O. Factors influencing the occurrence of onion downy mildew (Peronospora destructor) epidemics: Trends from 31 years of observational data. Agronomy 2020, 10, 738. [Google Scholar] [CrossRef]
- Yarwood, C.E. Onion downy mildew. Hilgardia 1943, 14, 595–691. [Google Scholar] [CrossRef] [Green Version]
- Demizu, T. Epidemiological and therapeutic studies on the downy mildew of onions. Bull. Osaka Agric. Exp. Stn. 1963, 4, 1–95. [Google Scholar]
- Gilles, T.; Phelps, K.; Clarkson, J.P.; Kennedy, R. Development of MILIONCAST, an improved model for predicting downy mildew sporulation on onions. Plant Dis. 2004, 88, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, P.D.; Sutton, J.C. Weather variables in relation to an epidemic of onion downy mildew. Phytopathology 1982, 72, 219–224. [Google Scholar] [CrossRef]
- Jesperson, G.D.; Sutton, J.C. Evaluation of a forecaster for downy mildew of onion (Allium cepa L.). Crop Prot. 1987, 6, 95–103. [Google Scholar] [CrossRef]
- De Visser, C.L.M. Development of a downy mildew advisory model based on downcast. Eur. J. Plant Pathol. 1998, 104, 933–943. [Google Scholar] [CrossRef]
- Friedrich, S.; Leinhos, G.M.E.; Löpmeier, F.-J. Development of ZWIPERO, a model forecasting sporulation and infection periods of onion downy mildew based on meteorological data. Eur. J. Plant Pathol. 2003, 109, 35–45. [Google Scholar] [CrossRef]
- Gisi, U. Chemical control of downy mildews. In Advances in Downy Mildew Research; Spencer-Phillips, P.T.N., Gisi, U., Lebeda, A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 119–159. [Google Scholar]
- Gisi, U.; Sierotzki, H. Fungicide modes of action and resistance in downy mildews. Eur. J. Plant Pathol. 2008, 122, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Fungicide Resistance Action Committee. FRAC Code List 2022. Available online: http:/www.frac.info (accessed on 17 June 2022).
- O’Brien, R.G. Control of onion downy mildew in the presence of phenylamide-resistant strains of Peronospora destructor (Berk.) Caspary. Aust. J. Exp. Agric. 1992, 32, 669–674. [Google Scholar] [CrossRef]
- O’Brien, R.G.; Galea, V.; MacManus, G. VG402: Development of an Onion Downy Mildew Forecasting System. Horticulture Australia Ltd. 1997. Available online: https://ausveg.com.au/app/data/technical-insights/docs/VG402.pdf (accessed on 2 February 2022).
- Wright, P.J. Resistance of Peronospora destructor (onion downy mildew) to metalaxyl fungicides. In Proceedings of the 57th New Zealand Plant Protection Society’s Annual Conference, Rydges Le Grand. Hamilton, New Zealand, 10–12 August 2004; p. 347. [Google Scholar] [CrossRef]
- Sutton, J.C.; Hildebrand, P.D. Environmental water in relation to Peronospora destructor and related pathogens. Can. J. Plant Pathol. 1985, 7, 323–330. [Google Scholar] [CrossRef]
- Gilles, T. Forecasting downy mildew diseases. In Advances in Downy Mildew Research; Spencer-Phillips, P.T.N., Jeger, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; Volume 2, pp. 35–67. [Google Scholar]
- OEPP/EPPO. Guidelines on good plant protection practice: ALLIUM CROPS. Bull. OEPP/EPPO Bull. 2001, 31, 211–230. [Google Scholar] [CrossRef]
- Bashi, E.; Aylor, D.E. Survival of detached sporangia of Peronospora destructor and Peronospora tabacina. Phytopathology 1983, 73, 1135–1139. [Google Scholar] [CrossRef]
- Palti, J. Epidemiology, prediction and control of onion downy mildew caused by Peronospora destructor. Phytoparasitica 1989, 17, 31–48. [Google Scholar] [CrossRef]
- DEFRA. Forecasting Diseases of Allium Crops: Project Report HH1742SFV; Department for Environment, Food and Rural Affairs: London, UK, 2002; pp. 11–18. Available online: http://randd.defra.gov.uk/Default.aspx?Menu=Menu&Module=More&Location=None&Completed=0&ProjectID=7930 (accessed on 2 February 2022).
- Leach, C.M. Effect of still and moving moisture-saturated air on sporulation of Drechslera and Peronospora. Trans. Br. Mycol. Soc. 1985, 84, 179–183. [Google Scholar] [CrossRef]
- Su, H.; van Bruggen, A.H.C.; Subbarao, K.V.; Scherm, H. Sporulation of Bremia lactucae affected by temperature, relative humidity, and wind in controlled conditions. Phytopathology 2004, 94, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Scherm, H.; van Bruggen, A.H.C. Response surface models for germination and infection of Bremia lactucae, the fungus causing downy mildew of lettuce. Ecol. Modell. 1993, 65, 281–296. [Google Scholar] [CrossRef]
- Scherm, H.; van Bruggen, A.H.C. Weather variables associated with infection of lettuce by downy mildew (Bremia lactucae) in coastal California. Phytopathology 1994, 84, 860–865. [Google Scholar] [CrossRef]
- Ogura, Y. General Meteorology, 2nd ed.; University of Tokyo Press: Tokyo, Japan, 1999. [Google Scholar]
- Brischetto, C.; Bove, F.; Languasco, L.; Rossi, V. Can spore sampler data be used to predict Plasmopara viticola infection in vineyards? Front. Plant Sci. 2020, 11, 1187. [Google Scholar] [CrossRef]
- Brischetto, C.; Bove, F.; Fedele, G.; Rossi, V. A weather-driven model for predicting infections of grapevines by sporangia of Plasmopara viticola. Front. Plant Sci. 2021, 12, 636607. [Google Scholar] [CrossRef] [PubMed]
- Turechek, W.W.; Wilcox, W.F. Evaluating predictors of apple scab with receiver operating characteristic curve analysis. Phytopathology 2005, 95, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Mandrekar, J.N. Receiver operating characteristic curve in diagnostic test assessment. J. Thorac. Oncol. 2010, 5, 1315–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youden, W.J. Index for rating diagnostic tests. Cancer 1950, 3, 32–35. [Google Scholar] [CrossRef]
- Schwinn, F.J.; Staub, T. Oomycetes fungicides: Phenylamides and other fungicides against Oomycetes. In Modern Selective Fungicides, 2nd ed.; Lyr, H., Ed.; Gustav Fischer: Jena, Germany, 1995; pp. 323–346. [Google Scholar]
- Ankita; Chandel, S.; Sharma, R.; Meena, V.K. Epidemiological studies of downy mildew of onion. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 1–12. [Google Scholar] [CrossRef]
- Nuninger, C.; Watson, G.; Leadbitter, N.; Ellgehausen, H. CGA 329351: Introduction of the enantiomeric from of the fungicide metalaxyl. In Proceedings of the Brighton Crop Protection Conference: Pest and Disease, British Crop Protection Council. Farnham, UK, 18–21 November 1996; Volume 1, pp. 41–46. [Google Scholar]
- Bliss, C.I. The method of probits. Science 1934, 79, 38–39. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Marcuzzo, L.L.; Carvalho, J.; Nascimento, A. Epidemiological aspects of onion downy mildew in the region of Alto Vale do Itajaí, Santa Catarina State, Brazil. Summa Phytopathol. 2017, 43, 218–222. [Google Scholar] [CrossRef] [Green Version]
- González, P.H.; Colnago, P.; Peluffo, S.; Idiarte, H.G.; Zipitría, J.; Galván, G.A. Quantitative studies on downy mildew (Peronospora destructor Berk. Casp.) affecting onion seed production in southern Uruguay. Eur. J. Plant Pathol. 2011, 129, 303–314. [Google Scholar] [CrossRef]
- Maeso, D. Enfermedades del cultivo de cebolla. In Tecnología Para la Producción de Cebolla; Arboleya, J., Ed.; INIA Boletín de divulgación 88: Montevideo, Uruguay, 2005; pp. 151–188. [Google Scholar]
- Chynoweth, R.W.; Beresford, R.M.; Henshall, W.R.; Wright, P.J. Use of disease forecasting models for control of onion downy mildew in New Zealand. In Advances in Downy Mildew Research; Spencer-Phillips, P.T.N., Jeger, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; Volume 2, pp. 91–97. [Google Scholar]
- Koshimizu, Y. A forecasting method for occurrence of rice leaf blast with AMeDAS data. Bull. Tohoku Natl. Agric. Exp. Stn. 1988, 78, 67–121. [Google Scholar]
- Neufeld, K.N.; Keinath, A.P.; Ojiambo, P.S. A model to predict the risk of infection of cucumber by Pseudoperonospora cubensis. Microb. Risk Anal. 2017, 6, 21–30. [Google Scholar] [CrossRef]
- Neufeld, K.N.; Keinath, A.P.; Ojiambo, P.S. Evaluation of a model for predicting the infection risk of squash and cantaloupe by Pseudoperonospora cubensis. Plant Dis. 2018, 102, 855–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celetti, M. Increased risk of downy mildew on onions. In Hort Matters; Ontario Ministry of Agriculture and Food and Ministry of Rural Affairs: Guelph, ON, Canada, 2015. Available online: http://www.omafra.gov.on.ca/english/crops/hort/news/hortmatt/2015/16hrt15a2.htm (accessed on 26 February 2022).
- Pscheidt, J.W.; Ocamb, C.M. Onion (Allium cepa)-Downy Mildew. In Pacific Northwest Plant Disease Management Handbook; Oregon State University: Corvallis, OR, USA, 2022; Available online: https://pnwhandbooks.org/plantdisease/host-disease/onion-allium-cepa-downy-mildew (accessed on 26 February 2022).
- Smith, R.W.; Lorbeer, J.W.; Abd-Elrazik, A.A. Reappearance and control of onion downy mildew epidemics in New York. Plant Dis. 1985, 69, 703–706. [Google Scholar] [CrossRef]
- Mo, C.Y.; Lee, J.H.; Ko, S.J.; Yang, K.Y. Control efficacy of several fungicides against downy mildew of onion at nursery seedling stage. Res. Plant Dis. 2016, 22, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Araújo, E.R.; Resende, R.S.; Alves, D.P.; Higashikawa, F.S. Field efficacy of fungicides to control downy mildew of onion. Eur. J. Plant Pathol. 2020, 156, 305–359. [Google Scholar] [CrossRef]
- Raziq, F.; Alam, I.; Naz, I.; Khan, H. Evaluation of fungicides for controlling downy mildew of onion under field conditions. Sarhad J. Agric. 2008, 24, 85–91. [Google Scholar]
- Araújo, E.R.; Resende, R.S. Propineb efficiency in controlling downy mildew of onion under field conditions. Australas. Plant Dis. Notes 2020, 15, 34. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Kan, M. Effects of control on rot of onion bulbs during storage through fungicide application during the growing season. Kyushu Plant Prot. Res. 1982, 28, 70–73. [Google Scholar] [CrossRef]
- Parra, G.; Ristaino, J.B. Resistance to mefenoxam and metalaxyl among field isolates of Phytophthora capsici causing Phytophthora blight of bell pepper. Plant Dis. 2001, 85, 1069–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year b | Disease Survey Field (Location) | Onion Cultivation | Assessment Period | Placement of Infected Plants (No. c) | ||||
|---|---|---|---|---|---|---|---|---|
| Variety (Maturity) | Planting Date | Harvest Date | Disease Survey Field (Unsprayed Plots) | Fungicide Experiment Plots | ||||
| 2016 | A | Saga | Takanishiki (very early) | 12 November 2015 | 15 April 2016 | 1 March–7 April | 1 March–7 April (expts. 1-1, 1-2) | 2–29 March (4 d) |
| B | Shiroishi | Shippo-wase 7 (early) | 12 November 2015 | 25 April 2016 | 3 March–1 April | 3 March–1 April (expt. 2) | – | |
| C | Saga | Spart (early) | 24 November 2015 | 2 May 2016 | 29 March–29 April | 29 March–29 April (expt. 3) | 30 March–23 April (2 d) | |
| D | Shiroishi | Advance (early) | 20 December 2015 | 18 May 2016 | 30 March–25 April | – | – | |
| E | Saga | Tarzan (medium–late) | 8 December 2015 | 27 May 2016 | 11 April–15 May | – | – | |
| 2017 | F | Saga | Spart (early) | 2 December 2016 | 1 May 2017 | 22 March–28 April | 19–25 April (expt. 4) | 23 March–19 April (4) |
| B | Shiroishi | Shippo-wase 7 (early) | 12 December 2016 | 2 May 2017 | 4 April–28 April | – | – | |
| E | Saga | Tarzan (medium–late) | 12 December 2016 | 31 May 2017 | 10 April–15 May | – | – | |
| G | Shiroishi | Tarzan (medium–late) | 21 November 2016 | 25 May 2017 | 4 April–17 May | – | – | |
| 2018 | H | Saga | Spart (early) | 21 November 2017 | 1 May 2018 | 15 March–26 April | 18–26 April (expt. 5) | 15 March–22 April (3) |
| I | Saga | Shippo-wase 7 (early) | 21 November 2017 | 11 May 2018 | 15 March–27 April | – | – | |
| B | Shiroishi | Shippo-wase 7 (early) | 27 November 2017 | 2 May 2018 | 14 March–27 April | – | – | |
| G | Shiroishi | Tarzan (medium–late) | 6 December 2017 | 15 May 2018 | 14 March–14 May | – | – | |
| Fungicide Active Ingredients (% ai) | Dosage (%) | Date of the Sprays | |||||
|---|---|---|---|---|---|---|---|
| 2016 | 2016 | 2016 | 2016 | 2017 | 2018 | ||
| Field A (Fungicide Expt. 1-1) | Field A (Fungicide Expt. 1-2) | Field B (Fungicide Expt. 2) | Field C (Fungicide Expt. 3) | Field F (Fungicide Expt. 4) | Field H (Fungicide Expt. 5) | ||
| Chlorothalonil 50% +Benthiavalicarb- isopropyl 5% | 0.1 | 1, 8, and 15 March | 15, 22, and 29 March | 3, 14, and 25 March | 29 March; 5 and 12 April | not tested | not tested |
| Chlorothalonil 32% +Metalaxyl-M 3.3% | 0.125 | 1, 8, and 15 March | 15, 22, and 29 March | 3, 14, and 25 March | 29 March; 5 and 12 April | 23 and 30 March; 5 and 12 April | 15, 23, and 30 March; 5 and 12 April |
| Chlorothalonil 40% | 0.1 | 1, 8, and 15 March | 15, 22, and 29 March | 3, 14, and 25 March | 29 March; 5 and 12 April | not tested | not tested |
| Mancozeb 80% | 0.25 | not tested | not tested | not tested | not tested | 23 and 30 March; 5 and 12 April | 15, 23, and 30 March; 5 and 12 April |
| Date | 2016 | 2017 | 2018 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Saga | Sasebo | Fukuoka | Kumamoto | Saga | Sasebo | Fukuoka | Kumamoto | Saga | Sasebo | Fukuoka | Kumamoto | |
| 1 March | + | |||||||||||
| 2 March | + | |||||||||||
| 3 March | + | + | ||||||||||
| 4 March | + + | + | + + | + | + | + + | ||||||
| 5 March | + | + | + | + | + | |||||||
| 6 March | ||||||||||||
| 7 March | + + | + + | + + | + + | ||||||||
| 8 March | + | + | + | + | + | + | ||||||
| 9 March | + | + | ||||||||||
| 10 March | + | |||||||||||
| 11 March | + | + | ||||||||||
| 12 March | + | + | ||||||||||
| 13 March | + | + | + | + | + | + | + | |||||
| 14 March | + | + | ||||||||||
| 15 March | + | + | + | |||||||||
| 16 March | ||||||||||||
| 17 March | ||||||||||||
| 18 March | ||||||||||||
| 19 March | + + | + + | + | + | + | |||||||
| 20 March | + | + | + | + | ||||||||
| 21 March | + + | + | + | + | + | |||||||
| 22 March | + | + | + + | |||||||||
| 23 March | + | |||||||||||
| 24 March | + | |||||||||||
| 25 March | + | + | ||||||||||
| 26 March | + + | + + | ||||||||||
| 27 March | + | + | + | |||||||||
| 28 March | + | |||||||||||
| 29 March | + | |||||||||||
| 30 March | + | |||||||||||
| 31 March | + + | + + | + | + + | + | + | + | |||||
| 1 April | + | + + | + + | + + | + | |||||||
| 2 April | + | + + | + + | + + | + | + | ||||||
| 3 April | + | + | + | + | + | + | ||||||
| 4 April | + | + + | + + | + + | + | + + | + | |||||
| 5 April | + | + | + | + | + + | |||||||
| 6 April | + | + | + | |||||||||
| 7 April | + | + | + | + | + + | + + | + + | + + | ||||
| 8 April | + + | + + | + + | + + | + + | + | + + | + + | ||||
| 9 April | + | + | + + | + | + + | |||||||
| 10 April | + | + | + | + | + | + | ||||||
| 11 April | + | + + | + + | + | ||||||||
| 12 April | ||||||||||||
| 13 April | ||||||||||||
| 14 April | + + | + + | ||||||||||
| 15 April | + | + | ||||||||||
| 16 April | + | + | + + | + + | ||||||||
| 17 April | + | + | + | + | + | + | + | + | ||||
| 18 April | + | + | + | + | + + | |||||||
| 19 April | ||||||||||||
| 20 April | ||||||||||||
| 21 April | + | + | + | |||||||||
| 22 April | + | + + | + | + + | + | + | ||||||
| 23 April | + | + + | + | + + | + | + | ||||||
| 24 April | + + | + + | + + | + | + | + | + | |||||
| 25 April | + | + + | + | + | + | + | + | + | ||||
| 26 April | + | + + | + | + + | + | + | + | + | + | |||
| 27 April | + + | + + | + + | + + | + | |||||||
| 28 April | + | + | + | |||||||||
| 29 April | + | + | ||||||||||
| 30 April | + | + | ||||||||||
| 1 May | + | + | + | |||||||||
| 2 May | + | + | + | + | ||||||||
| 3 May | + | + | ||||||||||
| 4 May | ||||||||||||
| 5 May | + | + | + | + | + | |||||||
| 6 May | + | + | ||||||||||
| 7 May | + | + | + + | + + | + | + | + | |||||
| 8 May | + | + | + | + | + | + + | ||||||
| 9 May | + | + | + + | + + | + | |||||||
| 10 May | + | + | + | + | + | + | + + |
| Variable | Number of Days b | Odds Ratio (95% Confidence Interval) | p-Value (Fisher’s Exact Test) | |
|---|---|---|---|---|
| Output of infection risk model | Favourable and marginal meteorological conditions | 0.89 ± 0.10 | 2.48 (1.36–4.52) | 0.003 |
| Daily mean relative humidity (RH) | RH ≥ 90% | 0.21 ± 0.05 | 1.05 (0.40–2.75) | 0.924 |
| RH ≥ 85% | 0.43 ± 0.08 | 1.20 (0.65–2.22) | 0.561 | |
| RH ≥ 80% | 0.60 ± 0.09 | 1.60 (0.91–2.81) | 0.106 | |
| RH ≥ 75% | 0.97 ± 0.10 | 1.92 (1.14–3.25) | 0.015 | |
| Daily precipitation (Rain) | 0.5 mm ≤ Rain < 5.0 mm | 0.34 ± 0.05 | 0.85 (0.28–2.56) | 0.768 |
| 0.5 mm ≤ Rain < 20.0 mm | 0.67 ± 0.07 | 0.98 (0.44–2.17) | 0.963 | |
| 0.5 mm ≤ Rain < 40.0 mm | 0.88 ± 0.09 | 1.31 (0.74–2.34) | 0.353 | |
| 0.5 mm ≤ Rain | 0.95 ± 0.09 | 1.24 (0.71–2.15) | 0.450 |
| Treatment (Active Ingredients) | Date of Investigation (Fungicide Experiment 4, Conducted in 2017) | Date of Investigation (Fungicide Experiment 5, Conducted in 2018) | ||||||
|---|---|---|---|---|---|---|---|---|
| Disease Incidence (%) a | Disease Severity (%) a | Disease Incidence (%) | Disease Severity (%) | |||||
| 19 April | 25 April | 19 April | 25 April | 18 April | 26 April | 18 April | 26 April | |
| Mancozeb | 6.7 ± 3.5 | 9.3 ± 4.1 *,b | 1.3 ± 0.7 | 1.9 ± 0.8 * | 2.7 ± 0.7 ** | 21.3 ± 16.3 * | 0.5 ± 0.1 * | 4.4 ± 3.4 ** |
| Chlorothalonil +Metalaxyl-M | 10.7 ± 3.7 | 65.3 ± 15.7 | 2.1 ± 0.7 | 13.5 ± 3.4 | 40.7 ± 21.0 | 77.3 ± 16.7 | 8.5 ± 4.6 | 16.3 ± 3.6 |
| Control | 22.0 ± 14.0 | 76.0 ± 11.1 | 4.4 ± 2.8 | 17.1 ± 3.4 | 80.7 ± 4.4 | 100.0 ± 0.0 | 17.9 ± 2.1 | 28.5 ± 2.5 |
| Geographic Origin | Percent (%) of Germ-Tube Length Relative to That of Control (Total Number Measured) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Metalaxyl (µg ai/mL) | Metalaxyl-M (µg ai/mL) | ||||||||||||||
| 200 | 40 | 8 | 1.6 | 0.32 | 0 | EC50 a | 200 | 40 | 8 | 1.6 | 0.32 | 0.064 | 0 | EC50 | |
| Saga, Honjou | – b | 29.0 (12) | 59.7 (30) | 88.8 (13) | 109.9 (16) | 100 (30) | 13.1 | – | – | – | – | – | – | – | – |
| Saga, Kawasoe | 25.4 (30) | 23.2 (24) | 34.9 (30) | 38.3 (30) | 38.8 (30) | 100 (29) | <0.32 | 26.9 (30) | 33.2 (30) | 22.7 (30) | 33.8 (30) | 20.8 (30) | 58.5 (30) | 100 (29) | <0.32 |
| Saga, Kubota | – | – | – | – | 15.5 (9) | 100 (11) | <0.32 | – | 18.6 (6) | 26.6 (6) | 24.2 (7) | 30.6 (13) | 13.3 (5) | 100 (11) | <0.064 |
| Kashima 1 | – | 19.5 (24) | 39.3 (15) | 61.9 (22) | 73.6 (23) | 100 (30) | 3.1 | – | – | – | – | – | – | – | |
| Kashima 2 | – | 63.3 (31) | 120.0 (27) | 130.2 (14) | 133.2 (14) | 100 (10) | >40 | 75.1 (30) | 119.3 (18) | 106.6 (19) | 113.5 (28) | 122.7 (19) | 127.4 (23) | 100 (10) | >200 |
| Kashima 3 | 90.3 (30) | 120.0 (17) | 133.5 (23) | 138.8 (21) | 78.7 (29) | 100 (22) | >200 | 28.3 (30) | 118.7 (30) | 109.3 (23) | 132.1 (27) | 113.7 (30) | 116.5 (22) | 100 (22) | >40 |
| Kanzaki | 52.7 (30) | 71.1 (19) | 74.0 (17) | 82.6 (20) | 79.6 (21) | 100 (30) | >200 | – | – | – | – | – | – | – | – |
| Miyaki 1 | – | 85.7 (31) | 109.9 (12) | 120.9 (17) | 113.4 (22) | 100 (29) | >40 | – | – | – | – | – | – | – | – |
| Miyaki 2 | 107.8 (12) | 104.0 (17) | 110.0 (12) | 106.4 (18) | 99.7 (19) | 100 (21) | >200 | – | – | – | – | – | – | – | – |
| Shiroishi 1 | 56.7 (8) | 69.5 (4) | 56.8 (5) | 109.7 (7) | 149.9 (6) | 100 (7) | >200 | – | – | – | – | – | – | – | – |
| Shiroishi 2 | 70.4 (6) | 133.9 (6) | 124.5 (5) | 110.7 (4) | 139.5 (7) | 100 (5) | >200 | – | – | – | – | – | – | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Syobu, S.-i.; Watanabe, S. Characteristics of Meteorological Conditions during a Severe Outbreak of Onion Downy Mildew and Metalaxyl Sensitivity of Peronospora destructor in Saga, Japan, in 2016. Horticulturae 2022, 8, 578. https://doi.org/10.3390/horticulturae8070578
Syobu S-i, Watanabe S. Characteristics of Meteorological Conditions during a Severe Outbreak of Onion Downy Mildew and Metalaxyl Sensitivity of Peronospora destructor in Saga, Japan, in 2016. Horticulturae. 2022; 8(7):578. https://doi.org/10.3390/horticulturae8070578
Chicago/Turabian StyleSyobu, Shin-ichirou, and Sachiko Watanabe. 2022. "Characteristics of Meteorological Conditions during a Severe Outbreak of Onion Downy Mildew and Metalaxyl Sensitivity of Peronospora destructor in Saga, Japan, in 2016" Horticulturae 8, no. 7: 578. https://doi.org/10.3390/horticulturae8070578
APA StyleSyobu, S. -i., & Watanabe, S. (2022). Characteristics of Meteorological Conditions during a Severe Outbreak of Onion Downy Mildew and Metalaxyl Sensitivity of Peronospora destructor in Saga, Japan, in 2016. Horticulturae, 8(7), 578. https://doi.org/10.3390/horticulturae8070578