
Simplified Method for Agrobacterium-Mediated Genetic Transformation of Populus x berolinensis K. Koch



Abstract
:1. Introduction
2. Materials and Methods
2.1. Nutrient Media Composition
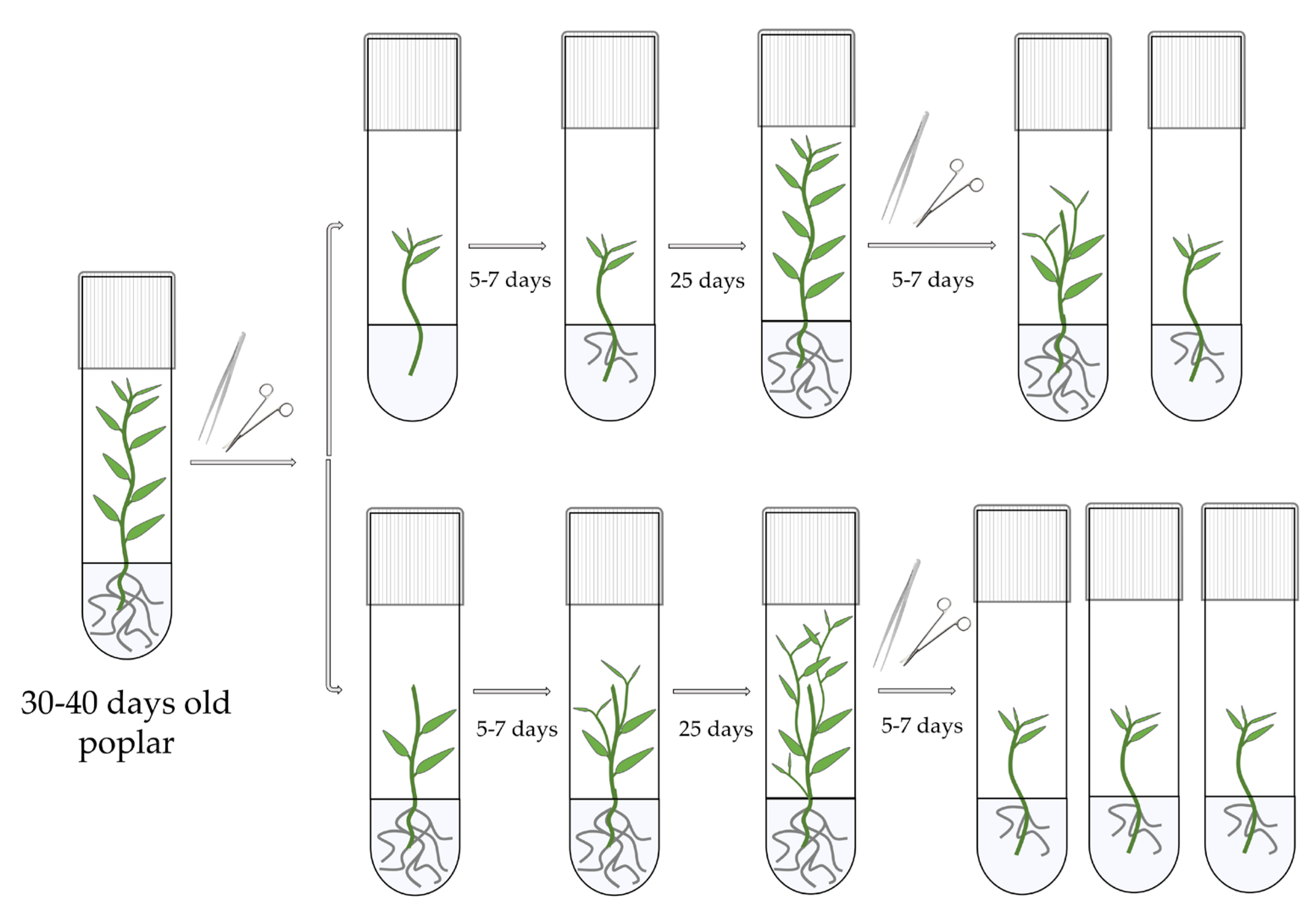
2.2. Micropropagation Technique and Growing Conditions
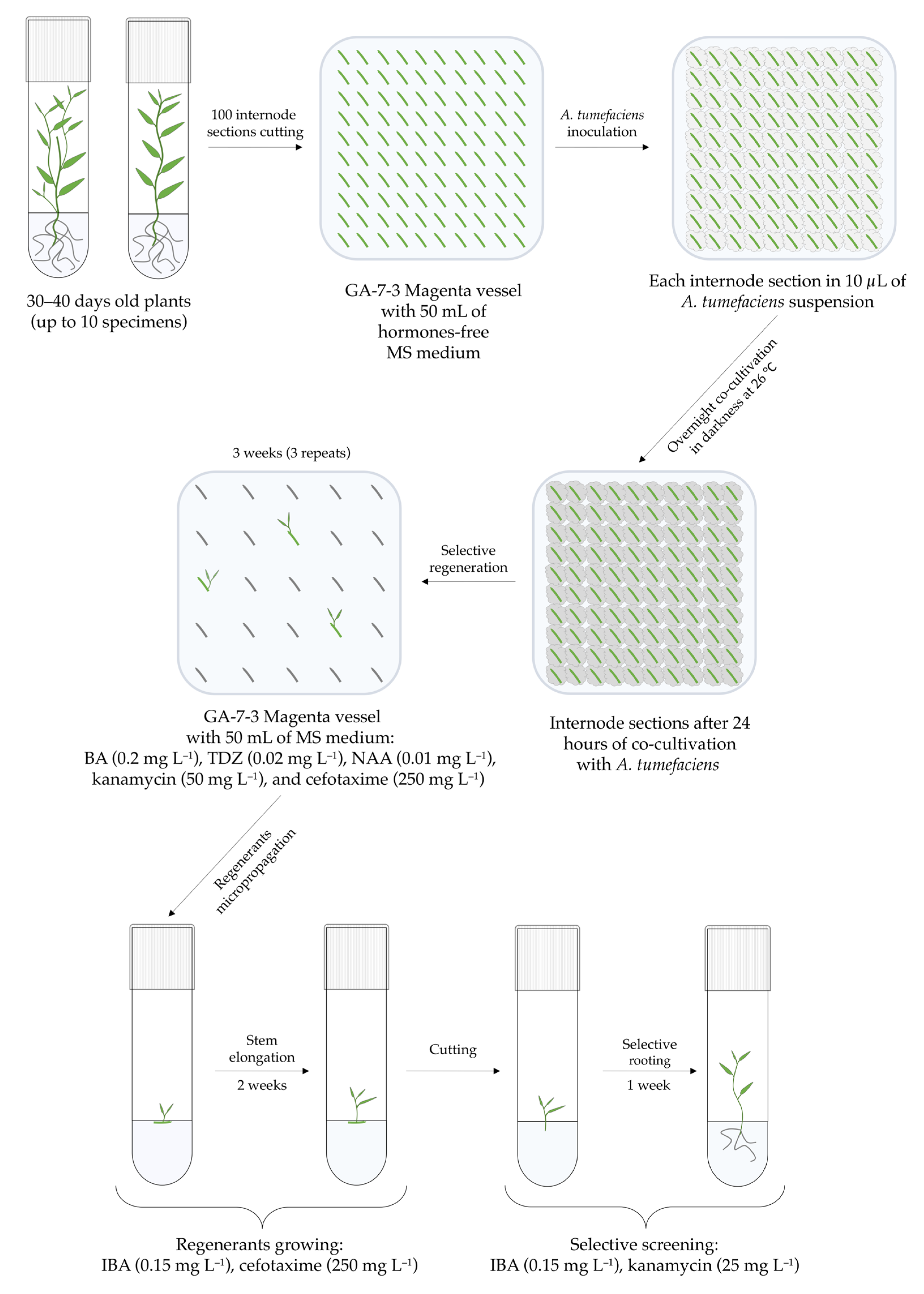
2.3. Agrobacterium-Mediated Transformation Protocol
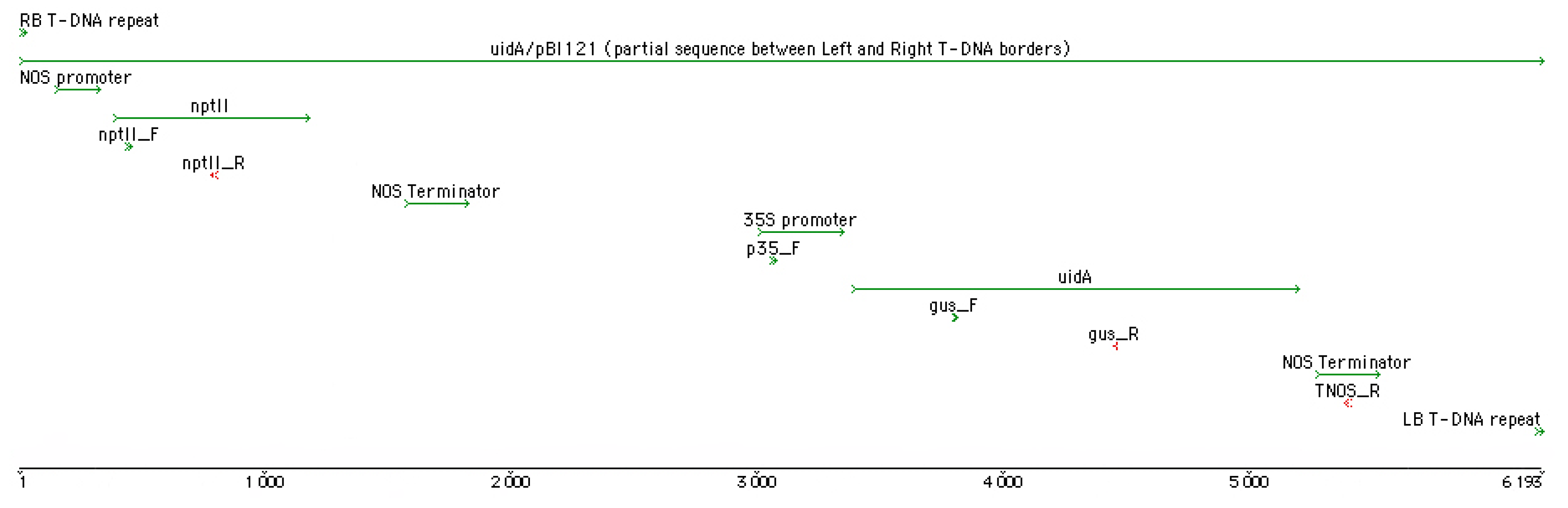
2.4. Total DNA Isolation, PCR, and Sequencing
2.5. GUS Staining Assay
3. Results
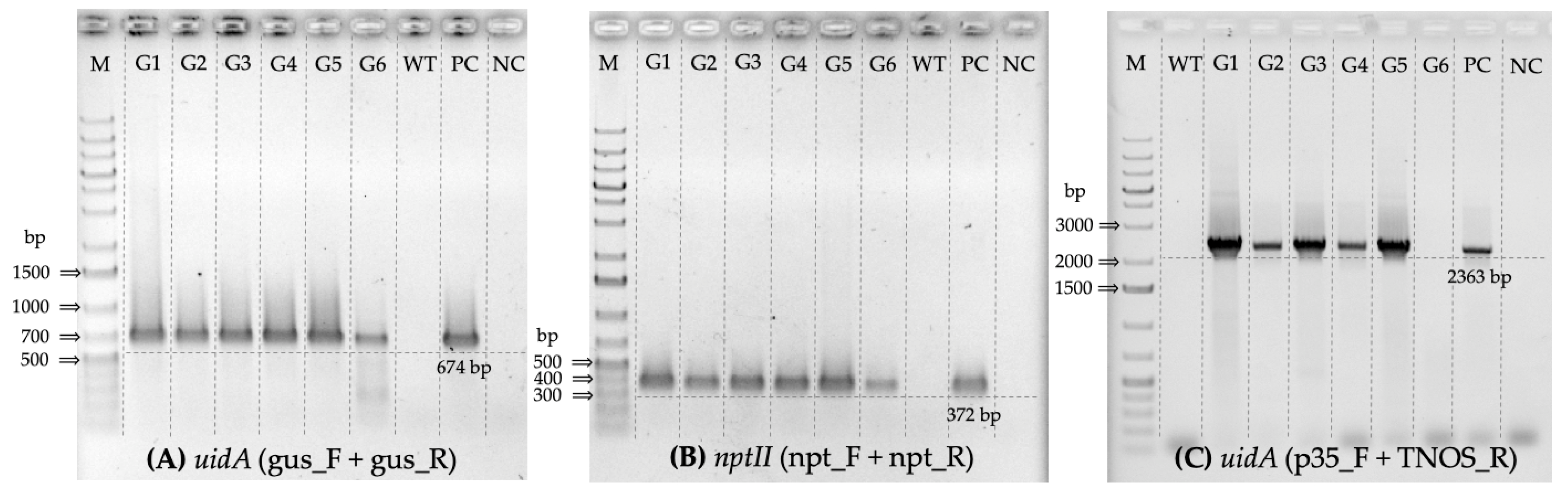
3.1. PCR-Based Screening of Kanamycin-Selected Regenerated Poplar Lines
3.2. Verification of nptII Expression in Transgenic Poplar Lines
3.3. Verification of uidA Expression in Transgenic Poplar Lines
4. Discussion
5. Conclusions
- Solid medium co-cultivation step. Explants were co-cultivated in 10 µL droplets of Agrobacterium suspension directly on the surface of a solid agar-based medium without subsequent rinsing or sterile paper drying after inoculation.
- Nutrient medium. A only one regeneration MS-based medium supplemented with BA (0.2 mg·L−1), TDZ (0.02 mg·L−1) and NAA (0.01 mg·L−1) was used.
- Explant type and direct morphogenesis. In vitro internode sections without axillary buds were used as explants for direct morphogenesis.
- Acetosyringone absence. Acetosyringone was not used as an induction agent for the vir genes during the genetic transformation.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parsons, T.J.; Sinkar, V.P.; Stettler, R.F.; Nester, E.W.; Gordon, M.P. Transformation of Poplar by Agrobacterium tumefaciens. Nat. Biotechnol. 1986, 4, 533–536. [Google Scholar] [CrossRef]
- Song, J.; Lu, S.; Chen, Z.-Z.; Lourenco, R.; Chiang, V.L. Genetic Transformation of Populus trichocarpa Genotype Nisqually-1: A Functional Genomic Tool for Woody Plants. Plant Cell Physiol. 2006, 47, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Han, B. Plant Regeneration and Agrobacterium-Mediated Transformation of Vacuolar H+-ATPase c Subunit Gene in Hybrid Poplar Populus davidiana Dode × P. bollena Lauche. BIO Web Conf. 2017, 8, 03004. [Google Scholar] [CrossRef]
- Maheshwari, P.; Kovalchuk, I. Agrobacterium-Mediated Stable Genetic Transformation of Populus angustifolia and Populus balsamifera. Front. Plant Sci. 2016, 7, 296. [Google Scholar] [CrossRef] [PubMed]
- Coleman, G.D.; Ernst, S.G. In Vitro Shoot Regeneration of Populus deltoides: Effect of Cytokinin and Genotype. Plant Cell Rep. 1989, 8, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Leple, J.C.; Brasileiro, A.C.M.; Michel, M.F.; Delmotte, F.; Jouanin, L. Transgenic Poplars: Expression of Chimeric Genes Using Four Different Constructs. Plant Cell Rep. 1992, 11, 1387–1141. [Google Scholar] [CrossRef] [PubMed]
- John, E.; Maqbool, A.; Malik, K.A. Optimization of Agrobacterium tumefaciens Mediated Transformation in Populus deltoides. Pak. J. Bot. 2014, 46, 1079–1086. [Google Scholar]
- Li, S.; Zhen, C.; Xü, W.; Wang, C.; Cheng, Y. Simple, Rapid and Efficient Transformation of Genotype Nisqually-1: A Basic Tool for the First Sequenced Model Tree. Sci. Rep. 2017, 7, 2638. [Google Scholar] [CrossRef]
- Wen, S.-S.; Ge, X.-L.; Wang, R.; Yang, H.-F.; Bai, Y.-E.; Guo, Y.-H.; Zhang, J.; Lu, M.-Z.; Zhao, S.-T.; Wang, L.-Q. An Efficient Agrobacterium-Mediated Transformation Method for Hybrid Poplar 84K (Populus alba × P. glandulosa) Using Calli as Explants. Int. J. Mol. Sci. 2022, 23, 2216. [Google Scholar] [CrossRef]
- Jensen, C.S.; Tzfira, T.; Vainstein, A.; Altman, A. Direct Regeneration and Selection of Populus tremula L. Transgenic Shoots from Agrobacterium tumefaciens-Transformed Stem Explants. In Biology of Root Formation and Development; Chu, E.H.Y., Hollaender, A., Eds.; International Society for Horticultural Science: Leuven, Belgium, 1997; pp. 209–211. [Google Scholar]
- Pavlichenko, V.V.; Protopopova, M.V.; Voinikov, V.K. A Comparative Study of Various Light Source Influences on the Plants Regenerative Potential Using Populus berolinensis Root Explants as an Example. IOP Conf. Ser. 2020, 548, 062093. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Wang, C.-K.; Soong, S.-C.; To, K.-Y. Complete Sequence of the Binary Vector PBI121 and Its Application in Cloning T-DNA Insertion from Transgenic Plants. Mol. Breed. 2003, 11, 287–293. [Google Scholar] [CrossRef]
- Estopà, M.; Marfà, V.; Melé, E.; Messeguer, J. Study of Different Antibiotic Combinations for Use in the Elimination of Agrobacterium with Kanamycin Selection in Carnation. Plant Cell Tissue Organ Cult. 2001, 65, 211–220. [Google Scholar] [CrossRef]
- Siti Suhaila, A.R.; Saleh, N.M. Inhibitory Effect of Kanamycin on In Vitro Culture of Lycopersicon esculentum Mill Cv. Mt11. J. Agrobiotechnol. 2010, 1, 79–86. [Google Scholar]
- Hull, G.A.; Devic, M. The β-Glucuronidase (Gus) Reporter Gene System Gene: Fusions; Spectrophotometric, Fluorometric, and Histochemical Detection. Plant Gene Transf. Expr. Protoc. 1995, 49, 125–141. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Protopopova, M.V.; Pavlichenko, V.V. Eranthis Salisb. (Ranunculaceae) in South Siberia: Insights into Phylogeography and Taxonomy. Diversity 2022, 14, 779. [Google Scholar] [CrossRef]
- Tzfira, T.; Jensen, C.S.; Wang, W.; Zuker, A.; Vinocur, B.; Altman, A.; Vainstein, A. Transgenic Populus tremula: A Step-By-Step Protocol for Its Agrobacterium-Mediated Transformation. Plant Mol. Biol. Rep. 1997, 15, 219–235. [Google Scholar] [CrossRef]
- Meilan, R.; Ma, C. Poplar (Populus Spp.). Methods Mol. Biol. 2006, 344, 143–151. [Google Scholar] [CrossRef]
- Cseke, L.J.; Cseke, S.B.; Podila, G.K. High Efficiency Poplar Transformation. Plant Cell Rep. 2007, 26, 1529–1538. [Google Scholar] [CrossRef]
- Yevtushenko, D.P.; Misra, S. Efficient Agrobacterium-Mediated Transformation of Commercial Hybrid Poplar Populus nigra L. × P. maximowiczii A. Henry. Plant Cell Rep. 2010, 29, 211–221. [Google Scholar] [CrossRef]
- Song, C.; Lu, L.; Guo, Y.; Xu, H.; Li, R. Efficient Agrobacterium-Mediated Transformation of the Commercial Hybrid Poplar Populus alba × Populus glandulosa Uyeki. Int. J. Mol. Sci. 2019, 20, 2594. [Google Scholar] [CrossRef]
- Nobakht Vakili, A.; Bagheri, H.; Azadi, P. Elimination of Macro Elements from Inoculation and Co-Cultivation Media Enhances the Efficiency of Agrobacterium-Mediated Transformation in Petunia. Physiol. Mol. Biol. Plants 2018, 24, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Giovanella, T.S.; Degenhardt, J.; Zanella, L.B.; Bespalhok, C. Organogenesis and Sonication-Assisted Agrobacterium-Mediated Transformation of Poplar Roots. Res. Sq. 2021. [CrossRef]
- Yücesan, B. Thidiazuron (TDZ): A Callus Minimizer for In Vitro Plant Production. In Thidiazuron: From Urea Derivative to Plant Growth Regulator; Ahmad, N., Faisal, M., Eds.; Springer: Singapore, 2018; pp. 289–295. [Google Scholar]
- Movahedi, A.; Zhang, J.; Amirian, R.; Zhuge, Q. An Efficient Agrobacterium-Mediated Transformation System for Poplar. Int. J. Mol. Sci. 2014, 15, 10780–10793. [Google Scholar] [CrossRef] [PubMed]
- Stachel, S.E.; Messens, E.; Montagu, M.V.; Zambryski, P. Identification of the Signal Molecules Produced by Wounded Plant Cells That Activate T-DNA Transfer in Agrobacterium tumefaciens. Nature 1985, 318, 624–629. [Google Scholar] [CrossRef]
- Asande, L.K.; Omwoyo, R.O.; Oduor, R.O.; Nyaboga, E.N. A Simple and Fast Agrobacterium-Mediated Transformation System for Passion Fruit KPF4 (Passiflora edulis f. edulis × Passiflora edulis f. flavicarpa). Plant Methods 2020, 16, 141. [Google Scholar] [CrossRef]
- Miguel, S.; Michel, C.; Biteau, F.; Hehn, A.; Bourgaud, F. In Vitro Plant Regeneration and Agrobacterium-Mediated Genetic Transformation of a Carnivorous Plant, Nepenthes mirabilis. Sci. Rep. 2020, 10, 17482. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, S.; Baghban Kohnehrouz, B. Efficient Agrobacterium-Mediated Transformation and Analysis of Transgenic Plants in Hybrid Black Poplar (Populus × Euromericana Dode Guinier). Plant Genet. Res. 2022, 8, 1–22. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Co-Cultivation | Regeneration (BTN-Medium) | Elongation | Rooting ** |
|---|---|---|---|---|
| MS5524 * | 4.3 g·L−1 | 4.3 g·L−1 | 4.3 g·L−1 | 4.3 g·L−1 |
| Thiamine | 1 mg·L−1 | 1 mg·L−1 | 1 mg·L−1 | 1 mg·L−1 |
| Pyridoxine | 0.5 mg·L−1 | 0.5 mg·L−1 | 0.5 mg·L−1 | 0.5 mg·L−1 |
| Nicotinic acid | 0.5 mg·L−1 | 0.5 mg·L−1 | 0.5 mg·L−1 | 0.5 mg·L−1 |
| Meso-Inositol | 50 mg·L−1 | 50 mg·L−1 | 50 mg·L−1 | 50 mg·L−1 |
| Sucrose | 3 g·L−1 | 3 g·L−1 | 2 g·L−1 | 2 g·L−1 |
| Agar | 7 g·L−1 | 7 g·L−1 | 7 g·L−1 | 7 g·L−1 |
| 6-Benzylaminopurine (BA) | – | 0.2 mg·L−1 | – | – |
| Thidiazuron (TDZ) | – | 0.02 mg·L−1 | – | – |
| 1-Naphthaleneacetic acid (NAA) | – | 0.01 mg·L−1 | – | – |
| Indole-3-butyric acid (IBA) | – | – | 0.15 mg·L−1 | 0.15 mg·L−1 |
| Kanamycin | – | 50 mg·L−1 | 50 mg·L−1 | 25 mg·L−1 |
| Cefotaxime | – | 250 mg·L−1 | 250 mg·L−1 | – |
| pH | 5.7 | 5.7 | 5.7 | 5.7 |
| Primer Name | Target | Sequence (5′ –> 3′) | Ta, °C | Amplicon, bp |
|---|---|---|---|---|
| npt_F | nptII | TGGAGAGGCTATTCGGCTATGA | 55 | 372 |
| npt_R | GATGTTTCGCTTGGTGGTCG | |||
| gus_F | uidA | CAACGAACTGAACTGGCAG | 55 | 674 |
| gus_R | AGAGGTTAAAGCCGACAGC | |||
| p35_F | 35S promoter (uidA) | CCATTGCCCAGCTATCTGTCACT | 55 | 2363 |
| TNOS_R | NOS terminator (uidA) | CCCATCTCATAAATAACGTCATGCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlichenko, V.V.; Protopopova, M.V. Simplified Method for Agrobacterium-Mediated Genetic Transformation of Populus x berolinensis K. Koch. Methods Protoc. 2024, 7, 12. https://doi.org/10.3390/mps7010012
Pavlichenko VV, Protopopova MV. Simplified Method for Agrobacterium-Mediated Genetic Transformation of Populus x berolinensis K. Koch. Methods and Protocols. 2024; 7(1):12. https://doi.org/10.3390/mps7010012
Chicago/Turabian StylePavlichenko, Vasiliy V., and Marina V. Protopopova. 2024. "Simplified Method for Agrobacterium-Mediated Genetic Transformation of Populus x berolinensis K. Koch" Methods and Protocols 7, no. 1: 12. https://doi.org/10.3390/mps7010012
APA StylePavlichenko, V. V., & Protopopova, M. V. (2024). Simplified Method for Agrobacterium-Mediated Genetic Transformation of Populus x berolinensis K. Koch. Methods and Protocols, 7(1), 12. https://doi.org/10.3390/mps7010012