
Multivariate Assessment of Genetic Relationships between Two Streptocarpus Cultivars and Their F1 Progenies Using Morphological Characteristics and SCoT Molecular Markers

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Artificial Hybridization Methodology
2.3. Morphological Characterization of Parents and F1 Plants
2.4. SCoT Molecular Analysis
2.5. Data Processing and Analysis
3. Results
3.1. Reciprocal Artificial Hybridization
3.2. Morphological Characterization of Parents and F1 Progeny from Reciprocal Crosses
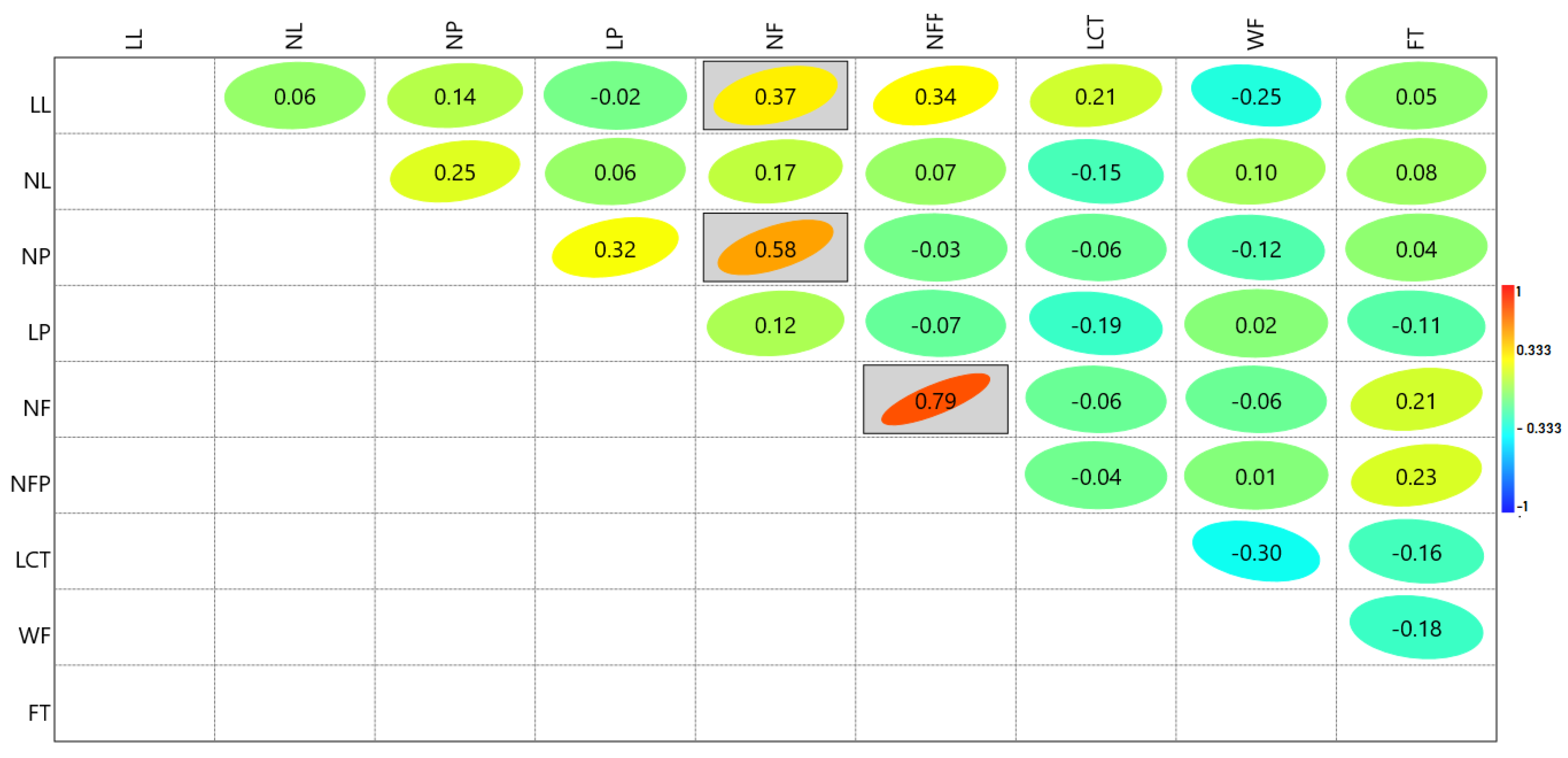
3.3. Correlations between Morphological Parameters of F1 Progeny from Reciprocal Crosses
3.4. Assessment of Genetic Relationships between Parents and F1 Plants
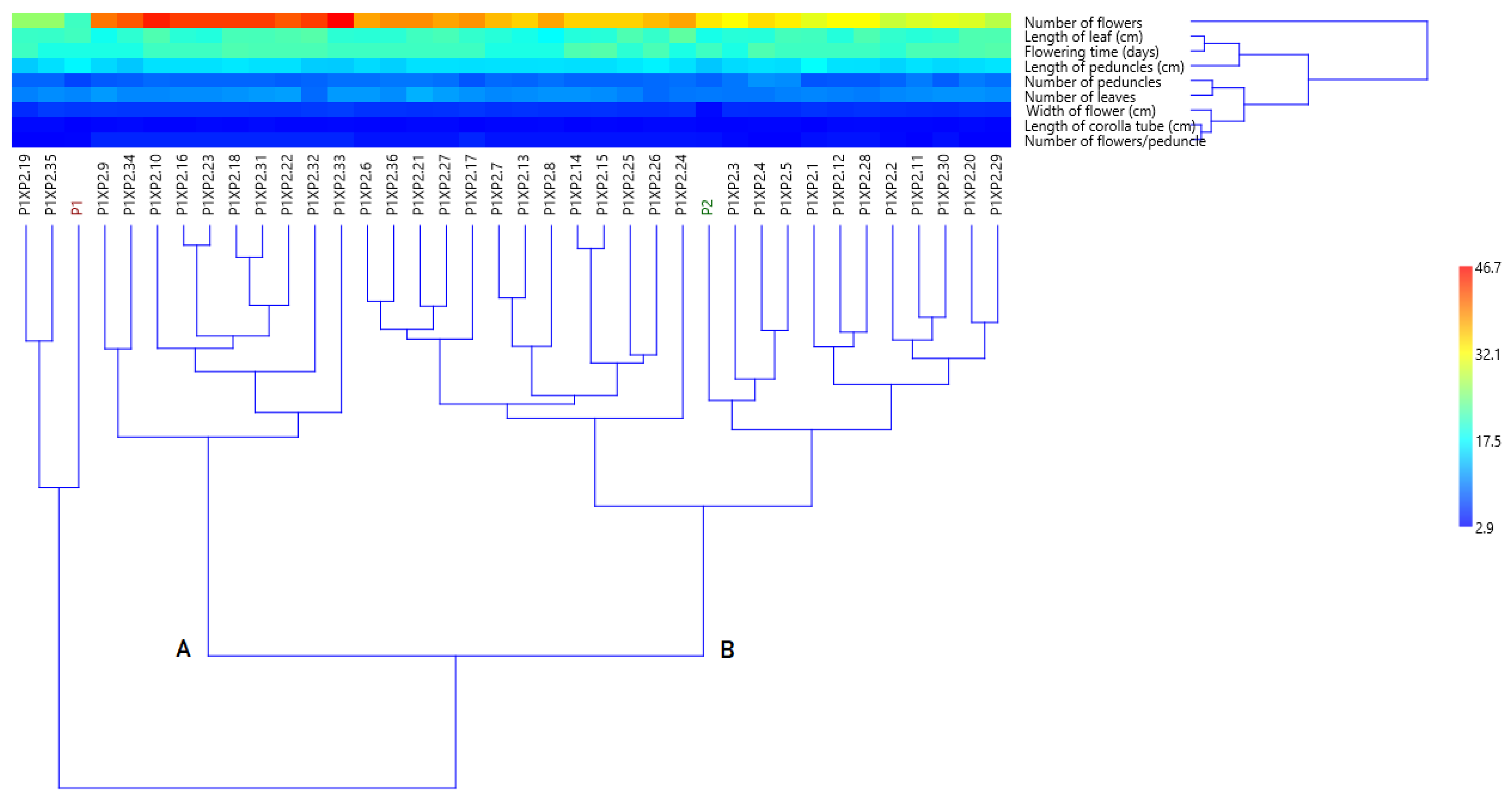
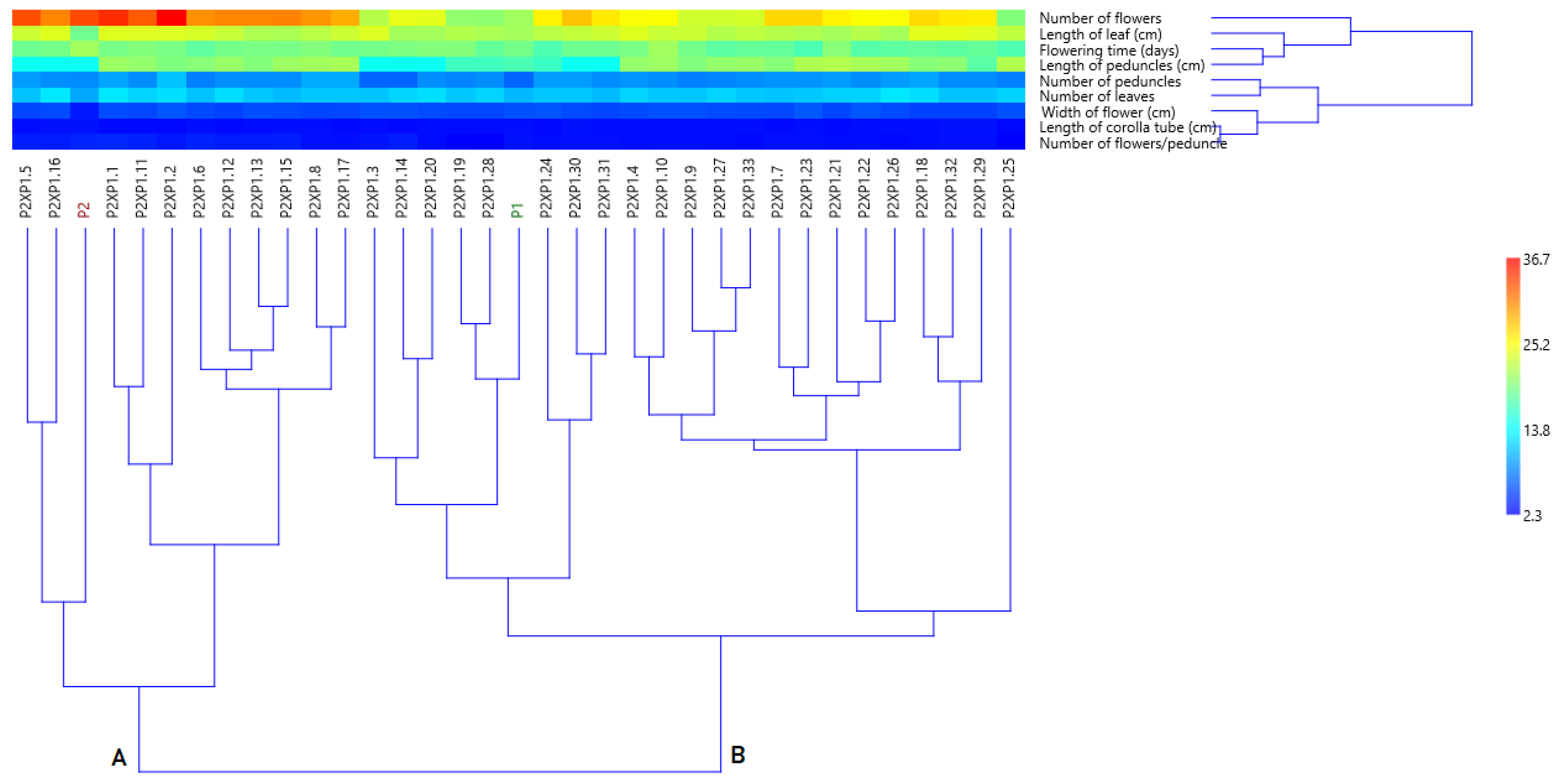
3.4.1. Cluster Analysis Based on Morphological Data
3.4.2. SCoT Markers Polymorphism
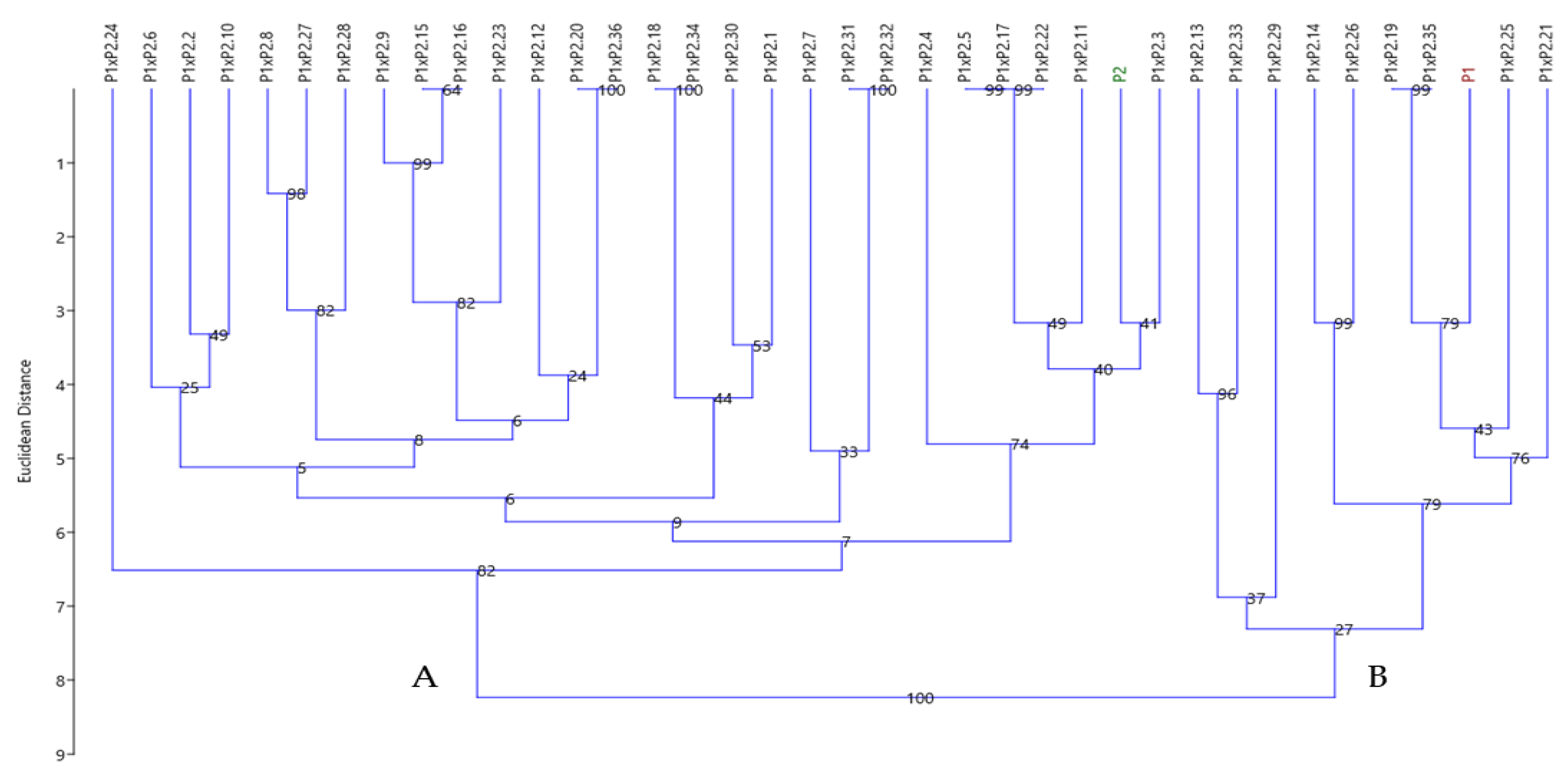
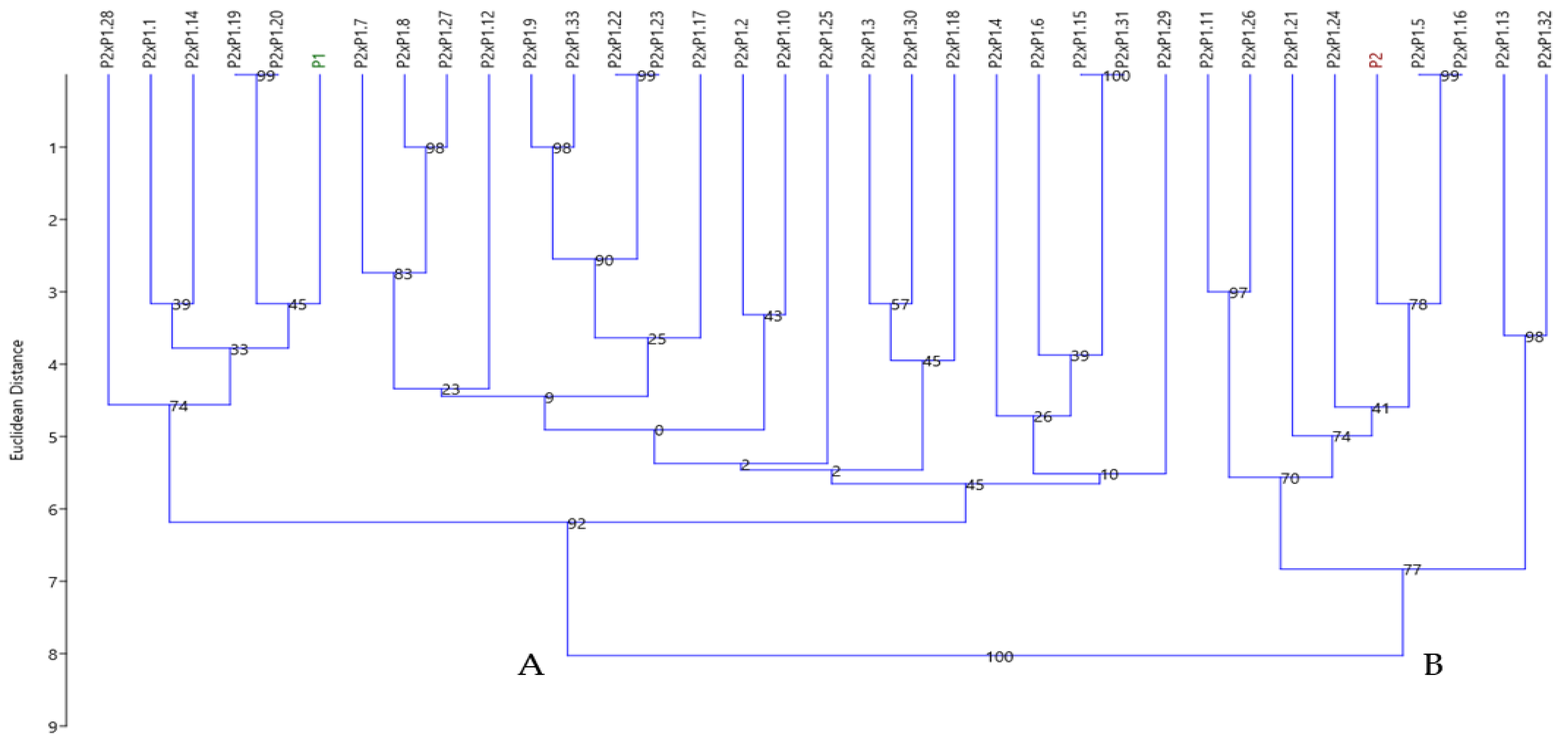
3.4.3. Cluster Analysis Based on SCoT Polymorphism
4. Discussion
4.1. Genetic Diversity of Streptocarpus Parents and Their F1 Progeny Based on Morphological Data
4.2. Genetic Diversity of Streptocarpus Parents and Their F1 Progeny Based on Molecular Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aida, R.; Ohmiya, A.; Onozaki, T. Current researches in ornamental plant breeding. Breed. Sci. 2018, 68, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huylenbroeck, J.V.; Bhattarai, K. Ornamental plant breeding: Entering a new era? Ornam. Hortic. 2022, 28, 297–305. [Google Scholar] [CrossRef]
- Cantor, M.; Stana, D.; Ioana, P.O.P. Streptocarpus-flowering pot plant-propagation and culture. Not. Bot. Horti Agrobot. Cluj-Napoca 2004, 32, 15. [Google Scholar] [CrossRef]
- Wolff, D.W.; Veilleux, R.E. Evaluation of inbred and hybrid populations of Streptocarpus × hybridus. J. Am. Soc. Hortic. Sci. 1986, 111, 622–626. [Google Scholar] [CrossRef]
- Chaudhury, A.; Power, J.B.; Davey, M.R. High frequency direct plant regeneration from leaf and petals of Cape Primrose (Streptocarpus). J. Crop. Sci. Biotechnol. 2010, 13, 107112. [Google Scholar] [CrossRef]
- Tymoszuk, A.; Miler, N. Silver and gold nanoparticles impact on in vitro adventitious organogenesis in chrysanthemum, gerbera and Cape Primrose. Sci. Hortic. 2019, 257, 108766. [Google Scholar] [CrossRef]
- Lawrence, W.J.C.; Scott-Moncrieff, R.; Sturgess, V.C. Studies on Streptocarpus: I. Genetics and chemistry of flower colour in the Garden strains. J. Genet. 1939, 38, 299–306. [Google Scholar] [CrossRef]
- Dibley, R. Streptocarpus varieties. In Streptocarpus, 3rd ed.; Nurseries, D., Ed.; Dibley’s: Wales, UK, 2018; pp. 24–40. [Google Scholar]
- Afkhami-Sarvestani, R.; Serek, M.; Winkelmann, T. Interspecific crosses within the Streptocarpus subgenus Streptocarpella and intergeneric crosses between Streptocarpella and Saintpaulia ionantha genotypes. Sci. Hortic. 2012, 148, 215–222. [Google Scholar] [CrossRef]
- Hârţa, M.; Borsai, O.; Muntean, C.M.; Dina, N.E.; Fǎlǎmaş, A.; Olar, L.E.; Szabo, K.; Pamfil, D.; Ştefan, R. Assessment of Genetic Relationships between Streptocarpus × hybridus V. Parents and F1 Progenies Using SRAP Markers and FT-IR Spectroscopy. Plants 2020, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Davies, F. The British Streptocarpus Society. In Booklet Gesneriads; Shalit, P., Ed.; The Gesneriad Society, Inc.: Seattle, WA, USA, 2016; Volume 66. [Google Scholar]
- Marston, M.E. The morphology of a Streptocarpus hybrid and its regeneration from leaf cuttings. Sci. Hortic. 1964, 17, 114–120. [Google Scholar]
- Gai, J.; He, J. Reciprocal Cross. In Brenner's Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 66–67. [Google Scholar]
- Yates, F. Analysis of data from all possible reciprocal crosses between a set of parental lines. Heredity 1947, 1, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Dibley, R. Streptocarpus varieties. In Streptocarpus, 2nd ed.; Nurseries, D., Ed.; Dibley’s: Wales, UK, 2008; pp. 12–20. [Google Scholar]
- Harta, M.; Pamfil, D.; Borsai, O.; Rodica, P.O.P.; Clapa, D.; Diaconeasa, Z.; Sisea, C.R. Molecular and phytochemical characterization of F1 Streptocarpus hybrids and antioxidant potential of their flower extracts. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 1341–1356. [Google Scholar] [CrossRef]
- Xu, Y.-X.; Shen, S.-Y.; Chen, W.; Chen, L. Analysis of genetic diversity and development of a SCAR marker for green tea (Camellia sinensis) cultivars in Zhejiang Province: The most famous green tea-producing area in China. Biochem. Genet. 2019, 57, 555–570. [Google Scholar] [CrossRef]
- Kobeissi, B.; Saidi, A.; Kobeissi, A.; Shafie, M. Applicability of SCoT and SSR molecular markers for genetic diversity analysis in Chrysanthemum morifolium genotypes. Proc. Natl. Acad. Sci. India B Biol. Sci. 2019, 89, 1067–1077. [Google Scholar] [CrossRef]
- Rayan, W.A.; Osman, S.A. Phylogenetic relationships of some Egyptian soybean cultivars (Glycine max L.) using SCoT marker and protein pattern. Bull. Natl. Res. Cent. 2019, 43, 161. [Google Scholar] [CrossRef] [Green Version]
- Mostafavi, A.S.; Omidi, M.; Azizinezhad, R.; Etminan, A.; Badi, H.N. Genetic diversity analysis in a mini core collection of Damask rose (Rosa damascena Mill.) germplasm from Iran using URP and SCoT markers. J. Genet. Eng. Biotechnol. 2021, 19, 144. [Google Scholar] [CrossRef] [PubMed]
- Lázaro, A.; Totland, Ø. The influence of floral symmetry, dependence on pollinators and pollination generalization on flower size variation. Ann. Bot. 2014, 114, 157–165. [Google Scholar] [CrossRef] [Green Version]
- RHS & Flower Council of Holland. RHS Colour Chart, 3rd ed.; RHS & Flower Council of Holland: Leiden, The Netherlands, 1995. [Google Scholar]
- The Azalea Society of America, Royal Horticultural Society. UCL and RGB Colors, Gamma = 1.4. 2021. Available online: https://www.azaleas.org/color-systems/ (accessed on 19 April 2022).
- Color Mixer-The Best Free Online Color Mixing Tool. Available online: https://artincontext.org/color-mixer/ (accessed on 19 April 2022).
- Lodhi, M.A.; Ye, G.-N.; Weeden, N.F.; Reisch, B.I. A simple and efficient method for DNA extraction from grapevine cultivars and Vitis species. Plant Mol. Biol. Rep. 1994, 12, 6–13. [Google Scholar] [CrossRef]
- Pop, R.; Ardelean, M.; Pamfil, D.; Gaboreanu, I.M. The efficiency of different DNA isolation and purification in ten cultivars of Vitis vinifera. Bull. UASVM Anim. Sci. Biotechnol 2003, 59, 259–261. [Google Scholar]
- Bodea, M.; Pamfil, D.; Pop, R.; Sisea, R. DNA isolation from desiccated leaf material from plum tree (Prunus domestica L.) molecular analysis. Bull. UASVM Hortic. 2016, 1, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collard, B.C.Y.; Mackill, D.J. Start codon targeted (SCoT) polymorphism: A simple, novel DNA marker technique for generating gene-targeted markers in plants. Plant Mol. Biol Rep. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Serrote, C.M.L.; Reiniger, L.R.S.; Silva, K.B.; dos Santos Rabaiolli, S.M.; Stefanel, C.M. Determining the Polymorphism Information Content of a molecular marker. Gene 2020, 726, 144175. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Tucić, B.; Avramov, S. Maternal effects on early juvenile traits in Iris pumila (Iridaceae). Plant Syst. Evol. 1996, 201, 179–197. [Google Scholar] [CrossRef]
- Santos, E.A.; Souza, M.M.; Viana, A.P.; Almeida, A.A.F.; Freitas, J.C.O.; Lawinscky, P.R. Multivariate analysis of morphological characteristics of two species of passion flower with ornamental potential and of hybrids between them. Genet. Mol. Res. 2011, 10, 2457–2471. [Google Scholar] [CrossRef]
- Ecker, R.; Barzilay, A.; Osherenko, E. Population means and correlation analyses of growth parameters in lisianthus (Eustoma grandiflorum Shinn.). Euphytica 1994, 78, 193–197. [Google Scholar] [CrossRef]
- Oehlkers, F. Cytoplasmic inheritance in the genus Streptocarpus Lindley. Adv. Genet. 1964, 12, 329–370. [Google Scholar] [CrossRef]
- Jung, C.; Müller, A.E. Flowering time control and applications in plant breeding. Trends Plant Sci. 2009, 14, 563–573. [Google Scholar] [CrossRef]
- Scariot, V.; Gullino, P. Evaluation of flowering time and ornamental characteristics in autumn camellias. In I International Symposium on Woody Ornamentals of the Temperate Zone; International Society for Horticultural Science: Leuven, Belgium, 2008; pp. 319–324. [Google Scholar] [CrossRef]
- Kurniati, R.; Yuniarto, K.; Kartikaningrum, S. Morphological diversity of F1 population in Anthurium through conventional breeding. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; p. 012100. [Google Scholar] [CrossRef]
- Lema-Rumińska, J.; Zalewska, M. Changes in flower colour among Lady Group of Chrysanthemum× grandiflorum/Ramat./Kitam. as a result of mutation breeding. Folia Hortic 2005, 17, 61–72. [Google Scholar]
- Gitonga, V.W.; Stolker, R.; Ribot, S.; Keizer, P.; Koning-Boucoiran, C.F.S.; Krens, F.A. Inheritance of determinants of flower colour in tetraploid roses. In XXIII International Eucarpia Symposium, Section Ornamentals: Colourful Breeding and Genetics; International Society for Horticultural Science: Leuven, Belgium, 2009; pp. 55–60. [Google Scholar] [CrossRef] [Green Version]
- Donoso, A.; Rivas, C.; Zamorano, A.; Peña, Á.; Handford, M.; Aros, D. Understanding Alstroemeria pallida flower colour: Links between phenotype, anthocyanins and gene expression. Plants 2020, 10, 55. [Google Scholar] [CrossRef]
- Fan, Z.; Gao, Y.; Guo, L.; Cao, Y.; Liu, R.; Zhang, Q. Phenotypic Variations and Heritability in Hybrid Populations of Bearded Iris. HortScience 2019, 54, 988–992. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, J.; Zhang, Q.; Yu, P.; Zhou, Y.; Jia, G. The Composition of Anthocyanins and Carotenoids Influenced the Flower Color Heredity in Asiatic Hybrid Lilies. Horticulturae 2022, 8, 1206. [Google Scholar] [CrossRef]
- Li, X.; Lu, M.; Tang, D.; Shi, Y. Composition of carotenoids and flavonoids in narcissus cultivars and their relationship with flower color. PLoS ONE 2015, 10, e0142074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeknić, Z.; Jeknić, S.; Jevremović, S.; Subotić, A.; Chen, T.H.H. Alteration of flower color in Iris germanica L. ‘Fire Bride’ through ectopic expression of phytoene synthase gene (crtB) from Pantoea agglomerans. Plant Cell Rep. 2014, 33, 1307–1321. [Google Scholar] [CrossRef] [PubMed]
- Forkmann, G.; Stotz, G. Selection and characterisation of flavanone 3-hydroxylase mutants of Dahlia, Streptocarpus, Verbena and Zinnia. Planta 1984, 161, 261–265. [Google Scholar] [CrossRef]
- Titirică, I.; Roman, I.A.; Nicola, C.; Sturzeanu, M.; Iurea, E.; Botu, M.; Sestras, R.E.; Pop, R.; Militaru, M.; Ercisli, S. The Main Morphological Characteristics and Chemical Components of Fruits and the Possibilities of Their Improvement in Raspberry Breeding. Horticulturae 2023, 9, 50. [Google Scholar] [CrossRef]
- Hayakawa, H.; Muroi, M.; Hamachi, H.; Yokoyama, J.; Fukuda, T. Correlation of variation between leaf and flower characters in Cymbidium goeringii (Rchb. f.) Rchb. f. (Orchidaceae). J. Jap. Bot. 2011, 86, 82–92. [Google Scholar]
- Azimi, M.H.; Karimi Alvijeh, M.; Zarei, A. Intervarietal hybridization and observation of heterosis in the new hybrids of Iris germanica. Int. J. Hortic. Sci. Technol 2018, 5, 65–79. [Google Scholar] [CrossRef]
- Rai, M.K. Start codon targeted (SCoT) polymorphism marker in plant genome analysis: Current status and prospects. Planta 2023, 257, 34. [Google Scholar] [CrossRef]
- Feng, S.-G.; He, R.-F.; Jiang, M.-Y.; Lu, J.-J.; Shen, X.-X.; Liu, J.-J.; Wang, Z.-A.; Wang, H.-Z. Genetic diversity and relationships of medicinal Chrysanthemum morifolium revealed by start codon targeted (SCoT) markers. Sci. Hortic. 2016, 201, 118–123. [Google Scholar] [CrossRef]
- El Khodary, Y.A.; Ayoub, I.M.; El-Ahmady, S.H.; Ibrahim, N. Molecular and phytochemical variability among genus Albizia: A phylogenetic prospect for future breeding. Mol. Biol. Rep. 2021, 48, 2619–2628. [Google Scholar] [CrossRef]
- Li, Q.; Mo, J.; Wu, W.; Yang, J.; Li, J.; Lai, T.; Ou, Z.; Qiu, Z.; Guan, S.; Liao, J. Genetic diversity, population structure and identification of Dendrobium cultivars with high polysaccharide contents using SCoT, SCAR and nested PCR markers. Genet. Resour. Crop. Evol. 2019, 66, 71–88. [Google Scholar] [CrossRef]
- Cai, Y.; Gao, Y.; Zhang, Z.; Liu, H.; Wang, Y.; Ma, Y.; Li, Y.; Feng, S.; Wang, H. Development and Application of a Cultivar-Specific Sequence-Characterized Amplified Region (SCAR) Marker for the Detection of Chrysanthemum morifolium Ramat. ‘Daboju’. Plants 2022, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- Heikrujam, M.; Kumar, J.; Agrawal, V. Genetic diversity analysis among male and female Jojoba genotypes employing gene targeted molecular markers, start codon targeted (SCoT) polymorphism and CAAT box-derived polymorphism (CBDP) markers. Meta Gene 2015, 5, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Heikrujam, M.; Sharma, K.; Prasad, M.; Agrawal, V. Review on different mechanisms of sex determination and sex-linked molecular markers in dioecious crops: A current update. Euphytica 2015, 201, 161–194. [Google Scholar] [CrossRef]
- Clapa, D.; Hârța, M. Establishment of an Efficient Micropropagation System for Humulus lupulus L. cv. Cascade and Confirmation of Genetic Uniformity of the Regenerated Plants through DNA Markers. Agronomy 2021, 11, 2268. [Google Scholar] [CrossRef]
- Arora, K.; Rai, M.K.; Sharma, A.K. Tissue culture mediated biotechnological interventions in medicinal trees: Recent progress. Plant Cell Tissue Organ Cult. 2022, 150, 267–287. [Google Scholar] [CrossRef]
- Zheng, K.; Cai, Y.; Chen, W.; Gao, Y.; Jin, J.; Wang, H.; Feng, S.; Lu, J. Development, identification, and application of a germplasm specific SCAR Marker for Dendrobium officinale Kimura et Migo. Front. Plant Sci. 2021, 12, 669458. [Google Scholar] [CrossRef]
- Cornea-Cipcigan, M.; Pamfil, D.; Sisea, C.R.; Margaoan, R. Characterization of Cyclamen genotypes using morphological descriptors and DNA molecular markers in a multivariate analysis. Front. Plant Sci. 2023, 14, 1100099. [Google Scholar] [CrossRef]
- Yesson, C.; Culham, A. Biogeography of Cyclamen: An application of phyloclimatic modelling. In Climate Change, Ecology and Systematics, Systematics; Association Special Volume Series; Hodkinson, T., Jones, M., Waldren, S., Parnell, J., Eds.; Cambridge University Press: Cambridge, UK, 2011; Volume 78, pp. 265–279. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SV | d.f | Mean Square | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| LL (cm) | NL | LP (cm) | NP | NFP | LCT (cm) | WF (cm) | NF | FT (days) | ||
| Blocks | 5 | 6.794 | 1.563 | 7.178 | 1.546 | 6.962 | 0.704 | 0.339 | 15.236 | 24.611 |
| F-value | 2.474 | 4.656 | 2.817 | 2.460 | 19.470 | 50.938 | 2.650 | 2.800 | 14.210 | |
| p-value | 0.043 | 0.001 | 0.025 | 0.044 | 0.000 | 0.000 | 0.033 | 0.025 | 0.000 | |
| Groups | 11 | 6.849 | 0.698 | 29.085 | 1.262 | 0.735 | 0.028 | 0.313 | 98.037 | 3.613 |
| F-value | 2.494 | 2.079 | 11.414 | 2.007 | 2.054 | 1.998 | 2.447 | 18.016 | 2.086 | |
| p-value | 0.013 | 0.038 | 0.000 | 0.046 | 0.040 | 0.047 | 0.015 | 0.000 | 0.037 | |
| Error | 54 | 2.746 | 0.336 | 2.548 | 0.629 | 0.358 | 0.014 | 0.128 | 5.442 | 1.732 |
| Total | 70 | |||||||||
| Morphological Parameters | P1 Mean ± SD | P2 Mean ± SD | P1×P2 Mean ± SD | P2×P1 Mean ± SD |
|---|---|---|---|---|
| Length of leaves/plant (cm) | 25.19 ± 0.25 d | 16.85 ± 0.07 a | 20.48 ± 1.2 b | 22.54 ± 0.95 c ** |
| Number of leaves/plant | 10.53 ± 0.28 b | 9.87 ± 0.11 a | 11.05 ± 0.85 c * | 11.34 ± 0.76 c * |
| Length of peduncle (cm) | 18.14 ± 0.26 c | 14.33 ± 0.11 a | 15.82 ± 0.33 b | 20.48 ± 0.71 d * |
| Number of peduncle/plant | 6.77 ± 0.29 a | 8.59 ± 0.27 b | 8.99 ± 0.77 bc * | 9.46 ± 0.46 c * |
| Number of flowers/peduncle | 3.11 ± 0.04 b | 3.89 ± 0.13 c | 4.06 ± 0.74 c * | 2.72 ± 0.23 a |
| Length of corolla tube (cm) | 3.07 ± 0.10 c | 2.91 ± 0.04 b | 3.23 ± 0.12 d * | 2.78 ± 0.06 a |
| Width of flower (cm) | 5.96 ± 0.07 b | 3.43 ± 0.06 a | 5.72 ± 0.31 b ** | 5.67 ± 0.28 b ** |
| Number of flowers/plant | 21.04 ± 0.93 a | 33.43 ± 1.88 b | 36.19 ± 5.70 bc * | 23.93 ± 2.93 a |
| Flowering time (days) | 21.20 ± 0.13 b | 27.70 ± 0.39 c | 17.95 ± 0.38 a | 21.05 ± 0.81 b |
| Parental Cross | Primer Name | Size of Bands (bp) | NPB | NTB | PPB | PIC |
|---|---|---|---|---|---|---|
| P1×P2 | SCoT 1 | 450–6000 | 9 | 11 | 81.81 | 0.46 |
| SCoT 3 | 380–6500 | 15 | 17 | 88.23 | 0.47 | |
| SCoT 6 | 350–6400 | 16 | 18 | 88.88 | 0.47 | |
| SCoT 7 | 400–8200 | 17 | 20 | 85.00 | 0.49 | |
| SCoT 9 | 300–6300 | 12 | 14 | 85.71 | 0.48 | |
| SCoT 12 | 400–5000 | 13 | 15 | 86.66 | 0.48 | |
| SCoT 13 | 450–5500 | 13 | 15 | 86.66 | 0.47 | |
| SCoT 14 | 350–5500 | 16 | 18 | 88.88 | 0.47 | |
| SCoT 15 | 350–4000 | 16 | 19 | 84.21 | 0.48 | |
| Total | 127 | 147 | ||||
| Mean | 14.11 ± 0.86 | 16.33 ± 0.94 | 86.23 ± 0.78 | 0.47 ± 0.01 | ||
| P2×P1 | SCoT 1 | 400–6000 | 9 | 10 | 90.00 | 0.48 |
| SCoT 3 | 350–6500 | 15 | 17 | 88.23 | 0.47 | |
| SCoT 6 | 350–6500 | 15 | 17 | 88.23 | 0.48 | |
| SCoT 7 | 400–8000 | 17 | 19 | 89.47 | 0.49 | |
| SCoT 9 | 300–6000 | 10 | 12 | 83.33 | 0.49 | |
| SCoT 12 | 500–5000 | 12 | 14 | 85.71 | 0.48 | |
| SCoT 13 | 450–4000 | 11 | 13 | 84.61 | 0.48 | |
| SCoT 14 | 300–5000 | 15 | 17 | 88.23 | 0.48 | |
| SCoT 15 | 300–4000 | 17 | 19 | 89.47 | 0.46 | |
| Total | 121 | 138 | ||||
| Mean | 13.44 ± 1.01 | 15.33 ± 1.06 | 87.21 ± 0.89 | 0.48 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hârţa, M.; Clapa, D.; Cornea-Cipcigan, M.; Borsai, O.; Pop, R.; Cordea, M.I. Multivariate Assessment of Genetic Relationships between Two Streptocarpus Cultivars and Their F1 Progenies Using Morphological Characteristics and SCoT Molecular Markers. Horticulturae 2023, 9, 497. https://doi.org/10.3390/horticulturae9040497
Hârţa M, Clapa D, Cornea-Cipcigan M, Borsai O, Pop R, Cordea MI. Multivariate Assessment of Genetic Relationships between Two Streptocarpus Cultivars and Their F1 Progenies Using Morphological Characteristics and SCoT Molecular Markers. Horticulturae. 2023; 9(4):497. https://doi.org/10.3390/horticulturae9040497
Chicago/Turabian StyleHârţa, Monica, Doina Clapa, Mihaiela Cornea-Cipcigan, Orsolya Borsai, Rodica Pop, and Mirela Irina Cordea. 2023. "Multivariate Assessment of Genetic Relationships between Two Streptocarpus Cultivars and Their F1 Progenies Using Morphological Characteristics and SCoT Molecular Markers" Horticulturae 9, no. 4: 497. https://doi.org/10.3390/horticulturae9040497
APA StyleHârţa, M., Clapa, D., Cornea-Cipcigan, M., Borsai, O., Pop, R., & Cordea, M. I. (2023). Multivariate Assessment of Genetic Relationships between Two Streptocarpus Cultivars and Their F1 Progenies Using Morphological Characteristics and SCoT Molecular Markers. Horticulturae, 9(4), 497. https://doi.org/10.3390/horticulturae9040497