
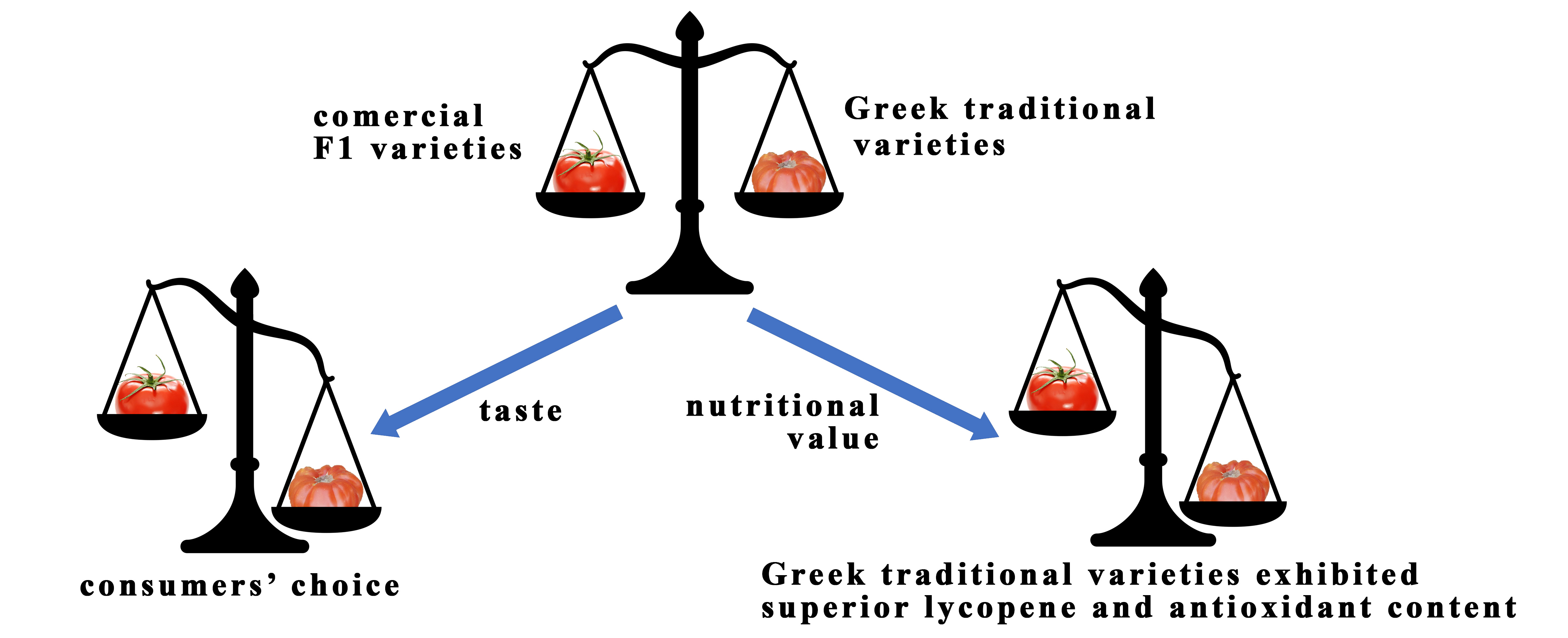
Comparison of Nutrient Composition and Antioxidant Activity of Hydroponically Grown Commercial and Traditional Greek Tomato Cultivars

 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Samples Preparation
- (i)
- Breaker (no more than 10% of the surface was red or pink),
- (ii)
- Red ripe (more than 90% of surface was red).
2.3. Nutrient Composition
2.3.1. Total Fat
2.3.2. Proteins
2.3.3. Carbohydrates
Preparation of Extract
Anthrone Assay
2.4. Antioxidants Composition
2.4.1. Total Phenols and Flavonoids Content
Preparation of Extract
Total Phenols Analysis
2.4.2. Lycopene Analysis
2.4.3. Ascorbic acid Analysis
2.5. Antioxidant Activity
2.5.1. Preparation of Extract
2.5.2. DPPH Assay
2.5.3. FRAP Assay
2.6. Statistical Analyses
3. Results and Discussion
3.1. Nutrition Value
3.2. Antioxidants Content
3.2.1. Ascorbic acid (AsA)
3.2.2. Lycopene
3.2.3. Total Phenolic and Flavonoids Compounds
3.3. Antioxidant Activity
3.3.1. Hydrophilic Antioxidant Activity (HAA)
3.3.2. Lipophilic Antioxidant Activity (LAA)
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Siddiqui, M.W.; Ayala-Zavala, J.F.; Dhua, R.S. Genotypic Variation in Tomatoes Affecting Processing and Antioxidant Attributes. Crit. Rev. Food Sci. Nutr. 2015, 55, 1819–1835. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.L.T.; Nguyen, C.V.; Hill, T.; Cheng, K.L.; Figueroa-Balderas, R.; Aktas, H.; Ashrafi, H.; Pons, C.; Fernández-Muñoz, R.; Vicente, A.; et al. Uniform ripening Encodes a Golden 2-like Transcription Factor Regulating Tomato Fruit Chloroplast Development. Science 2012, 336, 1711–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenucci, M.S.; Cadinu, D.; Taurino, M.; Piro, G.; Dalessandro, G. Antioxidant Composition in Cherry and High-Pigment Tomato Cultivars. J. Agric. Food Chem. 2006, 54, 2606–2613. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef] [Green Version]
- De Stefani, E.; Oreggia, F.; Boffetta, P.; Deneo-Pellegrini, H.; Ronco, A.; Mendilaharsu, M. Tomatoes, tomato-rich foods, lycopene and cancer of the upper aerodigestive tract: A case-control in Uruguay. Oral Oncol. 2000, 36, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Reimers, K.; Andon, M.; Jacobson, M.; Nguyen, V.; Rippe, J. Relationship of the tomato intake amount recommended in MyPlate to cardiovascular disease risk reduction (630.1). FASEB J. 2014, 28, 630–631. [Google Scholar] [CrossRef]
- Turati, F.; Rossi, M.; Pelucchi, C.; Levi, F.; La Vecchia, C. Fruit and vegetables and cancer risk: A review of southern European studies. Br. J. Nutr. 2015, 113, S102–S110. [Google Scholar] [CrossRef]
- Arab, L.; Steck, S. Lycopene and cardiovascular disease. Am. J. Clin. Nutr. 2000, 71, 1691S–1695S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assar, E.A.; Vidalle, M.C.; Chopra, M.; Hafizi, S. Lycopene acts through inhibition of IκB kinase to suppress NF-κB signaling in human prostate and breast cancer cells. Tumor Biol. 2016, 37, 9375–9385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gann, P.H.; Ma, J.; Giovannucci, E.; Willett, W.; Sacks, F.M.; Hennekens, C.H.; Stampfer, M.J. Lower Prostate Cancer Risk in Men with Elevated Plasma Lycopene Levels. Cancer Res. 1999, 59, 1225. [Google Scholar] [PubMed]
- Yuan, J.-M.; Ross, R.K.; Gao, Y.-T.; Qu, Y.-H.; Chu, X.-D.; Yu, M.C. Prediagnostic Levels of Serum Micronutrients in Relation to Risk of Gastric Cancer in Shanghai, China. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1772. [Google Scholar] [CrossRef]
- Ganji, V.; Kafai, M.R. Population Determinants of Serum Lycopene Concentrations in the United States: Data from the Third National Health and Nutrition Examination Survey, 1988–1994. J. Nutr. 2005, 135, 567–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, A.V.; Rao, L.G.; Young, G.L. Lycopene and Tomatoes in Human Nutrition and Health, 1st ed.; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Coyago-Cruz, E.; Corell, M.; Moriana, A.; Hernanz, D.; Benítez-González, A.M.; Stinco, C.M.; Meléndez-Martínez, A.J. Antioxidants (carotenoids and phenolics) profile of cherry tomatoes as influenced by deficit irrigation, ripening and cluster. Food Chem. 2018, 240, 870–884. [Google Scholar] [CrossRef] [Green Version]
- Azodanlou, R.; Darbellay, C.; Luisier, J.-L.; Villettaz, J.-C.; Amadò, R. Development of a model for quality assessment of tomatoes and apricots. LWT Food Sci. Technol. 2003, 36, 223–233. [Google Scholar] [CrossRef]
- Piombino, P.; Sinesio, F.; Moneta, E.; Cammareri, M.; Genovese, A.; Lisanti, M.T.; Mogno, M.R.; Peparaio, M.; Termolino, P.; Moio, L.; et al. Investigating physicochemical, volatile and sensory parameters playing a positive or a negative role on tomato liking. Food Res. Int. 2013, 50, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Causse, M.; Friguet, C.; Coiret, C.; Lépicier, M.; Navez, B.; Lee, M.; Holthuysen, N.; Sinesio, F.; Moneta, E.; Grandillo, S. Consumer Preferences for Fresh Tomato at the European Scale: A Common Segmentation on Taste and Firmness. J. Food Sci. 2010, 75, S531–S541. [Google Scholar] [CrossRef]
- Tieman, D.; Zhu, G.; Resende, M.F.R.; Lin, T.; Nguyen, C.; Bies, D.; Rambla, J.L.; Beltran, K.S.O.; Taylor, M.; Zhang, B.; et al. A chemical genetic roadmap to improved tomato flavor. Science 2017, 355, 391. [Google Scholar] [CrossRef] [PubMed]
- Sinesio, F.; Cammareri, M.; Cottet, V.; Fontanet, L.; Jost, M.; Moneta, E.; Palombieri, S.; Peparaio, M.; Romero del Castillo, R.; Saggia Civitelli, E.; et al. Sensory Traits and Consumer’s Perceived Quality of Traditional and Modern Fresh Market Tomato Varieties: A Study in Three European Countries. Foods 2021, 10, 2521. [Google Scholar] [CrossRef] [PubMed]
- Darras, A.I.; Kotsiras, A.; Delis, C.; Nifakos, K.; Pavlakos, E.; Demopoulos, V. Reaction of the native Greek tomato varieties ‘Chondrokatsari Messinias’ and ‘Katsari Santorinis’ to Fusarium oxysporum f. sp. lycopersici and Rhizoctonia solani infection. Hell. Plant Prot. J. 2017, 10, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Snyder, J.C.; Desborough, S.L. Rapid estimation of potato tuber total protein content with coomassie brilliant blue G-250. Theor. Appl. Genet. 1978, 52, 135–139. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Rebolloso-Fuentes, M.M. Nutrient composition and antioxidant activity of eight tomato (Lycopersicon esculentum) varieties. J. Food Compos. Anal. 2009, 22, 123–129. [Google Scholar] [CrossRef]
- Capanoglu, E.; Beekwilder, J.; Boyacioglu, D.; Hall, R.; de Vos, R. Changes in Antioxidant and Metabolite Profiles during Production of Tomato Paste. J. Agric. Food Chem. 2008, 56, 964–973. [Google Scholar] [CrossRef]
- Silva, L.; Pezzini, B.; Soares, L. Spectrophotometric determination of the total flavonoid content in Ocimum basilicum L. (Lamiaceae) leaves. Pharmacogn. Mag. 2015, 11, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.V.; Waseem, Z.; Agarwal, S. Lycopene content of tomatoes and tomato products and their contribution to dietary lycopene. Food Res. Int. 1998, 31, 737–741. [Google Scholar] [CrossRef]
- Klein, B.P.; Perry, A.K. Ascorbic Acid and Vitamin A Activity in Selected Vegetables from Different Geographical Areas of the United States. J. Food Sci. 1982, 47, 941–945. [Google Scholar] [CrossRef]
- Shi, J.; Qu, Q.; Kakuda, Y.; Xue, S.J.; Jiang, Y.; Koide, S.; Shim, Y.-Y. Investigation of the antioxidant and synergistic activity of lycopene and other natural antioxidants using LAME and AMVN model systems. J. Food Compos. Anal. 2007, 20, 603–608. [Google Scholar] [CrossRef]
- Figàs, M.R.; Prohens, J.; Raigón, M.D.; Fita, A.; García-Martínez, M.D.; Casanova, C.; Borràs, D.; Plazas, M.; Andújar, I.; Soler, S. Characterization of composition traits related to organoleptic and functional quality for the differentiation, selection and enhancement of local varieties of tomato from different cultivar groups. Food Chem. 2015, 187, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Pinela, J.; Barros, L.; Carvalho, A.M.; Ferreira, I.C.F.R. Nutritional composition and antioxidant activity of four tomato (Lycopersicon esculentum L.) farmer’ varieties in Northeastern Portugal homegardens. Food Chem. Toxicol. 2012, 50, 829–834. [Google Scholar] [CrossRef]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.-J. Protein Determination—Method Matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Kim, D.-S.; Kozukue, N.; Kim, H.-J.; Nishitani, Y.; Mizuno, M.; Levin, C.E.; Friedman, M. Protein, free amino acid, phenolic, β-carotene, and lycopene content, and antioxidative and cancer cell inhibitory effects of 12 greenhouse-grown commercial cherry tomato varieties. J. Food Compos. Anal. 2014, 34, 115–127. [Google Scholar] [CrossRef]
- Vinha, A.F.; Alves, R.C.; Barreira, S.V.P.; Castro, A.; Costa, A.S.G.; Oliveira, M.B.P.P. Effect of peel and seed removal on the nutritional value and antioxidant activity of tomato (Lycopersicon esculentum L.) fruits. LWT Food Sci. Technol. 2014, 55, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Ilahy, R.; Hdider, C.; Lenucci, M.S.; Tlili, I.; Dalessandro, G. Antioxidant activity and bioactive compound changes during fruit ripening of high-lycopene tomato cultivars. J. Food Compos. Anal. 2011, 24, 588–595. [Google Scholar] [CrossRef]
- Kaur, C.; Walia, S.; Nagal, S.; Walia, S.; Singh, J.; Singh, B.B.; Saha, S.; Singh, B.; Kalia, P.; Jaggi, S.; et al. Functional quality and antioxidant composition of selected tomato (Solanum lycopersicon L) cultivars grown in Northern India. LWT Food Sci. Technol. 2013, 50, 139–145. [Google Scholar] [CrossRef]
- Gould, W.A. Tomato Production, Processing, and Technology, 3rd ed.; Woodhead Publishing: Sawston, UK, 1992. [Google Scholar]
- Gautier, H.; Diakou-Verdin, V.; Bénard, C.; Reich, M.; Buret, M.; Bourgaud, F.; Poëssel, J.L.; Caris-Veyrat, C.; Génard, M. How Does Tomato Quality (Sugar, Acid, and Nutritional Quality) Vary with Ripening Stage, Temperature, and Irradiance? J. Agric. Food Chem. 2008, 56, 1241–1250. [Google Scholar] [CrossRef]
- Erba, D.; Casiraghi, M.C.; Ribas-Agustí, A.; Cáceres, R.; Marfà, O.; Castellari, M. Nutritional value of tomatoes (Solanum lycopersicum L.) grown in greenhouse by different agronomic techniques. J. Food Compos. Anal. 2013, 31, 245–251. [Google Scholar] [CrossRef]
- Nour, V.; Trandafir, I.; Ionica, E.M. Antioxidant Compounds, Mineral Content and Antioxidant Activity of Several Tomato Cultivars Grown in Southwestern Romania. Not. Bot. Horti Agrobot. Cluj Napoca 2013, 41, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, S.R.; Chae, Y.; Lee, J.G. Assessment of Phytochemicals, Quality Attributes, and Antioxidant Activities in Commercial Tomato Cultivars. Hortic. Sci. Technol. 2016, 34, 677–691. [Google Scholar] [CrossRef]
- Aoun, A.B.; Lechiheb, B.; Benyahya, L.; Ferchichi, A. Evaluation of fruit quality traits of traditional varieties of tomato (Solanum lycopersicum) grown in Tunisia. Afr. J. Food Sci. 2013, 7, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Ilahy, R.; Hdider, C.; Lenucci, M.S.; Tlili, I.; Dalessandro, G. Phytochemical composition and antioxidant activity of high-lycopene tomato (Solanum lycopersicum L.) cultivars grown in Southern Italy. Sci. Hortic. 2011, 127, 255–261. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Cho, M.-C.; Lee, J.G. Genotypic variation in carotenoid, ascorbic acid, total phenolic, and flavonoid contents, and antioxidant activity in selected tomato breeding lines. Hortic. Environ. Biotechnol. 2016, 57, 440–452. [Google Scholar] [CrossRef]
- Kotíková, Z.; Lachman, J.; Hejtmánková, A.; Hejtmánková, K. Determination of antioxidant activity and antioxidant content in tomato varieties and evaluation of mutual interactions between antioxidants. LWT Food Sci. Technol. 2011, 44, 1703–1710. [Google Scholar] [CrossRef]
- Adato, A.; Mandel, T.; Mintz-Oron, S.; Venger, I.; Levy, D.; Yativ, M.; Domínguez, E.; Wang, Z.; De Vos, R.C.H.; Jetter, R.; et al. Fruit-Surface Flavonoid Accumulation in Tomato Is Controlled by a SlMYB12-Regulated Transcriptional Network. PLoS Genet. 2009, 5, e1000777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vela-Hinojosa, C.; Escalona-Buendía, B.H.; Mendoza-Espinoza, A.J.; Villa-Hernández, M.J.; Lobato-Ortíz, R.; Rodríguez-Pérez, E.J.; Pérez-Flores, J.L. Antioxidant Balance and Regulation in Tomato Genotypes of Different Color. J. Am. Soc. Hortic. Sci. 2019, 144, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Asensio, E.; Sanvicente, I.; Mallor, C.; Menal-Puey, S. Spanish traditional tomato. Effects of genotype, location and agronomic conditions on the nutritional quality and evaluation of consumer preferences. Food Chem. 2019, 270, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Valverde, I.; Periago, M.J.; Provan, G.; Chesson, A. Phenolic compounds, lycopene and antioxidant activity in commercial varieties of tomato (Lycopersicum esculentum). J. Sci. Food Agric. 2002, 82, 323–330. [Google Scholar] [CrossRef]
- Karlova, R.; Chapman, N.; David, K.; Angenent, G.C.; Seymour, G.B.; de Maagd, R.A. Transcriptional control of fleshy fruit development and ripening. J. Exp. Bot. 2014, 65, 4527–4541. [Google Scholar] [CrossRef] [Green Version]
- Macheix, J.-J.; Fleuriet, A.; Billot, J. Fruit Phenolics, 1st ed.; CRC Press: Boca Raton, FL, USA, 1990; p. 390. [Google Scholar]
- Kavitha, P.; Shivashankara, K.S.; Rao, V.K.; Sadashiva, A.T.; Ravishankar, K.V.; Sathish, G.J. Genotypic variability for antioxidant and quality parameters among tomato cultivars, hybrids, cherry tomatoes and wild species. J. Sci. Food Agric. 2014, 94, 993–999. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Lee, J.G. Ripening-Dependent Changes in Antioxidants, Color Attributes, and Antioxidant Activity of Seven Tomato (Solanum lycopersicum L.) Cultivars. J. Anal. Methods Chem. 2016, 2016, 5498618. [Google Scholar] [CrossRef] [Green Version]
- Cano, A.; Acosta, M.; Arnao, M.B. Hydrophilic and lipophilic antioxidant activity changes during on-vine ripening of tomatoes (Lycopersicon esculentum Mill.). Postharvest Biol. Technol. 2003, 28, 59–65. [Google Scholar] [CrossRef]
- Ji, L.; Wu, J.; Gao, W.; Wei, J.; Yang, J.; Guo, C. Antioxidant Capacity of Different Fractions of Vegetables and Correlation with the Contents of Ascorbic Acid, Phenolics, and Flavonoids. J. Food Sci. 2011, 76, C1257–C1261. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Creissen, G.; Kular, B.; Firmin, J.; Robinson, S.; Verhoeyen, M.; Mullineaux, P. Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening. Planta 2002, 214, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Bangalore, D.V.; McGlynn, W.; Scott, D.D. Effect of β-Cyclodextrin in Improving the Correlation between Lycopene Concentration and ORAC Values. J. Agric. Food Chem. 2005, 53, 1878–1883. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]

{kind=link}
| Variety | Type | Weight (g) | Shape | Color in Full Ripening |
|---|---|---|---|---|
| Elpida F1 | uniform large round (beef-small beef type) | 200–250 | spherical | bright red |
| Short of Kythira | large beefsteake type | 250–300 | flattened with intense adhesions | light red |
| Long of Kythira | large beefsteake type | 150–200 | cordate elongated | light purple |
| Chiou | cherry | 14–19 | spherical | light red |
| Authentic of Santorini | marmande (coctail type) | 30–40 | flattened | deep red |
| Kaisia or Traditional of Santorini | cherry | 18–20 | light flattened, spherical | deep red |
| Variety | Fat (mg/100 g FW) | Total Carbohydrates (g/100 g FW) | Protein (mg/100 g FW) | Energy (Kcal/100 g FW) | ||||
|---|---|---|---|---|---|---|---|---|
| Red Ripe | Breaker | Red Ripe | Breaker | Red Ripe | Breaker | Red Ripe | Breaker | |
| Elpida F1 | 47.85 ± 3.90 a | 45.66 ± 5.45 a | 1.80 ± 0.09 ab | 1.84 ± 0.08 a | 255.32 ± 43.68 a* | 120.14 ± 12.02 a* | 7.91 ± 0.44 ab | 7.57 ± 0.26 a |
| Short of Kythira | 29.15 ± 5.45 a | 23.65 ± 2.33 a | 1.15 ± 0.06 cd | 1.52 ± 0.08 a | 275.99 ± 44.14 a* | 77.76 ± 2.44 b* | 5.68 ± 0.32 c | 6.22 ± 0.31 b |
| Long of Kythira | 24.20 ± 3.11 a | 38.50 ± 9.34 a | 2.15 ± 0.43 a* | 3.04 ± 0.33 b* | 277.41 ± 59.14 a | 234.90 ± 21.89 c | 9.39 ± 1.47 de* | 12.69 ± 1.23 c* |
| Chiou | 132.56 ± 10.11 b* | 167.22 ± 7.78 b* | 1.01 ± 0.03 d | 1.03 ± 0.03 c | 596.19 ± 4.01 b* | 394.96 ± 30.53 d* | 10.38 ± 0.42d* | 6.95 ± 0.25 ab* |
| Authentic of Santorini | 99.01 ± 7.80 c* | 176.02 ± 7.78 b* | 2.25 ± 0.40 a | 2.56 ± 0.05 b | 229.24 ± 28.36 a | 199.48 ± 52.09 ac | 10.26 ± 1.52d | 11.59 ± 0.35 cd |
| Kaisia or Traditional of Santorini | 103.96 ± 8.66 c* | 155.12 ± 4.67 b* | 1.56 ± 0.02 bc* | 2.37 ± 0.14 b* | 256.16 ± 12.02 a* | 113.36 ± 9.84 ab* | 7.32 ± 0.62be* | 10.62 ± 0.50 d* |
| Variety | Ascorbic acid (mg/100 g FW) | Lycopene (mg/100 g FW) | Total Phenols (mg GAE/100 g FW) | Flavonoids (mg RE/100 g FW) | ||||
|---|---|---|---|---|---|---|---|---|
| Red Ripe | Breaker | Red Ripe | Breaker | Red Ripe | Breaker | Red Ripe | Breaker | |
| Elpida F1 | 32.11 ± 0.75 a | 28.49 ± 2.40 a | 2.24 ± 0.22 a* | 1.20 ± 0.05 a* | 34.41 ± 6.42 ab* | 19.08 ± 0.19 a* | 6.90 ± 0.76 a* | 1.25 ± 0.05 a* |
| Short of Kythira | 29.74 ± 1.47 a* | 36.90 ± 0.38 b* | 6.04 ± 0.35 b* | 1.09 ± 0.09 ab* | 19.01 ± 3.07 c | 14.19 ± 1.09 b | 5.80 ± 0.63 ab* | 2.99 ± 0.59 bc* |
| Long of Kythira | 36.04 ± 0.56 b* | 39.03 ± 0.20 b* | 5.02 ± 0.08 c* | 0.79 ± 0.02 c* | 17.50 ± 1.08 c* | 13.68 ± 0.19 b* | 4.29 ± 0.50 b* | 1.79 ± 0.12 ab* |
| Chiou | 41.66 ± 3.35 bc | 40.65 ± 2.11 b | 3.41 ± 0.13 d* | 0.50 ± 0.02 d* | 45.26 ± 7.31 ad | 39.56 ± 4.54 c | 6.96 ± 0.76 a* | 4.11 ± 0.25 cd* |
| Authentic of Santorini | 54.41 ± 1.04 d* | 61.63 ± 0.61 c* | 4.34 ± 0.02 e* | 1.02 ± 0.03 b* | 27.89 ± 1.97 b* | 21.05 ± 2.31 a* | 5.06 ± 0.21 b | 4.46 ± 0.25 d |
| Kaisia or Traditional of Santorini | 43.82 ± 4.24 c | 53.06 ± 2.16 d | 7.98 ± 0.03 f* | 3.69 ± 0.09 e* | 47.89 ± 5.74 d* | 23.42 ± 1.49 a* | 6.25 ± 0.54 a | 6.67 ± 1.97 d |
| Variety | DPPH (mg TE/100 g FW) | FRAP (mg TE/100 g FW) | ||
|---|---|---|---|---|
| Red Ripe | Breaker | Red Ripe | Breaker | |
| Elpida F1 | 27.54 ± 1.59 a | 25.11 ± 2.07 a | 25.41 ± 2.51 a | 23.12 ± 0.53 a |
| Short of Kythira | 24.92 ± 2.09 a | 21.43 ± 1.05 b | 27.86 ± 0.85 a* | 15.03 ± 1.36 b* |
| Long of Kythira | 19.14 ± 0.9 b | 15.56 ± 3.10 cb | 26.33 ± 0.88 a | 23.63 ± 0.56 a |
| Chiou | 33.36 ± 0.71 c | 32.74 ± 0.16 d | 46.38 ± 1.48 b* | 42.77 ± 0.3 c* |
| Authentic of Santorini | 19.48 ± 1.44 b | 17.86 ± 2.58 cb | 39.64 ± 1.59 c* | 26.54 ± 0.65 a* |
| Kaisia or Traditional of Santorini | 48.34 ± 1.35 d | 45.65 ± 11.41 e | 56.09 ± 4.94 d | 47.81 ± 10.91 c |
| Variety | DPPH (mg TE/100 g FW) | FRAP (mg TE/100 g FW) | ||
|---|---|---|---|---|
| Red Ripe | Breaker | Red Ripe | Breaker | |
| Elpida F1 | 1.55 ± 0.04 a* | 0.76 ± 0.18 ab* | 2.45 ± 0.3 a | 1.82 ± 0.68 abc |
| Short of Kythira | 2.32 ± 0.28 bc* | 0.69 ± 0.05 a* | 5.11 ± 0.31 b* | 0.94 ± 0.05 a* |
| Long of Kythira | 2.16 ± 0.18 bc* | 0.33 ± 0.02 c* | 4.81 ± 0.03 b* | 0.89 ± 0.03 a* |
| Chiou | 1.80 ± 0.12 ab* | 0.26 ± 0.08 c* | 3.76 ± 0.21 c* | 1.61 ± 0.18 b* |
| Authentic of Santorini | 2.25 ± 0.15 c* | 0.89 ± 0.08 ab* | 4.60 ± 0.21 b* | 1.74 ± 0.20 b* |
| Kaisia or Traditional of Santorini | 3.25 ± 0.28 d* | 1.71 ± 0.16 c* | 5.21 ± 0.67 b* | 2.82 ± 0.77 c* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgaki, E.; Nifakos, K.; Kotsiras, A.; Fanourakis, D.; Tsaniklidis, G.; Delis, C.; Spiliopoulos, I.K. Comparison of Nutrient Composition and Antioxidant Activity of Hydroponically Grown Commercial and Traditional Greek Tomato Cultivars. Horticulturae 2023, 9, 163. https://doi.org/10.3390/horticulturae9020163
Georgaki E, Nifakos K, Kotsiras A, Fanourakis D, Tsaniklidis G, Delis C, Spiliopoulos IK. Comparison of Nutrient Composition and Antioxidant Activity of Hydroponically Grown Commercial and Traditional Greek Tomato Cultivars. Horticulturae. 2023; 9(2):163. https://doi.org/10.3390/horticulturae9020163
Chicago/Turabian StyleGeorgaki, Evangelia, Kallimachos Nifakos, Anastasios Kotsiras, Dimitrios Fanourakis, George Tsaniklidis, Costas Delis, and Ioakim K. Spiliopoulos. 2023. "Comparison of Nutrient Composition and Antioxidant Activity of Hydroponically Grown Commercial and Traditional Greek Tomato Cultivars" Horticulturae 9, no. 2: 163. https://doi.org/10.3390/horticulturae9020163
APA StyleGeorgaki, E., Nifakos, K., Kotsiras, A., Fanourakis, D., Tsaniklidis, G., Delis, C., & Spiliopoulos, I. K. (2023). Comparison of Nutrient Composition and Antioxidant Activity of Hydroponically Grown Commercial and Traditional Greek Tomato Cultivars. Horticulturae, 9(2), 163. https://doi.org/10.3390/horticulturae9020163