
In Vitro Inhibition of Influenza Virus Using CRISPR/Cas13a in Chicken Cells


Abstract
:1. Introduction
2. Materials and Methods
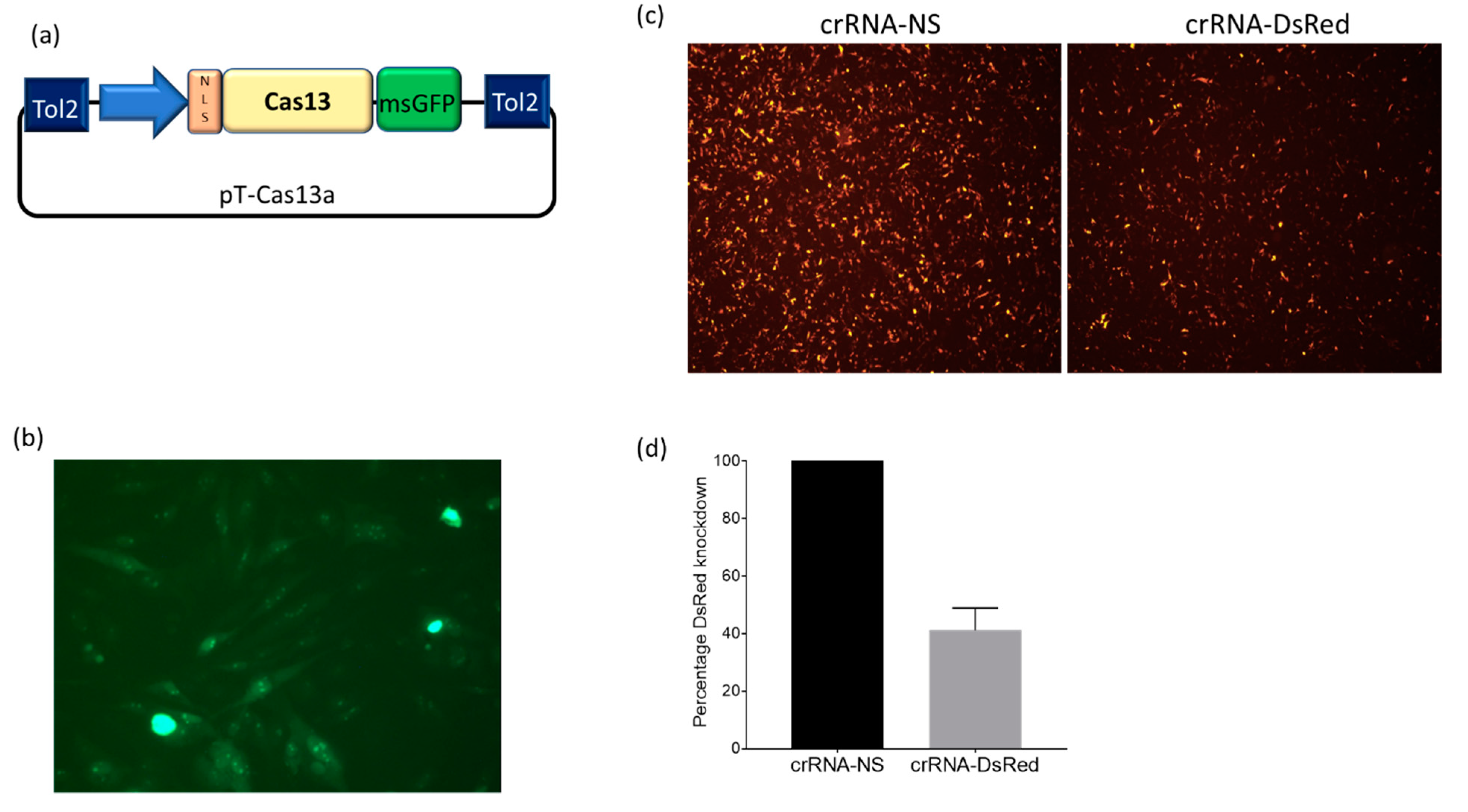
2.1. Plasmids
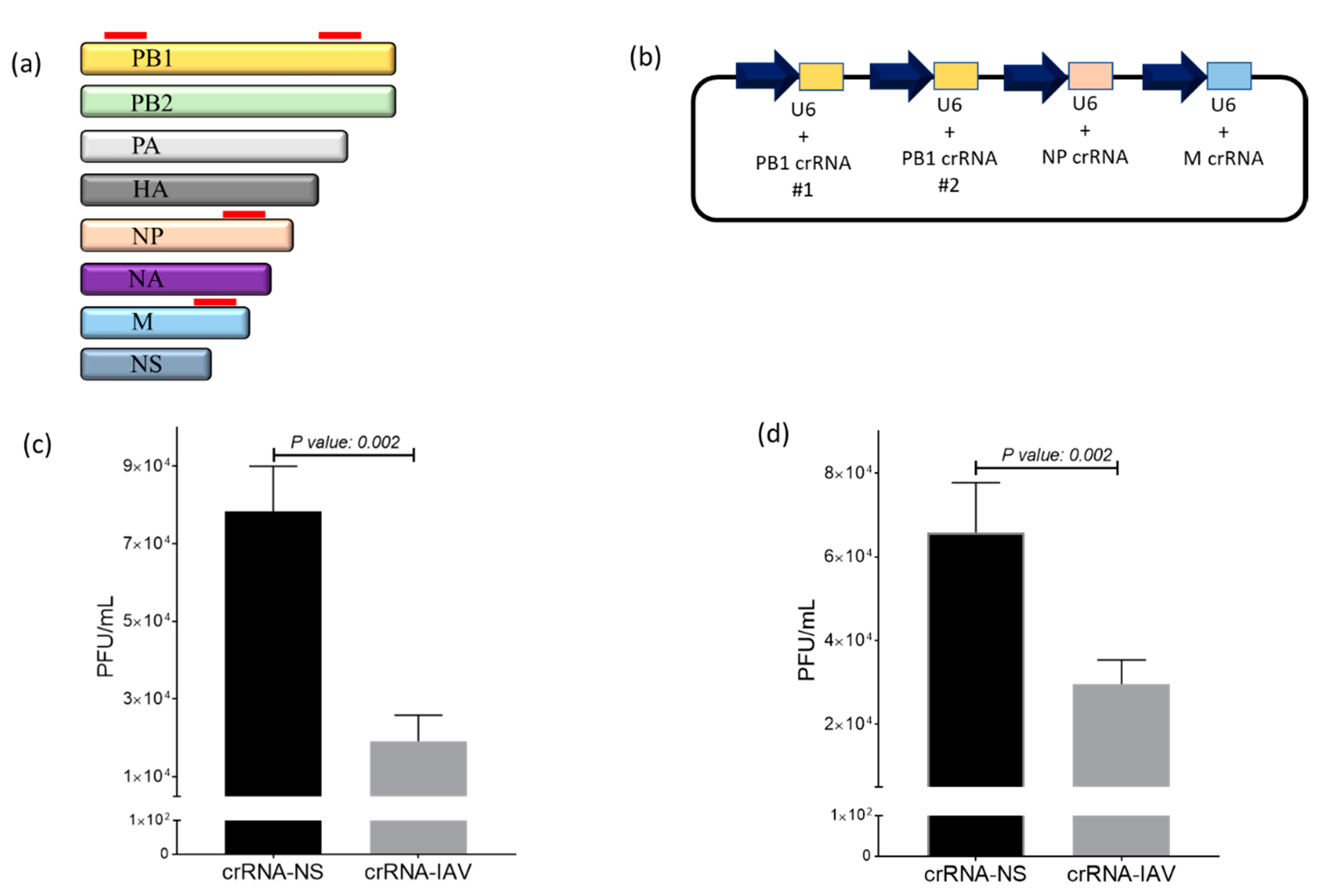
2.2. Design and Cloning of crRNA into the Expression Vector
2.3. Cell Culture and Transfections
2.4. DsRed Knockdown Assays
2.5. IAV Infection
2.6. Plaque Assay
2.7. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petrova, V.N.; Russell, C.A. The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 2018, 16, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; McManus, M.T.; Nguyen, T.; Shen, C.H.; Sharp, P.A.; Eisen, H.N.; Chen, J. RNA interference of influenza virus production by directly targeting mRNA for degradation and indirectly inhibiting all viral RNA transcription. Proc. Natl. Acad. Sci. USA 2003, 100, 2718–2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinton, T.M.; Challagulla, A.; Stewart, C.R.; Guerrero-Sanchez, C.; Grusche, F.A.; Shi, S.; Bean, A.G.; Monaghan, P.; Gunatillake, P.A.; Thang, S.H.; et al. Inhibition of influenza virus in vivo by siRNA delivered using ABA triblock copolymer synthesized by reversible addition-fragmentation chain-transfer polymerization. Nanomedicine 2014, 9, 1141–1154. [Google Scholar] [CrossRef] [PubMed]
- Castanotto, D.; Rossi, J.J. The promises and pitfalls of RNA-interference-based therapeutics. Nature 2009, 457, 426–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR–Cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR-Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, D.B.T.; Gootenberg, J.S.; Abudayyeh, O.O.; Franklin, B.; Kellner, M.J.; Joung, J.; Zhang, F. RNA editing with CRISPR-Cas13. Science 2017, 358, 1019–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.X.; Chong, S.; Zhang, H.; Makarova, K.S.; Koonin, E.V.; Cheng, D.R.; Scott, D.A. Cas13d Is a Compact RNA-Targeting Type VI CRISPR Effector Positively Modulated by a WYL-Domain-Containing Accessory Protein. Mol. Cell 2018, 70, 327–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyack, S.G.; Jenkins, K.A.; O’Neil, T.E.; Wise, T.G.; Morris, K.R.; Bruce, M.P.; McLeod, S.; Wade, A.J.; McKay, J.; Moore, R.J.; et al. A new method for producing transgenic birds via direct in vivo transfection of primordial germ cells. Transgenic Res. 2013, 22, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boë, F.; Masson, J.M. Selective cloning of a defined number of tandem DNA repeats in Escherichia coli. Nucleic Acids Res. 1996, 24, 2450–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challagulla, A.; Jenkins, K.A.; O’Neil, T.E.; Morris, K.R.; Wise, T.G.; Tizard, M.L.; Bean, A.G.; Schat, K.A.; Doran, T.J. Germline engineering of the chicken genome using CRISPR/Cas9 by in vivo transfection of PGCs. Anim. Biotechnol. 2020, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Izzard, L.; Ye, S.; Jenkins, K.; Xia, Y.; Tizard, M.; Stambas, J. miRNA modulation of SOCS1 using an influenza A virus delivery system. J. Gen. Virol. 2014, 95, 1880–1885. [Google Scholar] [CrossRef] [Green Version]
- Bawage, S.S.; Tiwari, P.M.; Santangelo, P.J. Synthetic mRNA expressed Cas13a mitigates RNA virus infections. bioRxiv 2018, 370460. [Google Scholar] [CrossRef] [Green Version]
- Brake, O.T.; Hooft, K.T.; Liu, Y.P.; Centlivre, M.; von Eije, K.J.; Berkhout, B. Lentiviral Vector Design for Multiple shRNA Expression and Durable HIV-1 Inhibition. Mol. Ther. 2008, 16, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Techakriengkrai, N.; Nedumpun, T.; Suradhat, S. Abrogation of PRRSV infectivity by CRISPR-Cas13b-mediated viral RNA cleavage in mammalian cells. Sci. Rep. 2020, 10, 9617. [Google Scholar] [CrossRef] [PubMed]
- Freije, C.A.; Myhrvold, C.; Boehm, C.K.; Lin, A.E.; Welch, N.L.; Carter, A.; Metsky, H.C.; Luo, C.Y.; Abudayyeh, O.O.; Gootenberg, J.S.; et al. Programmable Inhibition and Detection of RNA Viruses Using Cas13. Mol. Cell 2019, 76, 826–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challagulla, A.; Jenkins, K.A.; O’Neil, T.E.; Shi, S.; Morris, K.R.; Wise, T.G.; Paradkar, P.N.; Tizard, M.L.; Doran, T.J.; Schat, K.A. In Vivo Inhibition of Marek’s Disease Virus in Transgenic Chickens Expressing Cas9 and gRNA against ICP4. Microorganisms 2021, 9, 164. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target | Sequence (5′-3′) |
|---|---|
| DsRed | CGACATCCCCGACTACAAGAAGCTGTCC |
| PB1-#1 | TACACCATGGATACTGTCAACAGGACAC |
| PB2-#2 | CTGAGATCATGAAGATCTGTTCCACCAT |
| NP1 | AATGAAGGATCTTATTTCTTCGGAGACA |
| M | GAACACCGATCTTGAGGTTCTCATGGAA |
| NS control | ATGCATGCATGCATGCATGCATGCATGC |
| Primer | Sequence (5′-3′) |
|---|---|
| crRNA-DsRed-F | AAACGGACAGCTTCTTGTAGTCGGGGATGTCG |
| crRNA-DsRed-R | AAAACGACATCCCCGACTACAAGAAGCTGTCC |
| crRNA-PB1#1-F | AAACGTGTCCTGTTGACAGTATCCATGGTGTA |
| crRNA-PB1#1-R | AAAATACACCATGGATACTGTCAACAGGACAC |
| crRNA-PB1#2-F | AAACATGGTGGAACAGATCTTCATGATCTCAG |
| crRNA-PB1#2-R | AAAACTGAGATCATGAAGATCTGTTCCACCAT |
| crRNA-NP-F | AAACTGTCTCCGAAGAAATAAGATCCTTCATT |
| crRNA-NP-R | AAAAAATGAAGGATCTTATTTCTTCGGAGACA |
| crRNA-M-F | AAACTTCCATGAGAACCTCAAGATCGGTGTTC |
| crRNA-M-R | AAAAGAACACCGATCTTGAGGTTCTCATGGAA |
| crRNA-NS-F | AAACATGCATGCATGCATGCATGCATGCATGC |
| crRNA-NS-R | AAAAGCATGCATGCATGCATGCATGCATGCAT |
| TD1022 | ACTACCGGTGTCGACCGCCAGAGGGCCTATTTCCCA |
| TD1023 | ACTACCGGTCTCGAGGTGAATTCGAGCTCGGTACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Challagulla, A.; Schat, K.A.; Doran, T.J. In Vitro Inhibition of Influenza Virus Using CRISPR/Cas13a in Chicken Cells. Methods Protoc. 2021, 4, 40. https://doi.org/10.3390/mps4020040
Challagulla A, Schat KA, Doran TJ. In Vitro Inhibition of Influenza Virus Using CRISPR/Cas13a in Chicken Cells. Methods and Protocols. 2021; 4(2):40. https://doi.org/10.3390/mps4020040
Chicago/Turabian StyleChallagulla, Arjun, Karel A. Schat, and Timothy J. Doran. 2021. "In Vitro Inhibition of Influenza Virus Using CRISPR/Cas13a in Chicken Cells" Methods and Protocols 4, no. 2: 40. https://doi.org/10.3390/mps4020040
APA StyleChallagulla, A., Schat, K. A., & Doran, T. J. (2021). In Vitro Inhibition of Influenza Virus Using CRISPR/Cas13a in Chicken Cells. Methods and Protocols, 4(2), 40. https://doi.org/10.3390/mps4020040