


Adaptive Circadian Rhythms for Autonomous and Biologically Inspired Robot Behavior

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
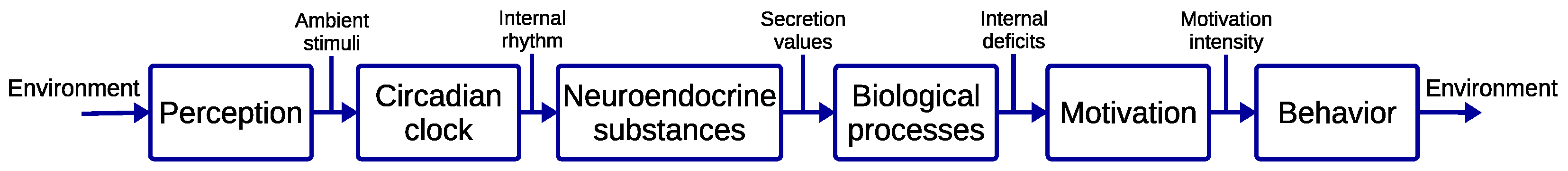
2. From Human to Robot Biological Rhythms
2.1. Human Biological Rhythms
2.2. Biological Rhythms in Robots
3. Modeling Biological Rhythms
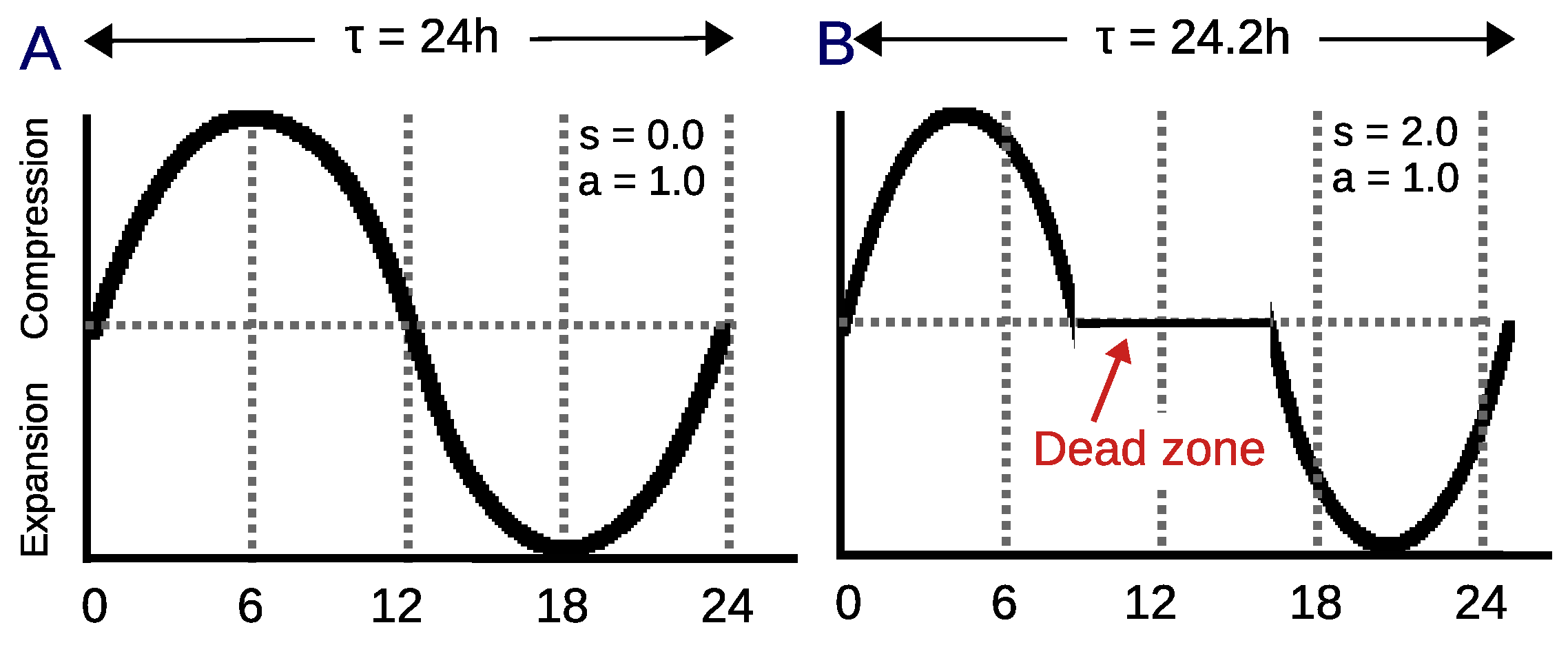
3.1. Dynamic Circadian Integrated Response Characteristic
3.2. Biologically Inspired Model
4. Experimental Setup
4.1. Scenarios
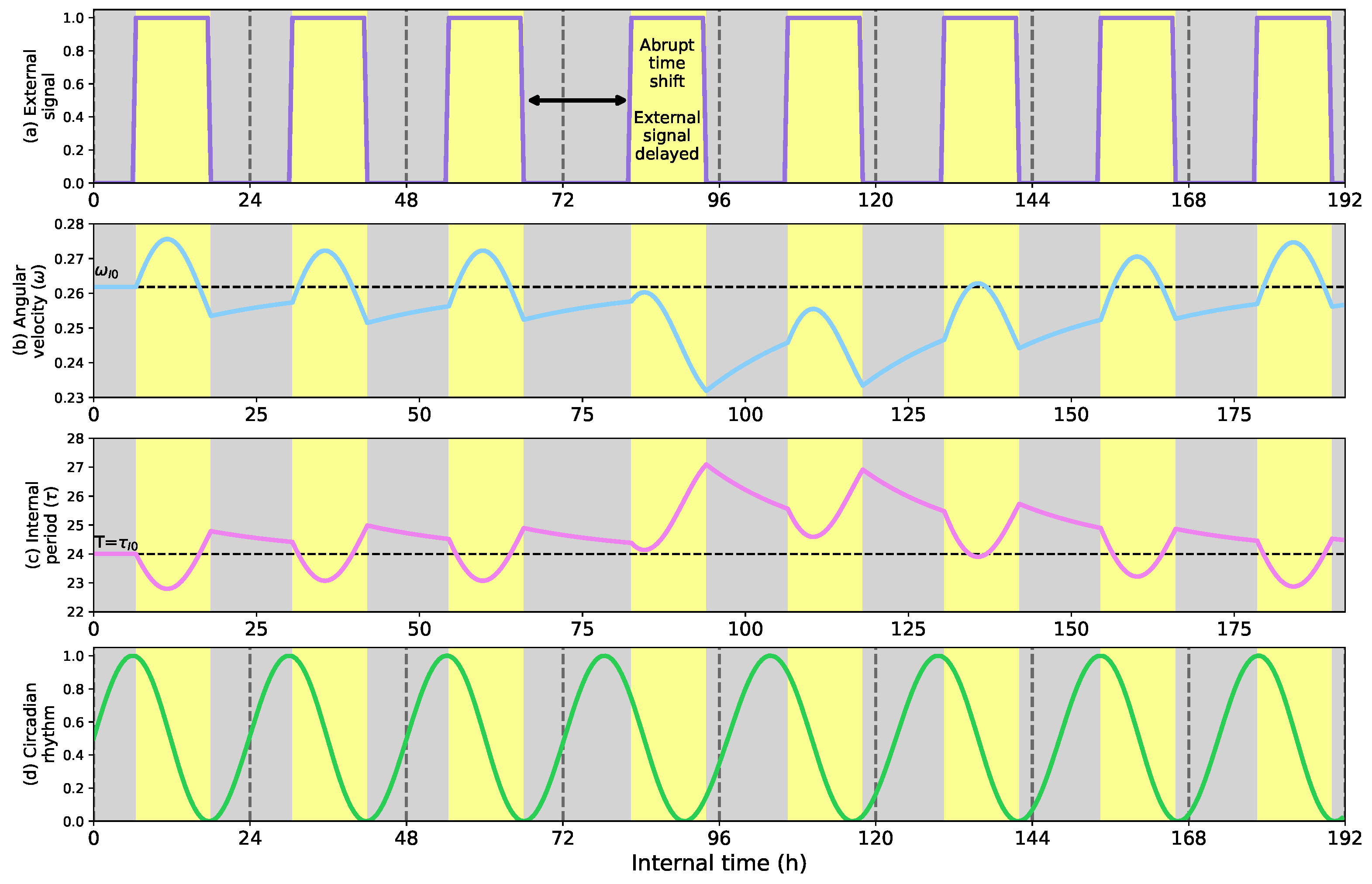
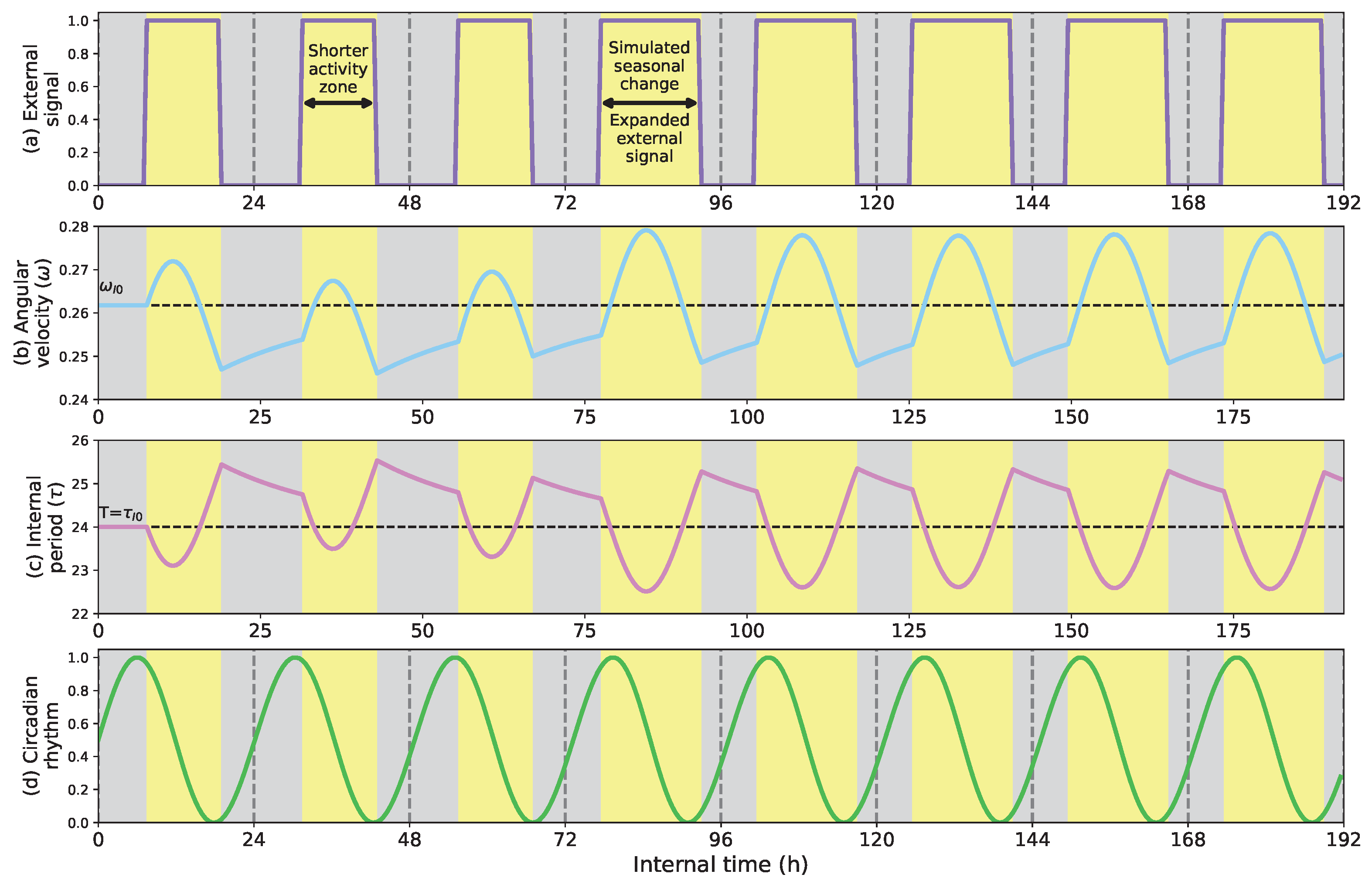
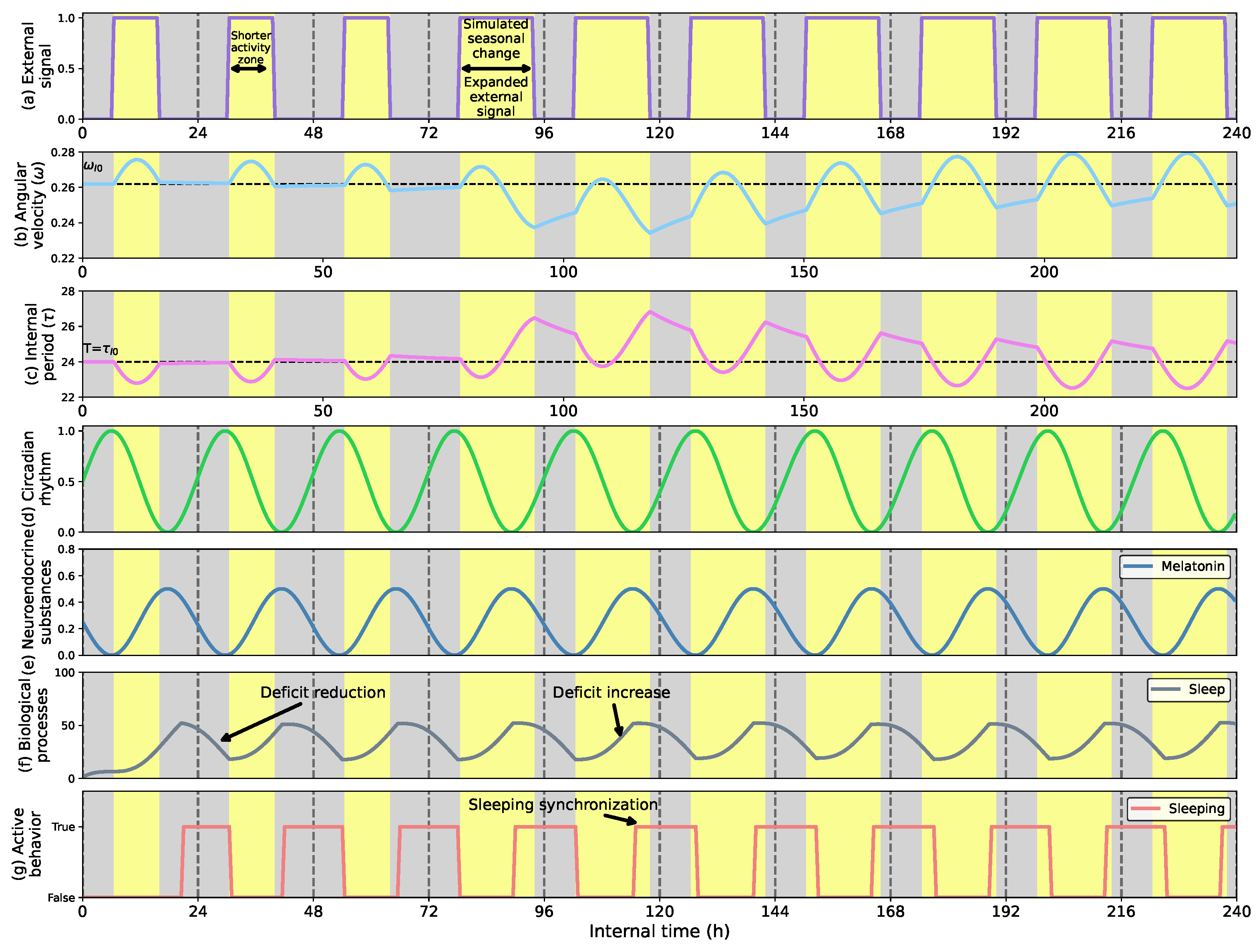
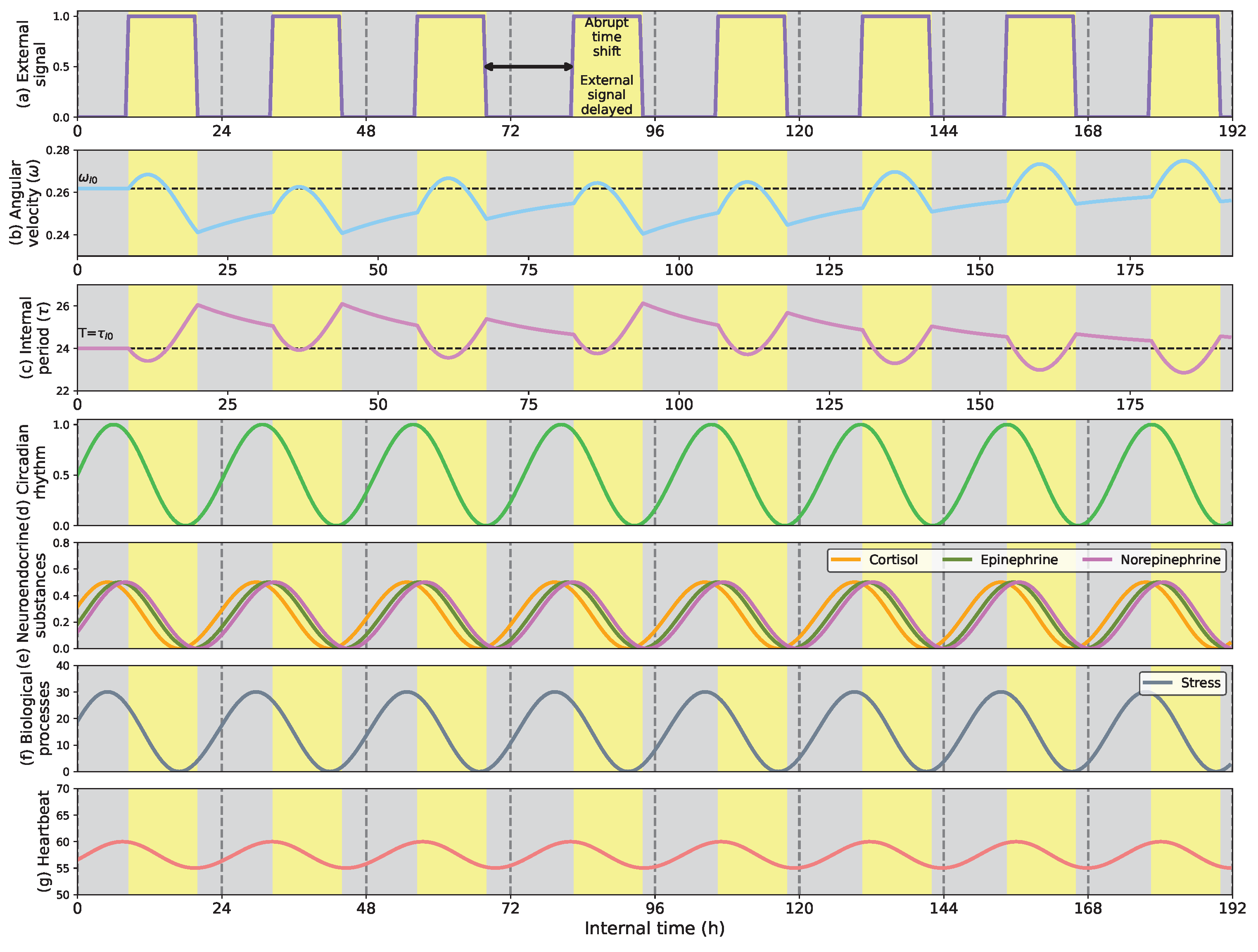
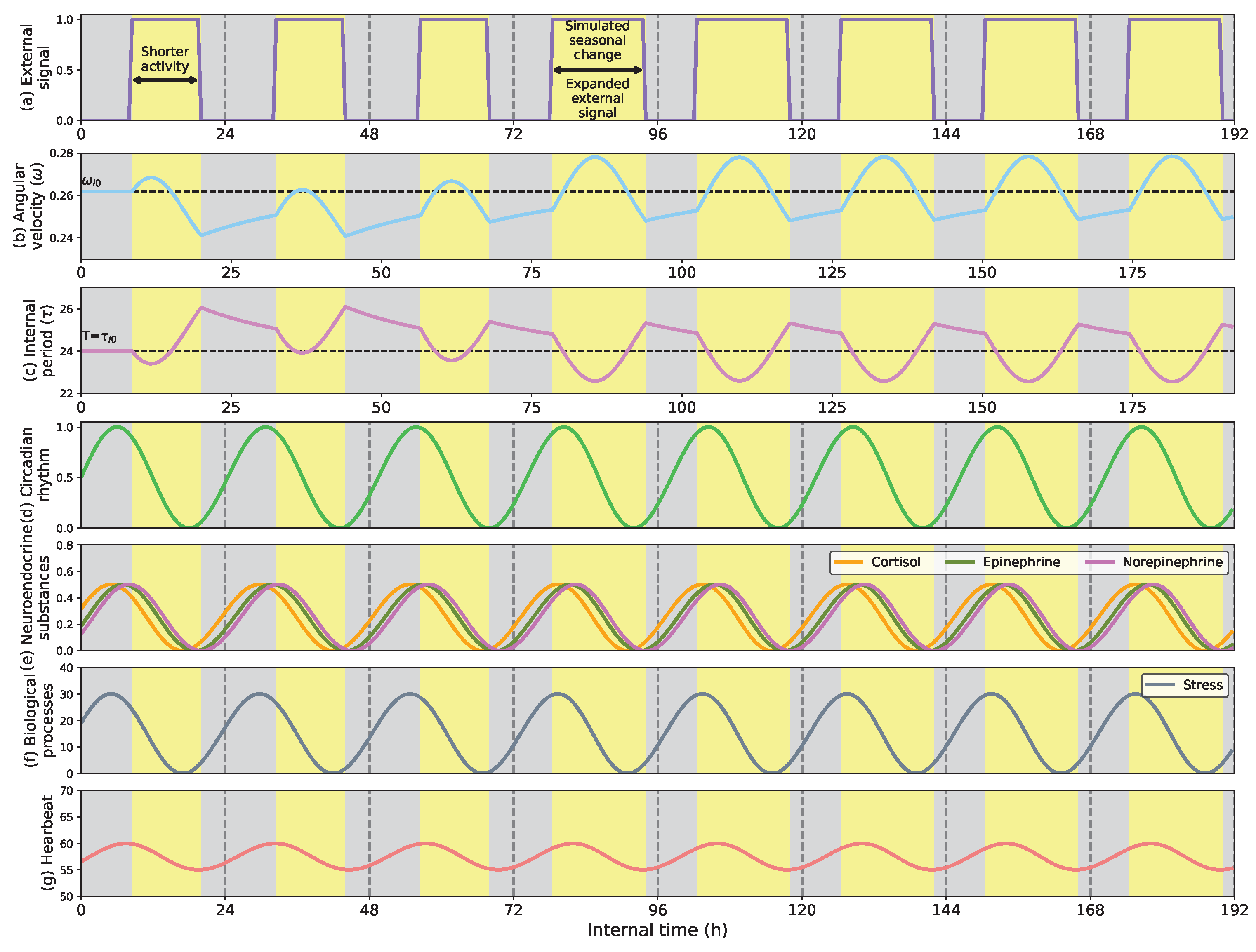
- Show how the circadian clock adapts time and angular velocity to changing environmental conditions. We demonstrate its performance in two cases: abrupt time shifts and seasonal changes in the External signal.
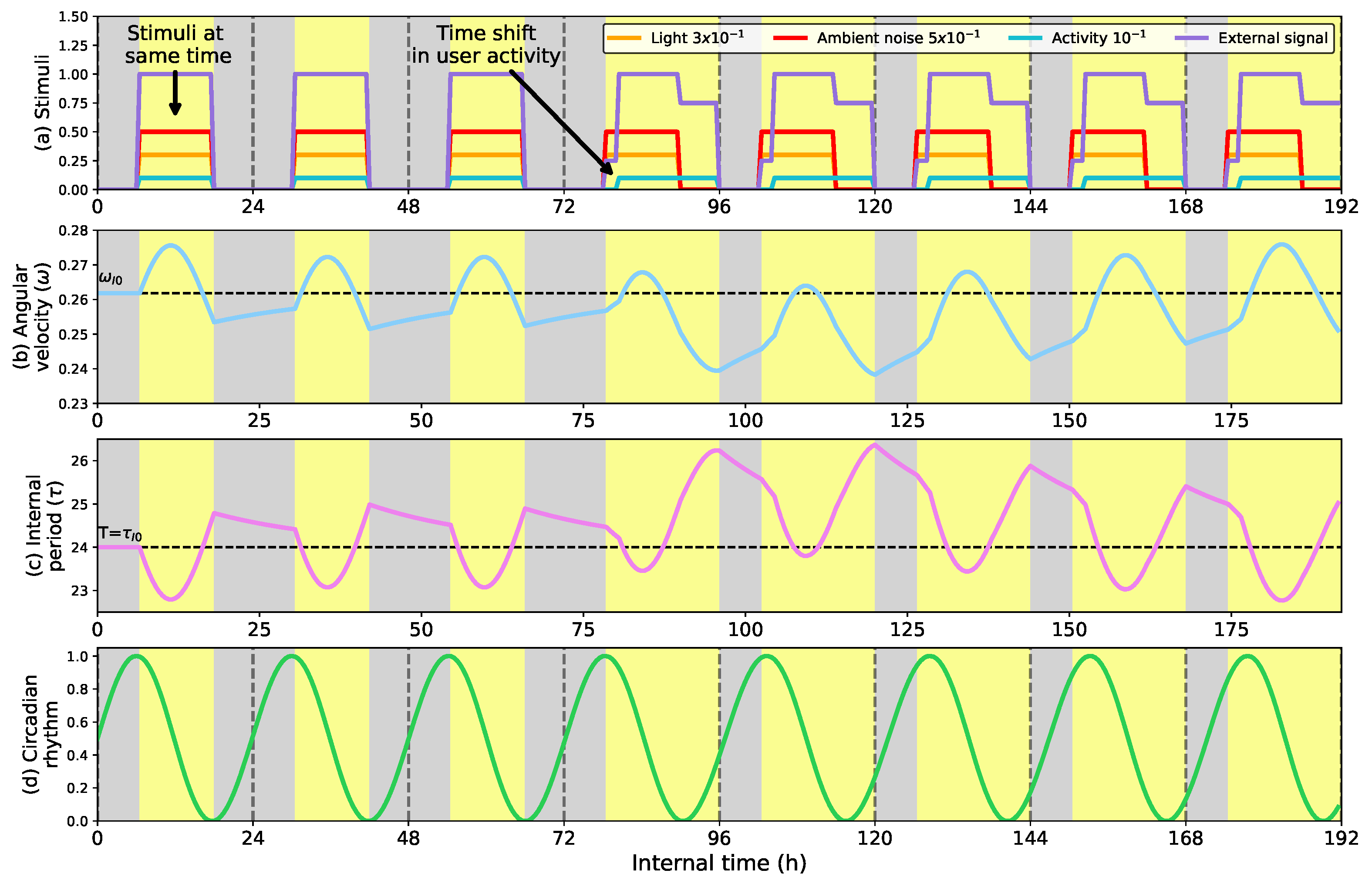
- Show the impact that ambient stimuli have on the circadian clock. Specifically, we show how giving more importance to the user activity can drive the robot to shift its activity to match that of the user.
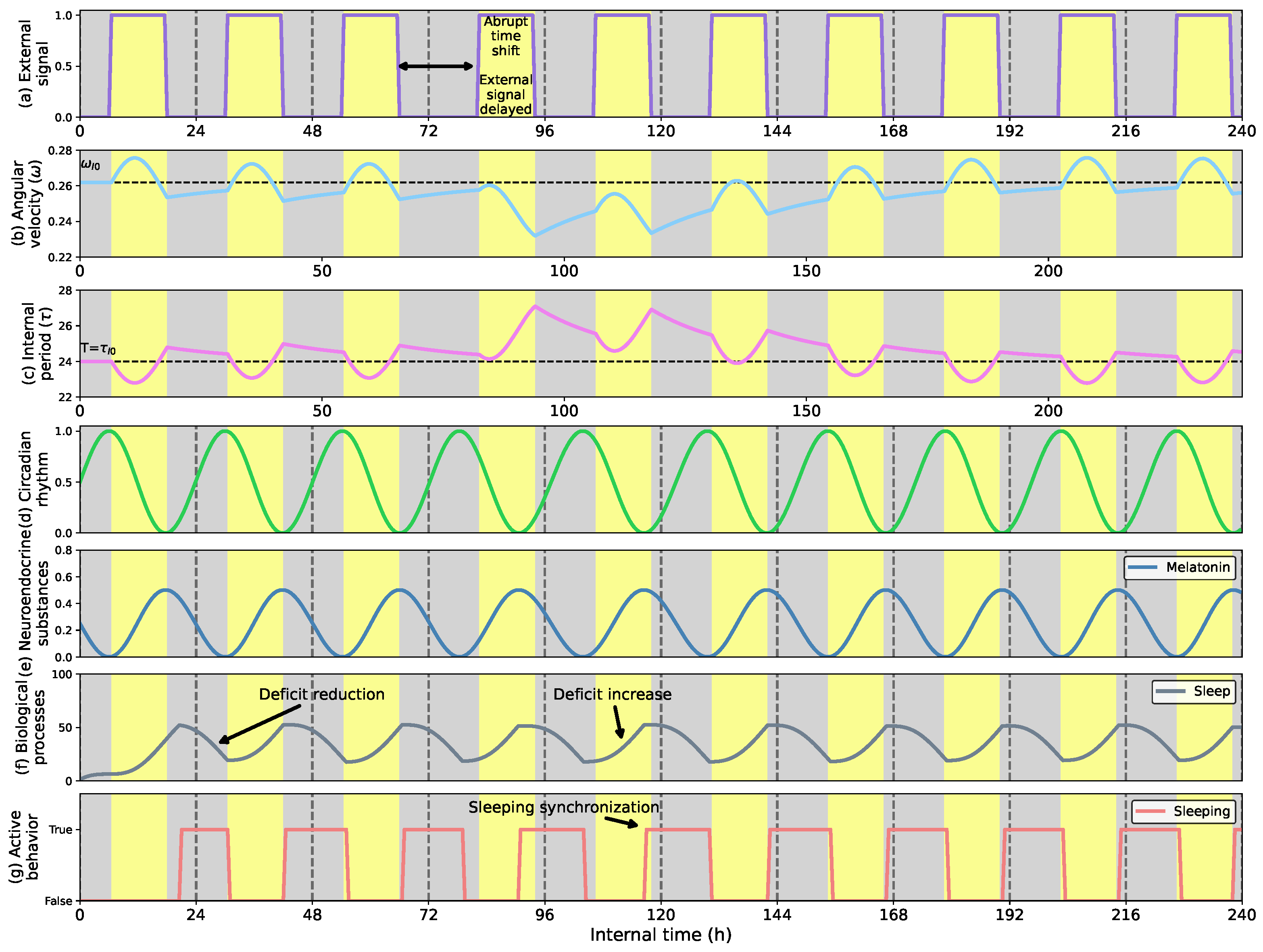
- Demonstrate how the robot can dynamically adjust long-lasting voluntary behaviors like the sleep-wake cycle to external conditions that make it change its activity periods.
- Demonstrate how the circadian clock drives involuntary behaviors like heartbeat rhythm through the action of other processes like stress peak in the early morning.
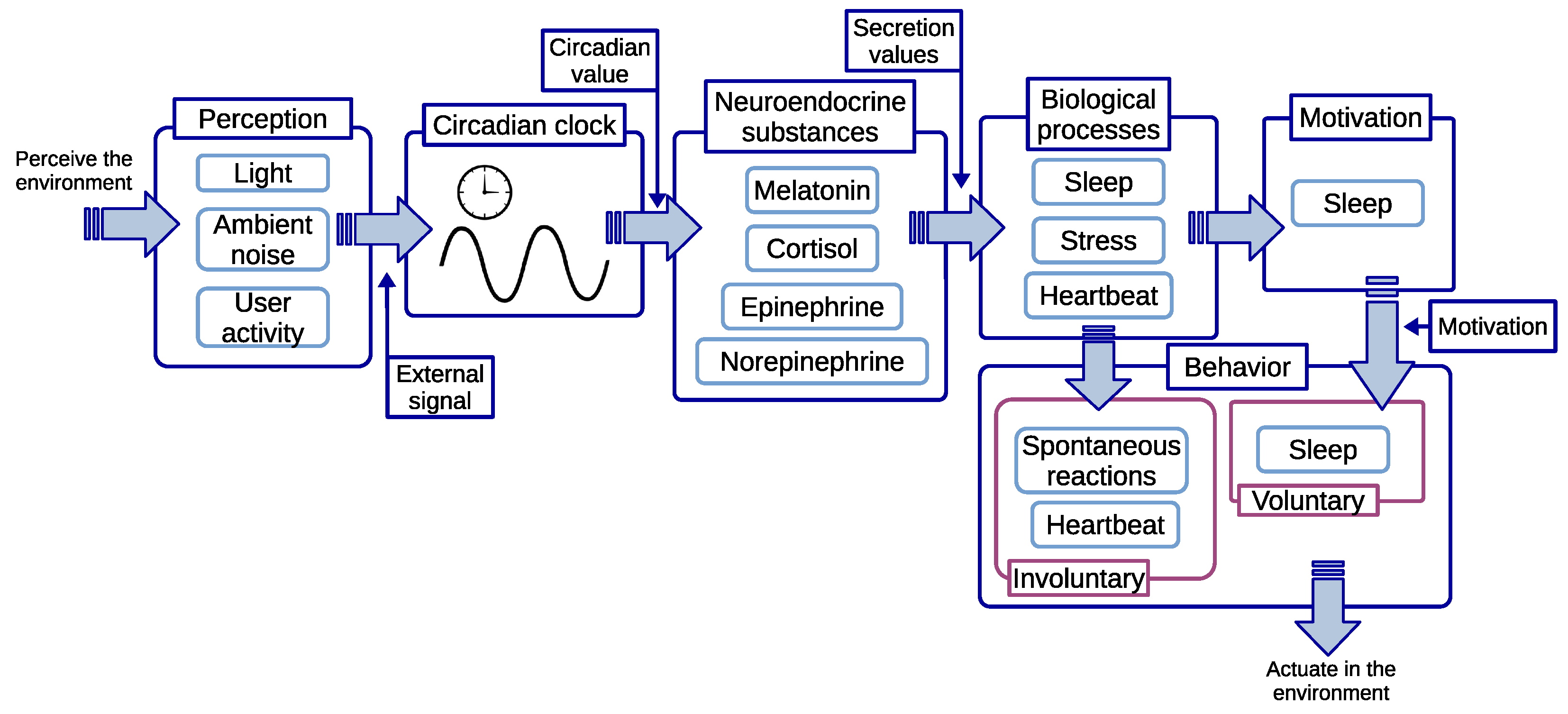
4.2. Circadian Definition
4.3. Stimuli
4.4. Neuroendocrine Substances
4.5. Biological Processes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Process | Basal Value () | Related Substances | Ideal Value () | ||
|---|---|---|---|---|---|
| Sleep | 0 | 1 | 5 | Melatonin | 0 |
| Stress | 0 | 0 | 10 | Cortisol | 0 |
| Heartbeat | 60 | 0 | 10 | Epinephrine Norepinephrine | 60 |
4.6. Motivation and Behavior
5. Results
- First, we describe how the circadian clock adjusts its time and angular velocity by reacting to abrupt time shifts and simulated seasonal changes in the ambient stimuli.
- Second, we show how the circadian clock adapts to different external stimuli (light, ambient noise, and activity), showing how the clock makes the robot more aware of the user’s activity.
- Third, we show how adaptive voluntary behavior emerges following the dynamics of the biological model to control the sleep-wake cycle.
- Fourth, we present how the model deals with adaptive involuntary behavior, showing the adaptation of stress and heartbeat to ambient conditions to maintain their periodic peaks in the early morning h.
5.1. Phase Shifts and Seasonal Changes in the Circadian Clock
5.2. Influence of Ambient Stimuli on the Clock
5.3. Controlling Cyclic Behavior: Sleep-Wake Case
5.4. Arousal Effects and the Circadian Clock
6. Discussion
6.1. Adapting the Master Clock
6.2. Regulating Long-Lasting Behavior
6.3. Limitations
- Model parameters: The primary drawback of the biological model presented in this paper is the high number of parameters that play a role in the circadian clock and the rest of biological functions like neuroendocrine substances and biological processes. These parameters affect the model performance and, therefore, the behavior exhibited by the robot, so obtaining them from real studies can be an alternative to produce more realistic biologically inspired systems.
- Scalability: Modeling biologically inspired phenomena in artificial agents like robots requires precise synchronization tuning. The emulation of biological processes affects the system’s scalability, increasing its complexity and dedication as the number of processes grows. This problem is important because as the number of robot behaviors increases, the users’ perception also improves. To overcome this limitation, it is essential to develop general methods that simplify the definition of the biological functions proposed in the robot. This paper proposes new advances like using the dCiRC model and methods to characterize motivation, neuroendocrine substances, and other biological processes from this rhythm.
- Robot hardware requirements: We would like to integrate and evaluate the model using our Mini social robot [37] to endow it with more natural and adaptive behavior. Mini possesses sensors and actuators to perceive stimuli and execute behavior. However, their performance can be affected if the model is applied to other robots or systems lacking the necessary hardware devices. We believe that the sensors for perceiving ambient light intensity or noise are widely available nowadays, so robots can easily integrate them into other robots to use the model.
- User evaluation: The generation of biologically inspired and adaptive behavior needs an evaluation from the users. In the future, we plan to extend the number of processes integrated into the model to obtain the users’ impressions of the robot’s performance.
7. New Insights and Potential Applications
8. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gerhart-Hines, Z.; Lazar, M.A. Circadian metabolism in the light of evolution. Endocr. Rev. 2015, 36, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Field, J.M.; Sehgal, A. Circadian rhythms, disease and chronotherapy. J. Biol. Rhythm. 2022, 36, 503–531. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.A.; St. Hilaire, M.A.; Grant, L.K.; Barger, L.K.; Brainard, G.C.; Czeisler, C.A.; Klerman, E.B.; Lockley, S.W. Dynamic lighting schedules to facilitate circadian adaptation to shifted timing of sleep and wake. J. Pineal Res. 2022, 73, e12805. [Google Scholar] [CrossRef] [PubMed]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. The mammalian circadian timing system and the suprachiasmatic nucleus as its pacemaker. Biology 2019, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.; Gumz, M.L. Advances in understanding the peripheral circadian clocks. FASEB J. 2012, 26, 3602–3613. [Google Scholar] [CrossRef] [PubMed]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445. [Google Scholar] [CrossRef]
- Pessoa, L. Intelligent architectures for robotics: The merging of cognition and emotion. Phys. Life Rev. 2019, 31, 157–170. [Google Scholar] [CrossRef]
- Gervasi, R.; Barravecchia, F.; Mastrogiacomo, L.; Franceschini, F. Applications of affective computing in human-robot interaction: State-of-art and challenges for manufacturing. Proc. Inst. Mech. Eng. Part J. Eng. Manuf. 2022, 237, 815–832. [Google Scholar] [CrossRef]
- Moro, C.; Lin, S.; Nejat, G.; Mihailidis, A. Social robots and seniors: A comparative study on the influence of dynamic social features on human–robot interaction. Int. J. Soc. Robot. 2019, 11, 5–24. [Google Scholar] [CrossRef]
- An, Z.; Piccoli, B.; Merrow, M.; Lee, K. A Unified Model for Entrainment by Circadian Clocks: Dynamic Circadian Integrated Response Characteristic (dCiRC). J. Biol. Rhythm. 2022, 37, 202–215. [Google Scholar] [CrossRef]
- Roenneberg, T.; Rémi, J.; Merrow, M. Modeling a circadian surface. J. Biol. Rhythm. 2010, 25, 340–349. [Google Scholar] [CrossRef]
- Fernández-Rodicio, E.; Maroto-Gómez, M.; Castro-González, Á.; Malfaz, M.; Salichs, M.Á. Emotion and mood blending in embodied artificial agents: Expressing affective states in the mini social robot. Int. J. Soc. Robot. 2022, 14, 1841–1864. [Google Scholar] [CrossRef]
- Maroto-Gómez, M.; Castro-González, Á.; Castillo, J.C.; Malfaz, M.; Salichs, M.Á. An adaptive decision-making system supported on user preference predictions for human–robot interactive communication. User Model. User-Adapt. Interact. 2023, 33, 359–403. [Google Scholar] [CrossRef]
- O’Madagain, C.; Tomasello, M. Shared intentionality, reason-giving and the evolution of human culture. Philos. Trans. R. Soc. B 2022, 377, 20200320. [Google Scholar] [CrossRef] [PubMed]
- Reinberg, A.; Ashkenazi, I. Concepts in human biological rhythms. Dialogues Clin. Neurosci. 2022, 5, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Tähkämö, L.; Partonen, T.; Pesonen, A.K. Systematic review of light exposure impact on human circadian rhythm. Chronobiol. Int. 2019, 36, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Blume, C.; Garbazza, C.; Spitschan, M. Effects of light on human circadian rhythms, sleep and mood. Somnologie 2019, 23, 147–156. [Google Scholar] [CrossRef]
- Duffy, J.F.; Kronauer, R.E.; Czeisler, C.A. Phase-shifting human circadian rhythms: Influence of sleep timing, social contact and light exposure. J. Physiol. 1996, 495, 289–297. [Google Scholar] [CrossRef]
- Asgari-Targhi, A.; Klerman, E.B. Mathematical modeling of circadian rhythms. Wiley Interdiscip. Rev. Syst. Biol. Med. 2019, 11, e1439. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, S.; Deng, X.; Liang, J. A Circadian Rhythms Neural Network for Solving the Redundant Robot Manipulators Tracking Problem Perturbed by Periodic Noise. IEEE/ASME Trans. Mechatr. 2021, 26, 3232–3242. [Google Scholar] [CrossRef]
- Arkin, R.C.; Fujita, M.; Takagi, T.; Hasegawa, R. Ethological modeling and architecture for an entertainment robot. In Proceedings of the 2001 ICRA, IEEE International Conference on Robotics and Automation (Cat. No. 01CH37164), IEEE, Seoul, Republic of Korea, 21–26 May 2001; Volume 1, pp. 453–458. [Google Scholar]
- Stoytchev, A.; Arkin, R.C. Incorporating motivation in a hybrid robot architecture. J. Adv. Comput. Intell. Vol. 2004, 8, 100–105. [Google Scholar] [CrossRef]
- Sauzé, C.; Neal, M. A biologically inspired approach to long term autonomy and survival in sailing robots. In Proceedings of the IRSC 2008 International Robotic Sailing Conference, Breitenbrunn, Austria, 20–25 May 2008; p. 6. [Google Scholar]
- Burattini, E.; Rossi, S. Periodic adaptive activation of behaviors in robotic systems. Int. J. Pattern Recognit. Artif. Intell. 2008, 22, 987–999. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Arkin, R.C. An Artificial Circadian System for a Slow and Persistent Robot. In Proceedings of the International Conference on Simulation of Adaptive Behavior, Frankfurt, Germany, 14–17 August 2018; Springer: Berlin/Heidelberg, Germany, 2018; pp. 149–161. [Google Scholar]
- Ito, H.; Sugi, T.; Nagai, K.H. Controllable Biological Rhythms and Patterns. J. Robot. Mechatronics 2022, 34, 253–256. [Google Scholar] [CrossRef]
- Maroto-Gómez, M.; Castro-González, Á.; Malfaz, M.; Fernández-Rodicio, E.; Salichs, M.Á. Modeling Neuroendocrine Autonomic Responses in Embodied Autonomous Robots. Adv. Intell. Syst. 2022, 5, 2200288. [Google Scholar] [CrossRef]
- Lu, Q.; Kim, J.Y. Mammalian circadian networks mediated by the suprachiasmatic nucleus. FEBS J. 2022, 289, 6589–6604. [Google Scholar] [CrossRef] [PubMed]
- Ono, D.; Wang, H.; Hung, C.J.; Wang, H.T.; Kon, N.; Yamanaka, A.; Li, Y.; Sugiyama, T. Network-driven intracellular cAMP coordinates circadian rhythm in the suprachiasmatic nucleus. Sci. Adv. 2023, 9, eabq7032. [Google Scholar] [CrossRef] [PubMed]
- Gnocchi, D.; Bruscalupi, G. Circadian rhythms and hormonal homeostasis: Pathophysiological implications. Biology 2017, 6, 10. [Google Scholar] [CrossRef]
- Poza, J.J.; Pujol, M.; Ortega-Albás, J.J.; Romero, O. Melatonin in sleep disorders. Neurologia 2022, 37, 575–585. [Google Scholar] [CrossRef]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2022, 8, 383–395. [Google Scholar] [CrossRef]
- Froemke, R.C.; Young, L.J. Oxytocin, neural plasticity, and social behavior. Annu. Rev. Neurosci. 2021, 44, 359–381. [Google Scholar] [CrossRef]
- Deschaine, S.L.; Leggio, L. From “hunger hormone” to “it’s complicated”: Ghrelin beyond feeding control. Physiology 2022, 37, 5–15. [Google Scholar] [CrossRef]
- Lewis, M.; Canamero, L. A robot model of stress-induced compulsive behavior. In Proceedings of the 2019 8th International Conference on Affective Computing and Intelligent Interaction (ACII), IEEE, Cambridge, UK, 3–6 September 2019; pp. 559–565. [Google Scholar]
- Nogo, D.; Jasrai, A.K.; Kim, H.; Nasri, F.; Ceban, F.; Lui, L.M.; Rosenblat, J.D.; Vinberg, M.; Ho, R.; McIntyre, R.S. The effect of ketamine on anhedonia: Improvements in dimensions of anticipatory, consummatory, and motivation-related reward deficits. Psychopharmacology 2022, 239, 2011–2039. [Google Scholar] [CrossRef]
- Salichs, M.A.; Castro-González, Á.; Salichs, E.; Fernández-Rodicio, E.; Maroto-Gómez, M.; Gamboa-Montero, J.J.; Marques-Villarroya, S.; Castillo, J.C.; Alonso-Martín, F.; Malfaz, M. Mini: A new social robot for the elderly. Int. J. Soc. Robot. 2020, 12, 1231–1249. [Google Scholar] [CrossRef]
- Bougouin, W.; Slimani, K.; Renaudier, M.; Binois, Y.; Paul, M.; Dumas, F.; Lamhaut, L.; Loeb, T.; Ortuno, S.; Deye, N.; et al. Epinephrine versus norepinephrine in cardiac arrest patients with post-resuscitation shock. Intensive Care Med. 2022, 48, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Joo, Y.; Kim, M.S.; Choe, H.K.; Tong, Q.; Kwon, O. Effects of intermittent fasting on the circulating levels and circadian rhythms of hormones. Endocrinol. Metab. 2021, 36, 745–756. [Google Scholar] [CrossRef]
- Golder, S.A.; Macy, M.W. Diurnal and seasonal mood vary with work, sleep, and daylength across diverse cultures. Science 2011, 333, 1878–1881. [Google Scholar] [CrossRef] [PubMed]











| Abbreviation | Model Use | Meaning | Range and Unit |
|---|---|---|---|
| Circadian clock | Phase of the external clock | [] radians | |
| Circadian clock | Phase of the internal clock | [] radians | |
| CiRC | Circadian clock | Circadian value in time | [0, 100] units |
| s | Circadian clock | Shape factor | [] units |
| a | Circadian clock | Asymmetry factor | [] units |
| Circadian clock | Period of the internal clock | (] h | |
| T | Circadian clock | Period of the External signal | (] h |
| Circadian clock | Angular velocity of the internal clock | (] radians per h | |
| Circadian clock | Angular velocity in constant darkness | (] radians per h | |
| Circadian clock | Impact of the stimuli on CiRC value | (] units | |
| Circadian clock | Elastic factor to correct the angular velocity | (] units | |
| External signal | Circadian clock | Intensity of stimuli | [] units |
| External signal | Impact of light on clock | [] units | |
| External signal | Impact of noise on clock | [] units | |
| External signal | Impact of the robot activity on clock | [] units | |
| light | External signal | Ambient light intensity | [] units |
| noise | External signal | Ambient noise intensity | [] units |
| activity | External signal | User activity value | [] units |
| Neuroendocrine subs. | Value of neuroendocrine substance n | [] units | |
| Neuroendocrine subs. | Impact of CiRC value on substance n | [] units | |
| Neuroendocrine subs. | Phase shift of substance n | [] radians | |
| Biological processes | Value of biological process p | [] units | |
| Biological processes | Basal value of process p | [] units | |
| Biological processes | Accumulative effect of process p | 0 or 1 unit | |
| Biological processes | Impact of substance n on process p | [] units | |
| Deficit | Deficit of a biological process p | [] units | |
| Deficit | Ideal value of a biological process | [] units | |
| Motivation | Threshold value for motivation k | [] units | |
| Motivation | Intensity of motivation k | [] units |
| Neuroendocrine Substance | Phase Shift () | Effects | |
|---|---|---|---|
| Melatonin | 12 h × 2 / | Induces sleep | |
| Cortisol | 23 h × 2 / | Stress regulation | |
| Epinephrine | 1 h× 2 / | Increased heartbeat | |
| Norepinephrine | 2 h× 2 / | Increased heartbeat |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maroto-Gómez, M.; Malfaz, M.; Castro-González, Á.; Carrasco-Martínez, S.; Salichs, M.Á. Adaptive Circadian Rhythms for Autonomous and Biologically Inspired Robot Behavior. Biomimetics 2023, 8, 413. https://doi.org/10.3390/biomimetics8050413
Maroto-Gómez M, Malfaz M, Castro-González Á, Carrasco-Martínez S, Salichs MÁ. Adaptive Circadian Rhythms for Autonomous and Biologically Inspired Robot Behavior. Biomimetics. 2023; 8(5):413. https://doi.org/10.3390/biomimetics8050413
Chicago/Turabian StyleMaroto-Gómez, Marcos, María Malfaz, Álvaro Castro-González, Sara Carrasco-Martínez, and Miguel Ángel Salichs. 2023. "Adaptive Circadian Rhythms for Autonomous and Biologically Inspired Robot Behavior" Biomimetics 8, no. 5: 413. https://doi.org/10.3390/biomimetics8050413
APA StyleMaroto-Gómez, M., Malfaz, M., Castro-González, Á., Carrasco-Martínez, S., & Salichs, M. Á. (2023). Adaptive Circadian Rhythms for Autonomous and Biologically Inspired Robot Behavior. Biomimetics, 8(5), 413. https://doi.org/10.3390/biomimetics8050413