

Influence of Benzothiadiazole on the Amino Acids and Aroma Compositions of ‘Cabernet Gernischt’ Grapes (Vitis vinifera L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Study Site, Treatment, and Sampling
2.3. Determination of Physicochemical Properties
2.4. Amino Acids Analysis
2.5. Analysis of Grape Aroma Compounds
2.6. Odor Activity Values (OAVs)
2.7. Statistical Analysis
3. Results
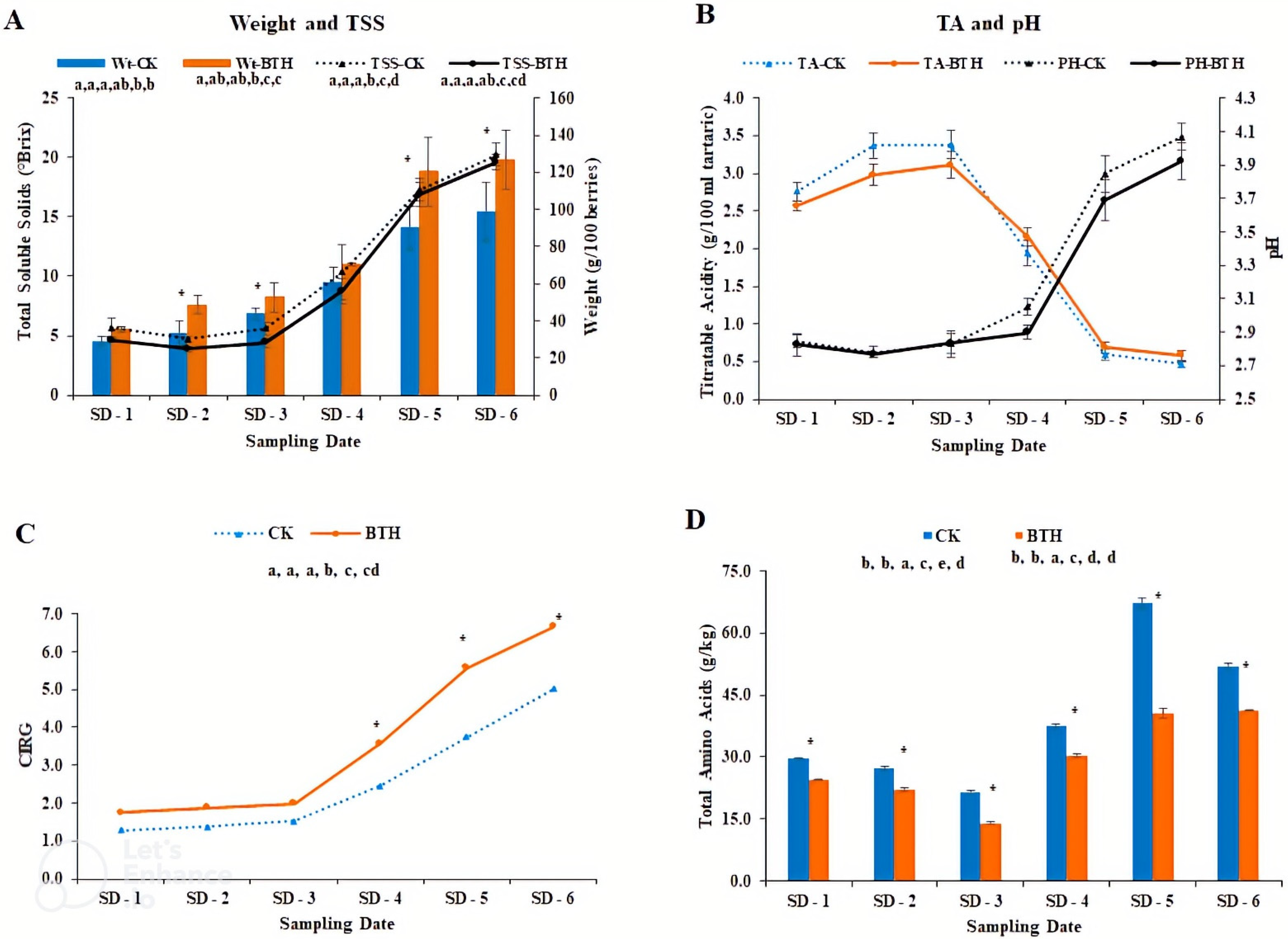
3.1. Physicochemical Parameters of ‘Cabernet Gernischt’ Grapes
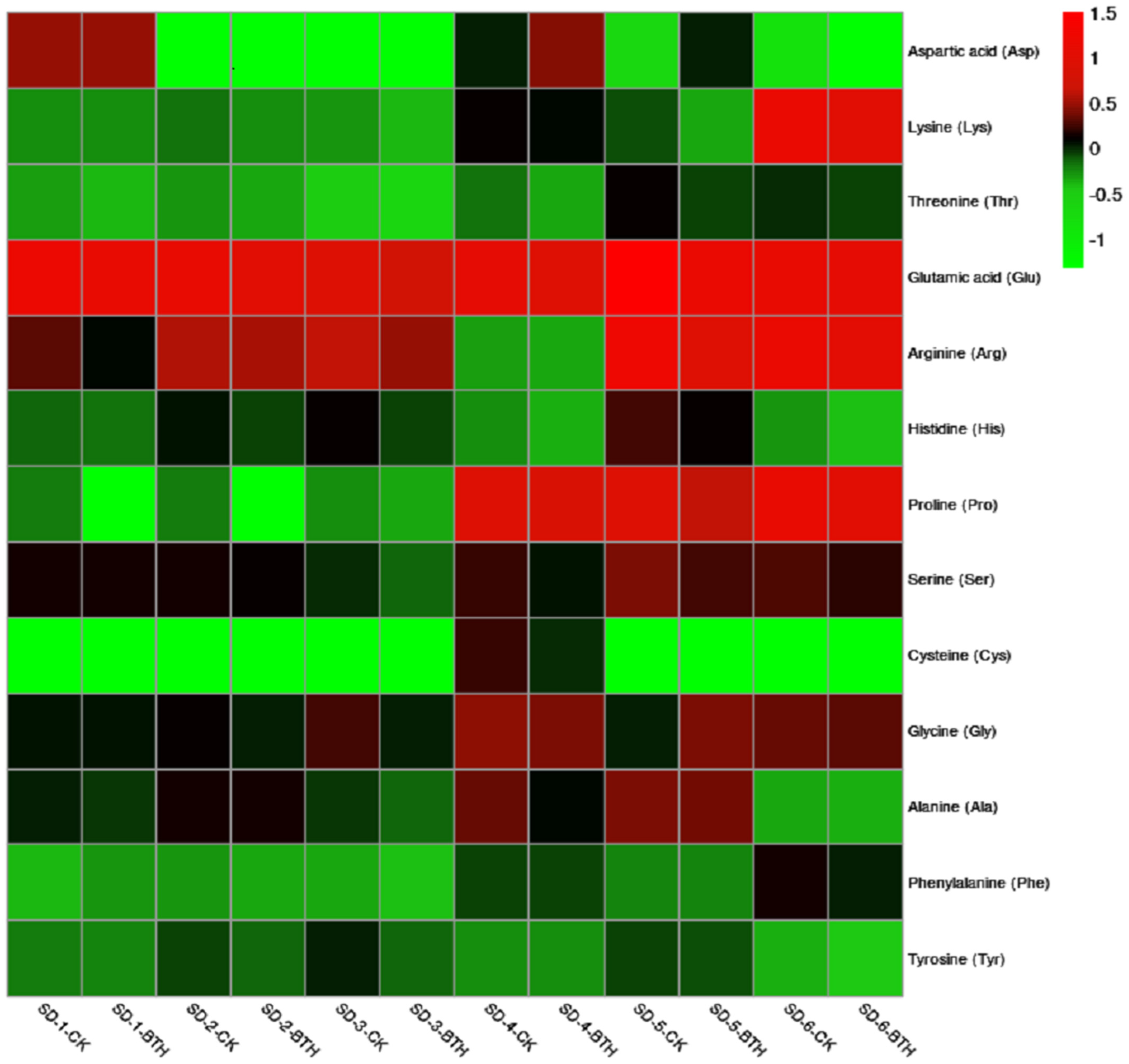
3.2. Amino Acids in ‘Cabernet Gernischt’ Grapes
3.3. Concentrations of Aroma Compounds in ‘Cabernet Gernischt’ Grapes
3.4. Odor Activity Values of Aroma Compounds
3.5. Correlation between Amino Acids and Amino Acids-Derived Aroma Compounds
4. Discussion
4.1. Effect of BTH on the Physicochemical Properties of ‘Cabernet Gernischt’ Grapes
4.2. Effect of BTH on the Amino Acids Profile of ‘Cabernet Gernischt’ Grapes
4.3. Effects of BTH on the Aroma Composition of ‘Cabernet Gernischt’ Grapes
4.4. Correlation Analysis of Amino Acids and Derived Aroma Compounds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salifu, R.; Chen, C.; Sam, F.E.; Jiang, Y. Application of Elicitors in Grapevine Defense: Impact on Volatile Compounds. Horticulturae 2022, 8, 451. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, Y.; Li, C.; Wang, B.; Ma, L.; Ren, Y.; Bi, Y.; Li, Y.; Xue, H.; Prusky, D. The effect of benzo-(1,2,3)-thiadiazole-7-carbothioic acid S-methyl ester (BTH) treatment on regulation of reactive oxygen species metabolism involved in wound healing of potato tubers during postharvest. Food Chem. 2020, 309, 125608. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Rossoni, M.; Borgo, M.; Ferrara, L.; Faoro, F. Induction of resistance to gray mold with benzothiadiazole modifies amino acid profile and increases proanthocyanidins in grape: Primary versus secondary metabolism. J. Agric. Food Chem. 2005, 53, 9133–9139. [Google Scholar] [CrossRef]
- Ge, Y.; Tang, Q.; Li, C.; Duan, B.; Li, X.; Wei, M.; Li, J. Acibenzolar-S-methyl treatment enhances antioxidant ability and phenylpropanoid pathway of blueberries during low temperature storage. LWT-Food Sci. Technol. 2019, 110, 48–53. [Google Scholar] [CrossRef]
- Miliordos, D.E.; Tsiknia, M.; Kontoudakis, N.; Dimopoulou, M.; Bouyioukos, C.; Kotseridis, Y. Impact of application of abscisic acid, benzothiadiazole and chitosan on berry quality characteristics and plant associated microbial communities of vitis vinifera l var. Mouhtaro plants. Sustainability 2021, 13, 5802. [Google Scholar] [CrossRef]
- Bektas, Y.; Eulgem, T. Synthetic plant defense elicitors. Front. Plant Sci. 2015, 5, 804. [Google Scholar] [CrossRef]
- Wang, K.; Liao, Y.; Cao, S.; Di, H.; Zheng, Y. Effects of benzothiadiazole on disease resistance and soluble sugar accumulation in grape berries and its possible cellular mechanisms involved. Postharvest Biol. Technol. 2015, 102, 51–60. [Google Scholar] [CrossRef]
- Poni, S.; Gatti, M.; Palliotti, A.; Dai, Z.; Duchêne, E.; Truong, T.T.; Ferrara, G.; Matarrese, A.M.S.; Gallotta, A.; Bellincontro, A.; et al. Grapevine quality: A multiple choice issue. Sci. Hortic. 2018, 234, 445–462. [Google Scholar]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.; Tavares, R.; Sousa, M.; Agasse, A.; Delrot, S.; Gerós, H. Biochemical changes throughout grape berry development and fruit and wine quality. Food 2007, 1, 1–22. [Google Scholar]
- Wu, Y.; Zhang, W.; Song, S.; Xu, W.; Zhang, C.; Ma, C.; Wang, L.; Wang, S. Evolution of volatile compounds during the development of Muscat grape ‘Shine Muscat’ (Vitis labrusca × V. vinifera). Food Chem. 2020, 309, 125778. [Google Scholar] [CrossRef]
- Gonda, I.; Bar, E.; Portnoy, V.; Lev, S.; Burger, J.; Schaffer, A.A.; Tadmor, Y.; Gepstein, S.; Giovannoni, J.J.; Katzir, N.; et al. Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L. fruit. J. Exp. Bot. 2010, 61, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Alem, H.; Rigou, P.; Schneider, R.; Ojeda, H.; Torregrosa, L. Impact of agronomic practices on grape aroma composition: A review. J. Sci. Food Agric. 2019, 99, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.S. Vineyard practice. In Wine Science; Academic Press: Cambridge, MA, USA, 2020; pp. 151–330. ISBN 9780128161180. [Google Scholar]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J. 2008, 54, 712–732. [Google Scholar] [CrossRef] [PubMed]
- Maoz, I.; Rikanati, R.D.; Schlesinger, D.; Bar, E.; Gonda, I.; Levin, E.; Kaplunov, T.; Sela, N.; Lichter, A.; Lewinsohn, E. Concealed ester formation and amino acid metabolism to volatile compounds in table grape (Vitis vinifera L.) berries. Plant Sci. 2018, 274, 223–230. [Google Scholar] [CrossRef]
- Kalua, C.M.; Boss, P.K. Evolution of volatile compounds during the development of cabernet sauvignon grapes (Vitis vinifera L.). J. Agric. Food Chem. 2009, 57, 3818–3830. [Google Scholar] [CrossRef]
- Paladines-Quezada, D.F.; Fernández-Fernández, J.I.; Moreno-Olivares, J.D.; Bleda-Sánchez, J.A.; Gómez-Martínez, J.C.; Martínez-Jiménez, J.A.; Gil-Muñoz, R. Application of elicitors in two ripening periods of Vitis vinifera L. Cv monastrell: Influence on anthocyanin concentration of grapes and wines. Molecules 2021, 26, 1689. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; Romero-Cascales, I.; Bautista-Ortín, A.B.; Gil-Muñoz, R.; Martínez-Cutillas, A.; Gómez-Plaza, E. Increasing bioactive phenolic compounds in grapes: Response of six monastrell grape clones to benzothiadiazole and methyl jasmonate treatments. Am. J. Enol. Vitic. 2013, 64, 459–465. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gómez-Plaza, E. Improving phenolic and chromatic characteristics of monastrell, merlot and syrah wines by using methyl jasmonate and benzothiadiazole. Oeno One 2017, 51, 17–27. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; Gil-Muñoz, R.; López-Roca, J.M.; Martínez-Cutillas, A.; Romero-Cascales, I.; Gómez-Plaza, E. Increasing the Phenolic Compound Content of Grapes by Preharvest Application of Abcisic Acid and a Combination of Methyl Jasmonate and Benzothiadiazole. J. Agric. Food Chem. 2013, 61, 3978–3983. [Google Scholar] [CrossRef]
- Paladines-Quezada, D.F.; Moreno-Olivares, J.D.; Fernández-Fernández, J.I.; Bautista-Ortín, A.B.; Gil-Muñoz, R. Influence of methyl jasmonate and benzothiadiazole on the composition of grape skin cell walls and wines. Food Chem. 2019, 277, 691–697. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; López-Roca, J.M.; Bautista-Ortín, A.B.; Gil-Muñoz, R.; Gómez-Plaza, E. Effect of combined use of benzothiadiazole and methyl jasmonate on volatile compounds of monastrell wine. Am. J. Enol. Vitic. 2014, 65, 238–243. [Google Scholar] [CrossRef]
- Gómez-Plaza, E.; Mestre-Ortuño, L.; Ruiz-García, Y.; Fernández-Fernández, J.I.; López-Roca, J.M. Effect of benzothiadiazole and methyl jasmonate on the volatile compound composition of Vitis vinifera L. Monastrell grapes and wines. Am. J. Enol. Vitic. 2012, 63, 394–401. [Google Scholar] [CrossRef]
- FAO and OIV Food and Agriculture Organization of the United Nations and the International Organisation of Vine and Wine 2016. 2016. p. 64. Available online: https://www.fao.org/publications/card/es/c/709ef071-6082-4434-91bf-4bc5b01380c6 (accessed on 10 March 2022).
- Kupe, M.; Ercisli, S.; Baron, M.; Sochor, J. Sustainable viticulture on traditional ‘baran’ training system in eastern turkey. Sustainability 2021, 13, 10236. [Google Scholar] [CrossRef]
- Taskesenlioglu, M.Y.; Ercisli, S.; Kupe, M.; Ercisli, N. History of Grape in Anatolia and Historical Sustainable Grape Production in Erzincan Agroecological Conditions in Turkey. Sustain. 2022, 14, 31496. [Google Scholar] [CrossRef]
- OIV International Organisation of Vine and Wine (OIV). OIV State of the World Vitivinicultural Sector in 2020; OIV International Organisation of Vine and Wine (OIV): Paris, France, 2021; pp. 1–19. [Google Scholar]
- Thakur, M.; Sohal, B.S. Role of Elicitors in Inducing Resistance in Plants against Pathogen Infection: A Review. ISRN Biochem. 2013, 2013, 762412. [Google Scholar] [CrossRef]
- Fan, W.; Xu, Y.; Jiang, W.; Li, J. Identification and quantification of impact aroma compounds in 4 nonfloral Vitis vinifera varieties grapes. J. Food Sci. 2010, 75, S81–S88. [Google Scholar] [CrossRef]
- Sun, W.X.; Hu, K.; Zhang, J.X.; Zhu, X.L.; Tao, Y.S. Aroma modulation of Cabernet Gernischt dry red wine by optimal enzyme treatment strategy in winemaking. Food Chem. 2018, 245, 1248–1256. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Li, J.; Xu, Y. Different influences of β-glucosidases on volatile compounds and anthocyanins of Cabernet Gernischt and possible reason. Food Chem. 2013, 140, 245–254. [Google Scholar] [CrossRef]
- Xie, S.; Tang, Y.; Wang, P.; Song, C.; Duan, B.; Zhang, Z.; Meng, J. Influence of natural variation in berry size on the volatile profiles of Vitis vinifera L. Cv. Merlot and Cabernet Gernischt grapes. PLoS ONE 2018, 13, e0201374. [Google Scholar] [CrossRef]
- Gómez-Plaza, E.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gil-Muñoz, R. Effect of elicitors on the evolution of grape phenolic compounds during ripening period. J. Sci. Food Agric. 2017, 3, 977–983. [Google Scholar] [CrossRef]
- OIV International Organisation of Vine and Wine (OIV). Compendium of International Methods of Wine and Must Analysis; OIV International Organisation of Vine and Wine (OIV): Paris, France, 2016; Volume 1. [Google Scholar]
- Yue, X.; Ju, Y.; Tang, Z.; Zhao, Y.; Jiao, X.; Zhang, Z. Effects of the severity and timing of basal leaf removal on the amino acids profiles of Sauvignon Blanc grapes and wines. J. Integr. Agric. 2019, 18, 2052–2062. [Google Scholar] [CrossRef]
- Koyama, R.; Roberto, S.R.; de Souza, R.T.; Borges, W.F.S.; Anderson, M.; Waterhouse, A.L.; Cantu, D.; Fidelibus, M.W.; Blanco-Ulate, B. Exogenous abscisic acid promotes anthocyanin biosynthesis and increased expression of flavonoid synthesis genes in Vitis vinifera × Vitis labrusca table grapes in a subtropical region. Front. Plant Sci. 2018, 9, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Xu, J.; Chen, H.; Zhang, H.; Li, S. Determination of Volatile Substances and Amino Acids in Yali Pear Fruit. Food Sci. Technol. 2002, 71–73. [Google Scholar] [CrossRef]
- Burdock, G.A. Fenaroli’s Handbook of Flavor Ingredients, 6th ed.; CRC Press: Abingdon, UK, 2005; ISBN 0849330343. [Google Scholar]
- Pino, J.A.; Quijano, C.E. Study of the volatile compounds from plum (Prunus domestica L. cv. Horvin) and estimation of their contribution to the fruit aroma. Food Sci. Technol. 2012, 32, 76–83. [Google Scholar] [CrossRef]
- Ferreira, V.; López, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; Romero-Cascales, I.; Gil-Muñoz, R.; Fernández-Fernández, J.I.; López-Roca, J.M.; Gómez-Plaza, E. Improving grape phenolic content and wine chromatic characteristics through the use of two different elicitors: Methyl jasmonate versus benzothiadiazole. J. Agric. Food Chem. 2012, 60, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Coombe, B.G.; Bishop, G.R. Development of the grape berry. II * Changes in diameter and deformability during veraison. Aust. J. Agric. Res. 1980, 31, 499–509. [Google Scholar] [CrossRef]
- Paladines-Quezada, D.F.; Moreno-Olivares, J.D.; Fernández-Fernández, J.I.; Bleda-Sánchez, J.A.; Martínez-Moreno, A.; Gil-Muñoz, R. Elicitors and pre-fermentative cold maceration: Effects on polyphenol concentration in monastrell grapes and wines. Biomolecules 2019, 9, 9110671. [Google Scholar] [CrossRef]
- Fernandez-Marin, M.I.; Guerrero, R.F.; Puertas, B.; Garcia-Parrilla, M.C.; Collado, I.G.; Cantos-Villar, E. Impact of preharvest and postharvest treatment combinations on increase of stilbene content in grape. J. Int. Sci. Vigne Vin 2013, 47, 203–212. [Google Scholar] [CrossRef]
- Beckers, G.J.M.; Spoel, S.H. Fine-tuning plant defence signalling: Salicylate versus jasmonate. Plant Biol. 2006, 8, 1–10. [Google Scholar] [CrossRef]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Filipe-Ribeiro, L.; Mendes-Faia, A. Validation and comparison of analytical methods used to evaluate the nitrogen status of grape juice. Food Chem. 2007, 3, 1272–1277. [Google Scholar] [CrossRef]
- Guan, L.; Wu, B.; Hilbert, G.; Li, S.; Gomès, E.; Delrot, S.; Dai, Z. Cluster shading modifies amino acids in grape (Vitis vinifera L.) berries in a genotype- and tissue-dependent manner. Food Res. Int. 2017, 98, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gamboa, G.; Pérez-Álvarez, E.P.; Rubio-Bretón, P.; Garde-Cerdán, T. Foliar application of methyl jasmonate to Graciano and Tempranillo vines: Effects on grape amino acid content during two consecutive vintages. Oeno One 2019, 53, 1–19. [Google Scholar] [CrossRef]
- Lalonde, S.; Tegeder, M.; Throne-Holst, M.; Frommer, W.B.; Patrick, J.W. Phloem loading and unloading of sugars and amino acids. Plant Cell Environ. 2003, 26, 37–56. [Google Scholar] [CrossRef]
- Kliewer, W.M. Changes in the concentration of free amino acids in grape berries during maturation. Am. J. Enol. Vitic. 1968, 19, 166–174. [Google Scholar]
- Gil-Muñoz, R.; Giménez-Bañón, M.J.; Moreno-Olivares, J.D.; Paladines-Quezada, D.F.; Bleda-Sánchez, J.A.; Fernández-Fernández, J.I.; Parra-Torrejón, B.; Ramírez-Rodríguez, G.B.; Delgado-López, J.M. Effect of methyl jasmonate doped nanoparticles on nitrogen composition of monastrell grapes and wines. Biomolecules 2021, 11, 1631. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Portu, J.; Santamaría, P.; López, R.; Garde-Cerdán, T. Effects on grape amino acid concentration through foliar application of three different elicitors. Food Res. Int. 2017, 99, 688–692. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Lorenzo, C.; Lara, J.F.; Prado, F.; Ancín-Azplicueta, C.; Salinas, M.R. Study of the Evolution of Nitrogen Compounds during Grape Ripening. Application to Differentiate. J. Agric. Food Chem. 2009, 57, 2410–2419. [Google Scholar] [CrossRef]
- Robinson, A.L.; Boss, P.K.; Solomon, P.S.; Trengove, R.D.; Heymann, H.; Ebeler, S.E. Origins of Grape and Wine Aroma. Part 1. Chemical Components and Viticultural Impacts. Am. J. Enol. Vitic. 2014, 65, 1–24. [Google Scholar] [CrossRef]
- El Hadi, M.A.M.; Zhang, F.J.; Wu, F.F.; Zhou, C.H.; Tao, J. Advances in fruit aroma volatile research. Molecules 2013, 18, 8200–8229. [Google Scholar] [CrossRef] [PubMed]
- Aragüez, I.; Valpuesta Fernández, V. Metabolic engineering of aroma components in fruits. Biotechnol. J. 2013, 8, 1144–1158. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.G.; Olías, R.; Luaces, P.; Sanz, C. Biosynthesis of strawberry aroma compounds through amino acid metabolism. J. Agric. Food Chem. 2002, 50, 4037–4042. [Google Scholar] [CrossRef]
- Rosati, C.; Diretto, G.; Giuliano, G. Biosynthesis and engineering of carotenoids and apocarotenoids in plants: State of the art and future prospects. Biotechnol. Genet. Eng. Rev. 2009, 26, 139–162. [Google Scholar] [CrossRef] [PubMed]
- García, E.; Chacón, J.L.; Martínez, J.; Izquierdo, P.M. Changes in volatile compounds during ripening in grapes of Airén, Macabeo and Chardonnay white varieties grown in La Mancha region (Spain). Food Sci. Technol. Int. 2003, 9, 33–41. [Google Scholar] [CrossRef]
- Vitalini, S.; Ruggiero, A.; Rapparini, F.; Neri, L.; Tonni, M.; Iriti, M. The application of chitosan and benzothiadiazole in the vineyard (Vitis vinífera L. cv Groppello Gentile) changes the aromatic profile and sensory attributes of wine. Food Chem. 2014, 162, 192–205. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Perestrelo, R.; Fernandes, A.; Albuquerque, F.F.; Marques, J.C.; Câmara, J.S. Analytical characterization of the aroma of Tinta Negra Mole red wine: Identification of the main odorants compounds. Anal. Chim. Acta 2006, 563, 154–164. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Garde-Cerdán, T.; Rubio-Bretón, P.; Pérez-Álvarez, E.P. Seaweed foliar applications at two dosages to Tempranillo blanco (Vitis vinifera L.) grapevines in two seasons: Effects on grape and wine volatile composition. Food Res. Int. 2020, 130, 108918. [Google Scholar] [CrossRef]
- Pott, D.M.; Osorio, S.; Vallarino, J.G. From central to specialized metabolism: An overview of some secondary compounds derived from the primary metabolism for their role in conferring nutritional and organoleptic characteristics to fruit. Front. Plant Sci. 2019, 10, 835. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Kessler, A.; Baldwin, I.T. Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gamboa, G.; Pérez-Álvarez, E.P.; Rubio-Bretón, P.; Garde-Cerdán, T. Changes on Grape Volatile Composition through Elicitation with Methyl Jasmonate, Chitosan, and a Yeast Extract in Tempranillo (Vitis vinifera L.) Grapevines; Elsevier: Amsterdam, The Netherlands, 2019; Volume 244, pp. 257–262. [Google Scholar]
- Marais, J.; Wyk, C.; Rapp, A. Effect of sunlight and shade on norisoprenoid levels in maturing Weisser Riesling and Chenin blanc grapes and Weisser Riesling wines. S. Afr. J. Enol. Vitic. Stellenbosch 1992, 13, 23–32. [Google Scholar] [CrossRef]
- Martin, D.M.; Gershenzon, J.; Bohlmann, J. Induction of volatile terpene biosynthesis and diurnal emission by methyl jasmonate in foliage of Norway spruce. Plant Physiol. 2003, 132, 21196. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Odor Activity Value (OAV) | Odor Threshold (µg/L) | Odor Descriptor | |
|---|---|---|---|---|
| Control | BTH | |||
| Hexenal | 13.30 ± 0.03 a | 22.13 ± 0.17 b | 4.5 | grassy, green |
| (E)-2-Hexenal | 2.50 ± 0.01 b | 0.29 ± 0.06 a | 17 | green, fresh, fruity |
| (E)-2-Hexenol | 0.22 ± 0.01 a | 0.23 ± 0.03 a | 100 | green |
| (Z)-3-Hexen-1-ol | 0.10 ± 0.01 b | 0.03 ± 0.01 a | 70 | green |
| (E)-2-Octenal | 0.07 ± 0.01 a | 0.19 ± 0.03 b | 3 | green, nut |
| Nonanal | 0.44 ± 0.02 a | 0.88 ± 0.05 b | 1 | citrusy, green |
| (Z)-2-Heptenal | 0.23 ± 0.01 a | 0.25 ± 0.03 a | 3 | dry fish, smoky |
| (E, Z)-2,6-Nonadienal | 8.50 ± 0.02 a | 10.00 ± 0.03 b | 0.02 | green, cucumber |
| 1-Octanol | 0.03 ± 0.00 a | 0.10 ± 0.03 b | 40 | floral, fatty |
| 1-Octen-3-ol | 1.54 ± 0.04 a | 1.95 ± 0.08 b | 1 | mushroom |
| Hexyl acetate | 0.03 ± 0.01 a | 0.10 ± 0.00 b | 2 | fruity |
| Hex-(3Z)-enyl butyrate | 0.04 ± 0.00 a | 0.05 ± 0.01 a | 20 | wine, green |
| (3Z)-3-Hexen-1-yl acetate | 0.37 ± 0.02 a | 0.55 ± 0.03 b | 8 | green, banana |
| (3Z)-3-Hexen-1-yl 3-methylbutanoate | 0.89 ± 0.05 b | 0.09 ± 0.02 a | 1 | fruity |
| β-ionone | 0.57 ± 0.05 a | 2.71 ± 0.07 b | 0.07 | floral, violet |
| Linalool | 0.02 ± 0.00 a | 0.04 ± 0.00 a | 25 | floral |
| β-Cyclocitral | 0.06 ± 0.01 a | 0.14 ± 0.02 b | 5 | minty |
| Terpinen-4-ol | 0.02 ± 0.00 a | 0.03 ± 0.00 a | 5 | nutmeg, floral, |
| β-damascenone | 1.78 ± 0.03 b | 1.33 ± 0.05 a | 0.09 | sweet, floral, fruity |
| TREATMENT | ASP | THR | SER | GLU | GLY | HIS | ALA | LYS | TYR | PHE | ARG | PRO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | ||||||||||||
| Benzaldehyde | −0.148 | 0.613 | 0.729 | 0.77 | −0.144 | 0.658 | −0.067 | 0.95 ** | 0.411 | 0.865 * | 0.559 | 0.058 |
| 2-Ethylhexanol | −0.478 | −0.012 | −0.183 | −0.529 | 0.655 | 0.256 | −0.182 | −0.312 | −0.199 | −0.253 | 0.221 | 0.48 |
| Benzyl alcohol | −0.369 | 0.419 | 0.241 | −0.058 | 0.268 | 0.271 | −0.593 | −0.282 | −0.694 | 0.898 * | 0.509 | −0.173 |
| Phenylethyl Alcohol | −0.285 | −0.045 | −0.348 | 0.863 * | −0.37 | 0.776 | −0.665 | 0.714 | 0.896 * | 0.939 ** | 0.847 * | 0.572 |
| (6S)-6-Methyl-1-octanol | −0.539 | 0.527 | 0.61 | 0.577 | −0.166 | 0.619 | −0.025 | 0.637 | 0.39 | 0.534 | 0.461 | 0.103 |
| 3-Methyl-3-heptanol | −0.141 | −0.013 | 0.041 | −0.372 | 0.799 | 0.019 | 0.62 | 0.927 ** | 0 | 0.334 | 0.024 | −0.003 |
| Phenol | 0.058 | −0.15 | −0.086 | −0.447 | 0.924 ** | 0.028 | 0.701 | 0.306 | 0.201 | 0.315 | −0.041 | 0.864 * |
| Styrene | −0.251 | 0.175 | 0.213 | 0.438 | −0.714 | 0.035 | −0.221 | −0.046 | 0.061 | 0.196 | 0.006 | −0.033 |
| BTH | ASP | THR | SER | GLU | GLY | HIS | ALA | LYS | TYR | PHE | ARG | PRO |
| Benzaldehyde | 0.562 | −0.156 | −0.161 | −0.112 | 0.58 | 0.563 | 0.47 | 0.912 * | 0.859 * | 0.235 | 0.142 | −0.403 |
| 2-Ethylhexanol | −0.053 | −0.294 | −0.471 | −0.487 | 0.282 | 0.495 | 0.53 | 0.401 | 0.847 * | 0.134 | 0.04 | −0.452 |
| Benzyl alcohol | 0.031 | 0.563 | 0.532 | 0.498 | 0.642 | 0.755 | 0.7 | 0.133 | 0.567 | 0.873 * | 0.49 | 0.047 |
| Phenylethyl Alcohol | −0.381 | 0.757 | 0.584 | 0.418 | 0.605 | 0.773 | 0.592 | −0.102 | 0.224 | −0.892 * | 0.714 | 0.422 |
| (6S)-6-Methyl-1-octanol | 0.267 | 0.265 | 0.228 | 0.233 | 0.69 | 0.779 | 0.851 * | 0.472 | 0.83 * | 0.621 | 0.366 | −0.204 |
| 3-Methyl-3-heptanol | 0.428 | −0.105 | −0.135 | −0.127 | 0.531 | 0.522 | 0.313 | 0.172 | 0.718 | 0.034 | 0.203 | −0.276 |
| Phenol | −0.376 | 0.575 | 0.38 | 0.125 | −0.812 * | 0.118 | −0.705 | −0.203 | 0.21 | −0.557 | 0.613 | 0.934 ** |
| Styrene | −0.744 | 0.071 | −0.285 | −0.56 | −0.029 | 0.105 | −0.656 | −0.061 | −0.413 | 0.387 | 0.387 | 0.542 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salifu, R.; Jiang, Y.; Ba, L.; Zhang, Z.; Feng, L.; Li, J. Influence of Benzothiadiazole on the Amino Acids and Aroma Compositions of ‘Cabernet Gernischt’ Grapes (Vitis vinifera L.). Horticulturae 2022, 8, 812. https://doi.org/10.3390/horticulturae8090812
Salifu R, Jiang Y, Ba L, Zhang Z, Feng L, Li J. Influence of Benzothiadiazole on the Amino Acids and Aroma Compositions of ‘Cabernet Gernischt’ Grapes (Vitis vinifera L.). Horticulturae. 2022; 8(9):812. https://doi.org/10.3390/horticulturae8090812
Chicago/Turabian StyleSalifu, Rafia, Yumei Jiang, Lingzhen Ba, Zhen Zhang, Lidan Feng, and Jixin Li. 2022. "Influence of Benzothiadiazole on the Amino Acids and Aroma Compositions of ‘Cabernet Gernischt’ Grapes (Vitis vinifera L.)" Horticulturae 8, no. 9: 812. https://doi.org/10.3390/horticulturae8090812
APA StyleSalifu, R., Jiang, Y., Ba, L., Zhang, Z., Feng, L., & Li, J. (2022). Influence of Benzothiadiazole on the Amino Acids and Aroma Compositions of ‘Cabernet Gernischt’ Grapes (Vitis vinifera L.). Horticulturae, 8(9), 812. https://doi.org/10.3390/horticulturae8090812