
Genome-Wide Identification of the Sweet Orange bZIP Gene Family and Analysis of Their Expression in Response to Infection by Penicillium digitatum
 , and
, and 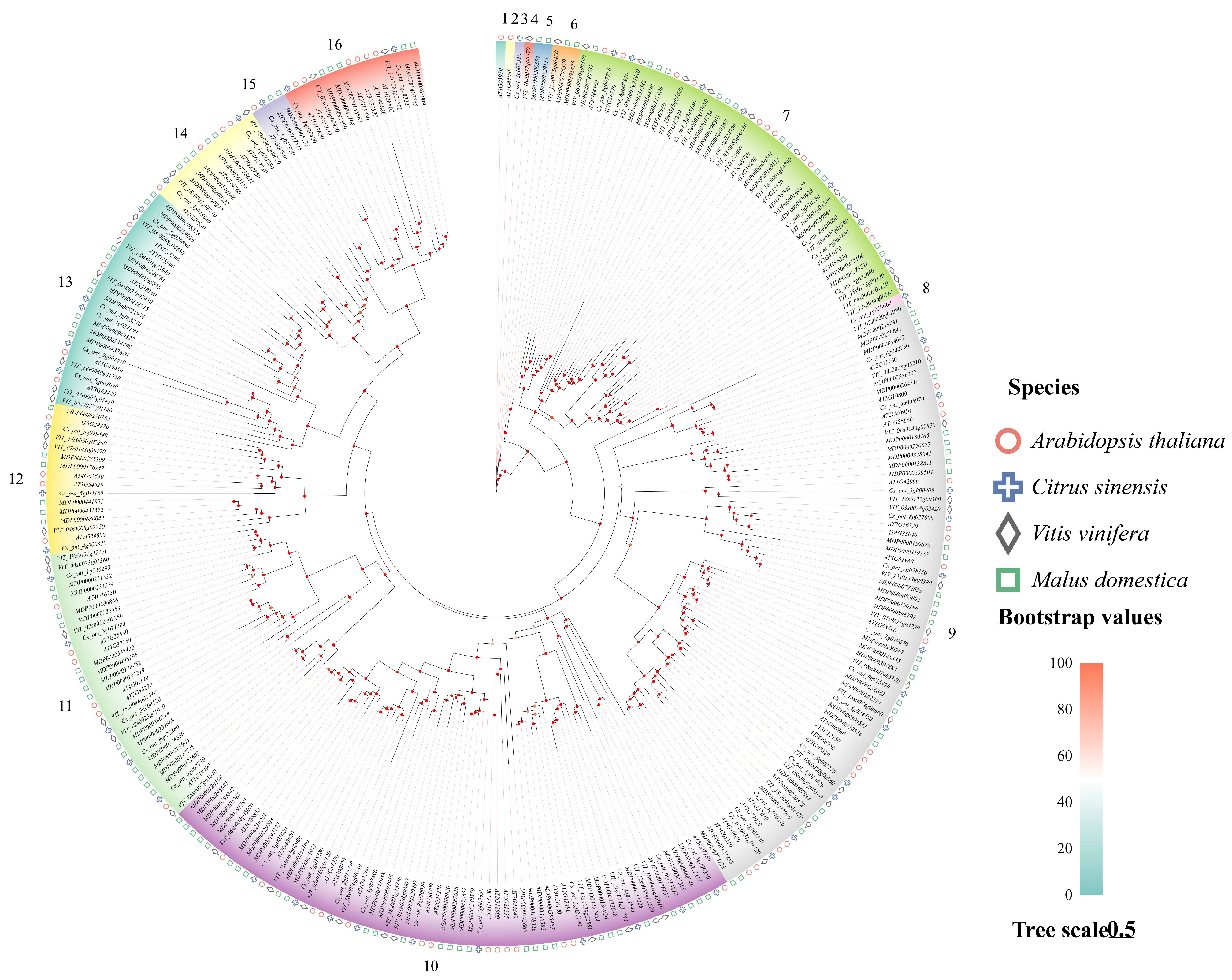{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of the bZIP Gene Family in Sweet Orange
2.2. Physicochemical Properties, Predicted Secondary Structure, and Subcellular Localization of the Sweet Orange bZIP Gene
2.3. Phylogenetic Analysis of the Sweet Orange bZIP Gene Family
2.4. Analysis of the Conserved Motifs and Gene Structure of the Sweet Orange bZIP Gene Family
2.5. Analysis of the Sweet Orange bZIP Gene Family’s Cis-Acting Elements
2.6. Analysis of the Gene Localization and Covariance of the Sweet Orange bZIP Gene Family
2.7. Analysis of the Tissue Expression of the Sweet Orange bZIP Gene Family
3. Results and Analysis
3.1. Identification and Characterization of the bZIP Transcription Factor Families of Sweet Orange
3.2. Phylogenetic Analysis of the Sweet Orange bZIP Gene Family
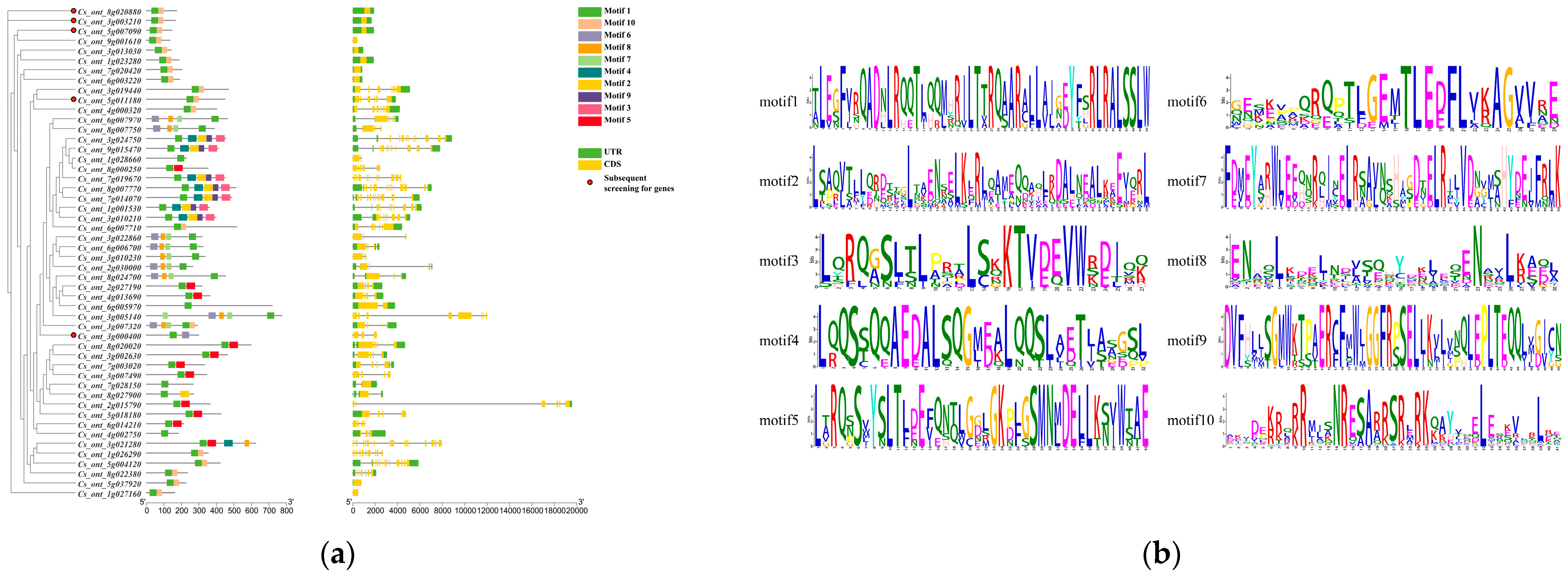
3.3. Gene Structure Analysis of the Sweet Orange bZIP Gene Family
3.4. Analysis of the Promoter Cis-Acting Elements of the Sweet Orange bZIP Gene Family
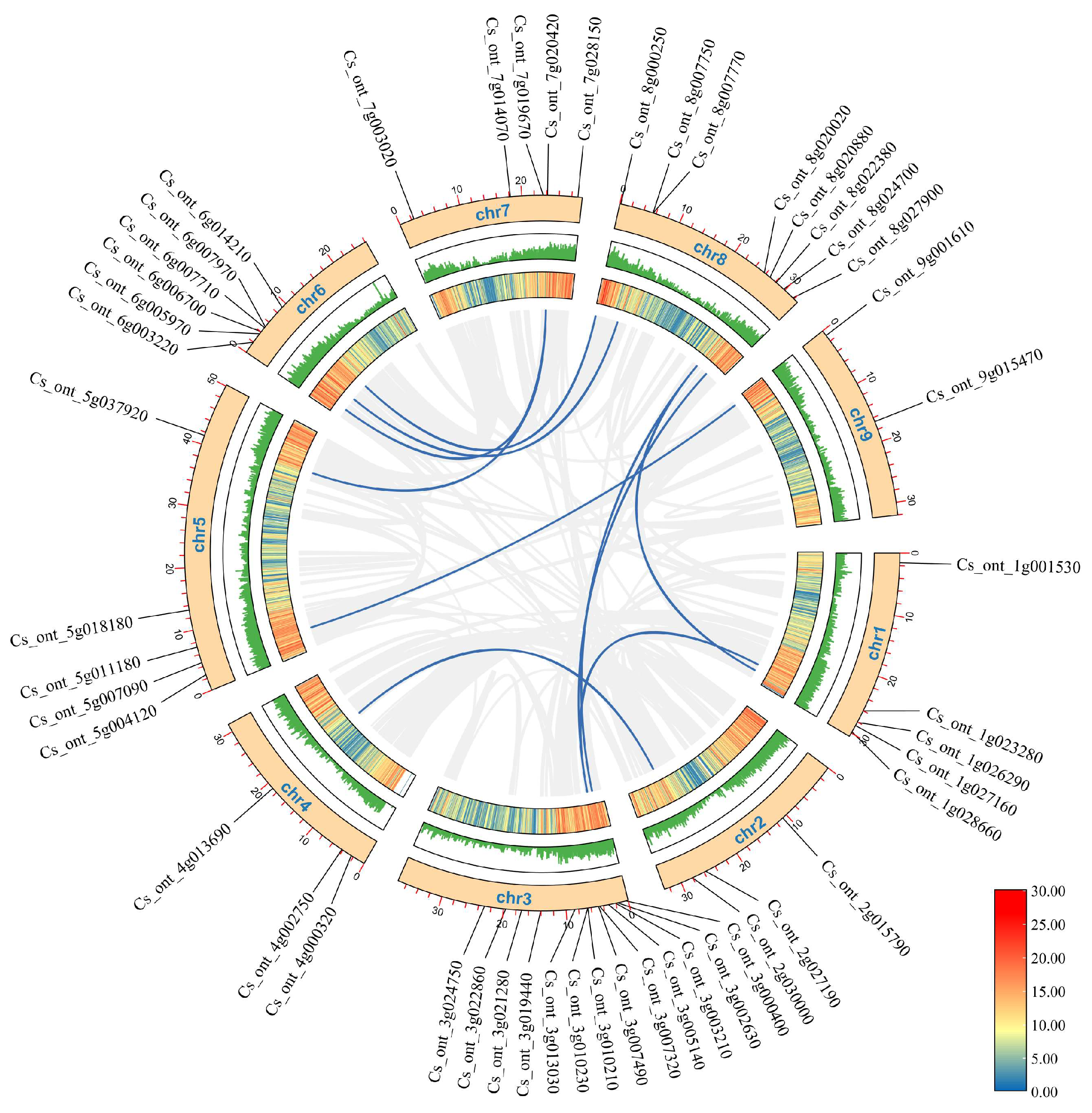
3.5. Analysis of Gene Covariation and Chromosomal Localization in the Sweet Orange bZIP Gene Family
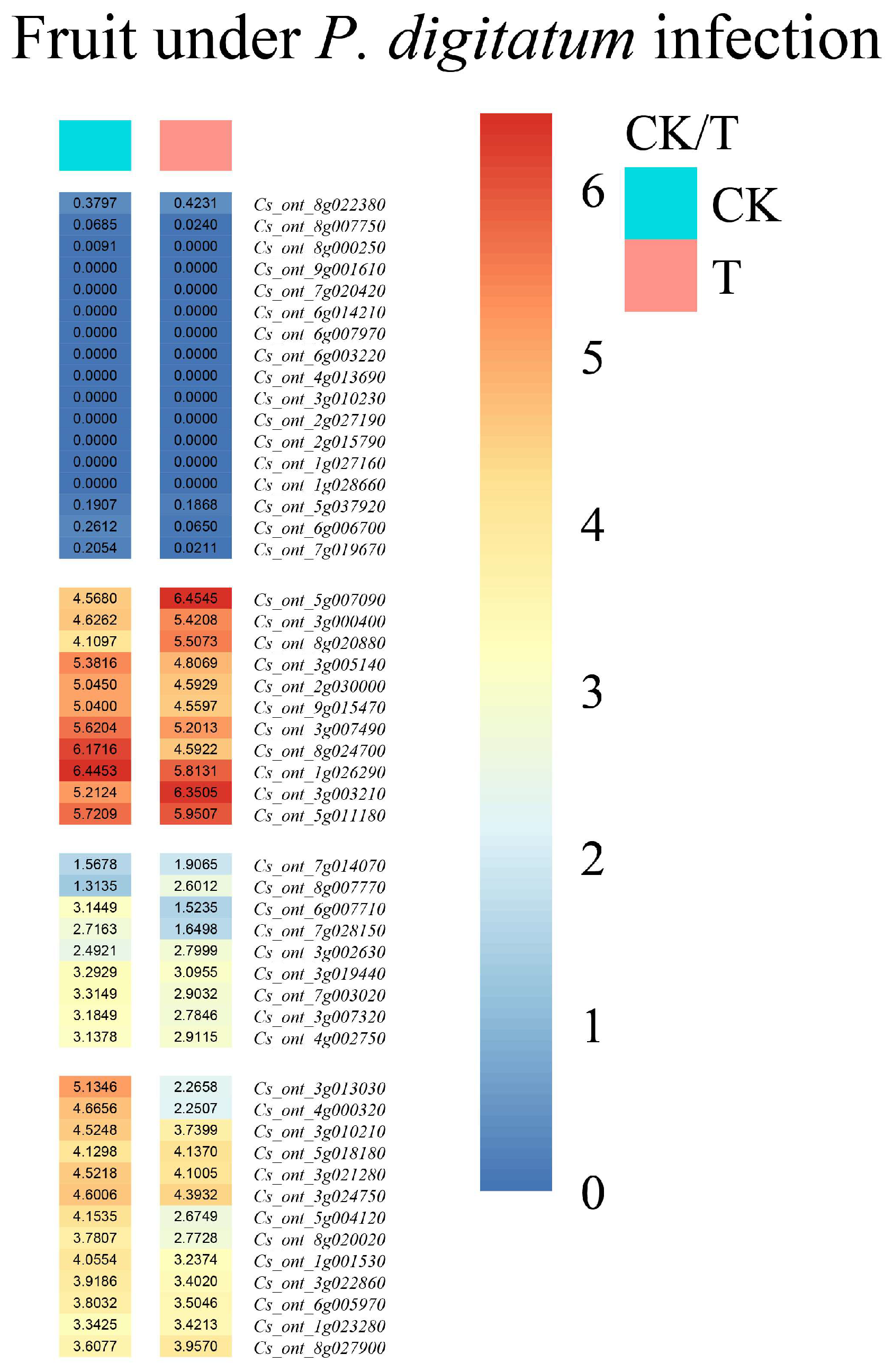
3.6. Analysis of the Tissue Expression of Sweet Orange bZIP Gene Family
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, X.; Zhou, Y.; Wang, B.; Ding, L.; Wang, Y.; Luo, L.; Zhang, Y.; Kong, W. Genome-wide identification and characterization of laccase gene family in Citrus sinensis. Gene 2019, 689, 114–123. [Google Scholar] [CrossRef]
- Chen, Y. Study on international competitiveness of China’s orange export. Guangdong Agric. Sci. 2016, 43, 176–182. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, C.; Lai, J.; Peng, Y.; Zhong, C.; Liang, M. Infection characteristics and interaction of three species of Penicillium in the postharvest storage of ’Gonggan’ mandarin (Citrus reticulata). Agric. Biotechnol. 2022, 11, 148–153. [Google Scholar]
- Moraes Bazioli, J.; Belinato, J.R.; Costa, J.H.; Akiyama, D.Y.; Pontes, J.G.M.; Kupper, K.C.; Augusto, F.; de Carvalho, J.E.; Fill, T.P. Biological Control of Citrus Postharvest Phytopathogens. Toxins 2019, 11, 460. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Stewart, C.N., Jr. Plant synthetic promoters and transcription factors. Curr. Opin. Biotechnol. 2016, 37, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Chu, Z. Genome-wide evolutionary characterization and analysis of bZIP transcription factors and their expression profiles in response to multiple abiotic stresses in Brachypodium distachyon. BMC Genom. 2015, 16, 227. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ye, S.; Du, Y.; Zhao, Q.; Du, J.; Zhang, Q. Identification and expression analysis of bZIP members under abiotic stress in Mung Bean (Vigna radiata). Life 2022, 12, 938. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription Factors Associated with Abiotic and Biotic Stress Tolerance and Their Potential for Crops Improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noman, A.; Liu, Z.; Aqeel, M.; Zainab, M.; Khan, M.I.; Hussain, A.; Ashraf, M.F.; Li, X.; Weng, Y.; He, S. Basic leucine zipper domain transcription factors: The vanguards in plant immunity. Biotechnol. Lett. 2017, 39, 1779–1791. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Yan, S. Bioinformatics and Expression Analysis of the Basic Leucine Zipper Gene Family in Plum. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2016. [Google Scholar]
- Liu, Y.; Wang, L.; Li, X.; Xiao, H.; Wan, S.; Zhang, Y. Bioinformatics and functional analysis of five members of the HbbZIP gene family in Hevea brasiliensis. Mol. Plant Breed. 2019, 17, 5613–5623. [Google Scholar] [CrossRef]
- Cao, P. Screening of Poplar bZIP Gene Family and Functional Characterization of the bZIP53 Transcription Factor. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2018. [Google Scholar] [CrossRef]
- Liu, J.; Chen, N.; Chen, F.; Cai, B.; Santo, S.D.; Tornielli, G.B.; Pezzotti, M.; Cheng, Z.-M.M. Genome-wide analysis and expression profile of the bZIP transcription factor gene family in grapevine (Vitis vinifera). BMC Genom. 2014, 15, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F. Comparative genomics analysis of bZIP family members in Actinidia chinensis. J. Appl. Environ. Biol. 2020, 26, 10. [Google Scholar] [CrossRef]
- Guo, Y.; Shi, J.; Zhou, M.; Yu, Y.; Wang, C. Analysis of the BpbZIP1 gene of Betula alba for drought and salt tolerance and identification of ABRE element binding. For. Sci. Res. 2020, 33, 9. [Google Scholar] [CrossRef]
- Jia, L.; Zhao, Q.; Chen, S. The expression profiling of 11 ZmbZIP genes under abiotic stress in maize. J. Anhui Agric. Univ. 2022, 49, 197–204. [Google Scholar] [CrossRef]
- Zhuang, W.; Yang, L.; Shao, L.; Yang, D.; Yang, J. Research progress on stress resistance of bZIP transcription factors in forest trees. Mol. Plant Breed. 2022, 12, 1–8. [Google Scholar]
- Tak, H.; Mhatre, M. Cloning and molecular characterization of a putative bZIP transcription factor VvbZIP23 from Vitis vinifera. Protoplasma 2013, 250, 333–345. [Google Scholar] [CrossRef]
- Li, Q.; Jia, R.; Dou, W.; Qi, J.; Qin, X.; Fu, Y.; He, Y.; Chen, S. CsBZIP40, a BZIP transcription factor in sweet orange, plays a positive regulatory role in citrus bacterial canker response and tolerance. PLoS ONE 2019, 14, e0223498. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Chen, S.; Yao, W.; Cheng, Z.; Zhou, B.; Jiang, T. Genome-wide analysis and expression profile of the bZIP gene family in poplar. BMC Plant Biol. 2021, 21, 122. [Google Scholar] [CrossRef]
- de Paula Santos Martins, C.; Pedrosa, A.M.; Du, D.; Goncalves, L.P.; Yu, Q.; Gmitter, F.G., Jr.; Costa, M.G.C. Genome-wide characterization and expression analysis of major intrinsic proteins during abiotic and biotic stresses in sweet orange (Citrus sinensis L. Osb.). PLoS ONE 2015, 10, e0138786. [Google Scholar] [CrossRef]
- Liu, H.; Wang, X.; Liu, S.; Huang, Y.; Guo, Y.-X.; Xie, W.-Z.; Liu, H.; Qamar, M.T.U.; Xu, Q.; Chen, L.-L. Citrus Pan-Genome to Breeding Database (CPBD): A comprehensive genome database for citrus breeding. Mol. Plant 2022, 15, 1503–1505. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Hernandez, M.; Berardini, T.; Chen, G.; Crist, D.; Doyle, A.; Huala, E.; Knee, E.; Lambrecht, M.; Miller, N.; Mueller, L.A.; et al. TAIR: A resource for integrated Arabidopsis data. Funct. Integr. Genom. 2002, 2, 239–253. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; López, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Polido, P.; Rampim, M.; Kaschuk, G.; de Souza, S.G.H. Genome-wide identification and phylogenetic analysis of the AP2/ERF gene superfamily in sweet orange (Citrus sinensis). Genet. Mol. Res. 2014, 13, 7839–7851. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, R.; Zhang, Z.; Zhao, T.; Zhang, D.; Sofkova, S.; Wu, Y.; Wang, Y. Genome-wide analysis of the bZIP gene lineage in apple and functional analysis of MhABF in Malus halliana. Planta 2021, 254, 78. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Yu, W.; Kong, G.; Chao, J.; Yin, T.; Tian, H.; Ya, H.; He, L.; Zhang, H. Genome-wide identification of the rubber tree superoxide dismutase (SOD) gene family and analysis of its expression under abiotic stress. PeerJ 2022, 10, e14251. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Srivastava, R.; Che, P.; Howell, S.H. Salt stress responses in Arabidopsis utilize a signal transduction pathway related to endoplasmic reticulum stress signaling. Plant J. 2007, 51, 897–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Lilay, G.H.; Castro, P.H.; Guedes, J.G.; Almeida, D.M.; Campilho, A.; Azevedo, H.; Aarts, M.G.; Saibo, N.J.; Assunção, A.G. Rice F-bZIP transcription factors regulate the zinc deficiency response. J. Exp. Bot. 2020, 71, 3664–3677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, T.; Han, P.; Xi, D.; Yu, W.; Zhu, L.; Du, C.; Yang, N.; Liu, X.; Zhang, H. Genome-wide identification, characterization, and expression profile of NBS-LRR gene family in sweet orange (Citrus sinensis). Gene 2023, 854, 147117. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, Z.; Zhao, W.; Yin, T.; Liu, X.; Zhang, H. Overexpression of MET4 leads to the upregulation of stress-related genes and enhanced sulfite tolerance in Saccharomyces Uvarum. Cells 2022, 11, 636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, C.; Huang, X.; Zhang, H.; Yuan, Z. Land-plant phylogenomic and pomegranate transcriptomic analyses reveal an evolutionary scenario of CYP75 genes subsequent to whole genome duplications. J. Plant Biol. 2019, 62, 48–60. [Google Scholar] [CrossRef]
- Xi, D.; Yin, T.; Han, P.; Yang, X.; Zhang, M.; Du, C.; Zhang, H.; Liu, X. Genome-wide identifification of sweet orange WRKY transcription factors and analysis of their expression in response to infection by Penicillium digitatum. Curr. Issues Mol. Biol. 2023, 45, 1250–1271. [Google Scholar] [CrossRef]
- Li, M.; Tang, Y.; Yu, M.; Fan, Y.; Khan, S.U.; Chang, W.; Li, X.; Wei, S.; Wei, L.; Qu, C.; et al. Systematic characterization of Brassica napus HIR gene family reveals a positive role of BnHIR2.7 in Sclerotinia sclerotiorum resistance. Horticulturae 2022, 8, 874. [Google Scholar] [CrossRef]
- Li, S.; Huang, H.; Ma, X.; Hu, Z.; Li, J.; Yin, H. Characterizations of MYB transcription factors in Camellia oleifera reveal the key regulators involved in Oil Biosynthesis. Horticulturae 2022, 8, 742. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, L.; Xia, Q. Research progress on the regulation of plant hormones to stress responses. J. Hubei Univ. Nat. Sci. 2021, 43, 13. [Google Scholar] [CrossRef]
- Sun, Y.; Cai, T.; Ji, X.; Zhang, J. Genome-wide bioinformatics analysis of bZIP gene family in Pyrus communis. For. Ecol. Sci. 2021, 36, 24–34. [Google Scholar] [CrossRef]
- Xing, Y. Genome-Wide Identification of the Basic Leucine Zipper (bZIP) Gene Family in Cotton. Master’s Thesis, Shandong Agricultural University, Taian, China, 2020. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Lynch, T.J. The Arabidopsis abscisic acid response gene ABI5 encodes a basic leucine zipper transcription factor. Plant Cell 2000, 12, 599–609. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Choi, H.W.; Hwang, I.S.; Choi, D.S.; Hwang, B.K. Functional roles of the pepper pathogen-induced bZIP transcription factor, CAbZIP1, in enhanced resistance to pathogen infection and environmental stresses. Planta 2006, 224, 1209–1225. [Google Scholar] [CrossRef]
- Lei, G.; Li, Z.; Wang, X.; Li, Z. Research progress on control of green mold of postharvest citrus fruits. Jiangsu Agric. Sci. 2018, 46, 27–30. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, F.; Xiao, L.; Peng, W.; Chen, M.; Chen, J.; Xiang, M. Effects of methyl jasmonate treatment on blue mold and defense enzymes activity of navel orange fruit. Acta Agric. Univ. Jiangxiensis 2022, 44, 560–568. [Google Scholar] [CrossRef]
- Bian, W. Effects of Microencapsulated Salicylic Acid Induced Resistance to Blue Mold in Postharvest Apple. Master’s Thesis, Shanghai University of Technology, Shanghai, China, 2019. [Google Scholar] [CrossRef]
- Alves, M.S.; Dadalto, S.P.; Gonçalves, A.B.; De Souza, G.B.; Barros, V.A.; Fietto, L.G. Plant bZIP transcription factors responsive to pathogens: A review. Int. J. Mol. Sci. 2013, 14, 7815–7828. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Yu, C.; Luo, L.; Pan, H.; Zhang, Q. Identification and expression analysis of bZIP transcription factor family in Rosa persica. J. Northwest A F Univ. (Nat. Sci. Ed.) 2022, 50, 82–92. [Google Scholar] [CrossRef]
- Sun, M.-Y.; Fu, X.-L.; Tan, Q.-P.; Liu, L.; Chen, M.; Zhu, C.-Y.; Li, L.; Chen, X.-D.; Gao, D.-S. Analysis of basic leucine zipper genes and their expression during bud dormancy in peach (Prunus persica). Plant Physiol. Biochem. 2016, 104, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.M.; Deng, L.; Meng, J.; Wang, Y.; Pan, L.; Niu, L.; Lu, Z.; Cui, G.; Zeng, W.; Wang, Z. Characterization and expression analysis of basic leucine zipper (bZIP) transcription factors responsive to chilling injury in peach fruit. Mol. Biol. Rep. 2022, 50, 361–376. [Google Scholar] [CrossRef]
- Liang, J.; Zhao, Q. Identification and analysis of bZlP transcription factor family regulating oil metabolism in Vernicia fordi. Plant Physiol. J. 2021, 57, 1135–1150. [Google Scholar] [CrossRef]
- Darwish, O.S.; Ali, M.R.; Khojah, E.; Samra, B.N.; Ramadan, K.M.A.; El-Mogy, M.M. Pre-harvest application of salicylic acid, abscisic acid, and methyl jasmonate conserve bioactive compounds of strawberry fruits during refrigerated storage. Horticulturae 2021, 7, 568. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, P.; Yin, T.; Xi, D.; Yang, X.; Zhang, M.; Zhu, L.; Zhang, H.; Liu, X. Genome-Wide Identification of the Sweet Orange bZIP Gene Family and Analysis of Their Expression in Response to Infection by Penicillium digitatum. Horticulturae 2023, 9, 393. https://doi.org/10.3390/horticulturae9030393
Han P, Yin T, Xi D, Yang X, Zhang M, Zhu L, Zhang H, Liu X. Genome-Wide Identification of the Sweet Orange bZIP Gene Family and Analysis of Their Expression in Response to Infection by Penicillium digitatum. Horticulturae. 2023; 9(3):393. https://doi.org/10.3390/horticulturae9030393
Chicago/Turabian StyleHan, Peichen, Tuo Yin, Dengxian Xi, Xiuyao Yang, Mengjie Zhang, Ling Zhu, Hanyao Zhang, and Xiaozhen Liu. 2023. "Genome-Wide Identification of the Sweet Orange bZIP Gene Family and Analysis of Their Expression in Response to Infection by Penicillium digitatum" Horticulturae 9, no. 3: 393. https://doi.org/10.3390/horticulturae9030393
APA StyleHan, P., Yin, T., Xi, D., Yang, X., Zhang, M., Zhu, L., Zhang, H., & Liu, X. (2023). Genome-Wide Identification of the Sweet Orange bZIP Gene Family and Analysis of Their Expression in Response to Infection by Penicillium digitatum. Horticulturae, 9(3), 393. https://doi.org/10.3390/horticulturae9030393