1. Introduction
Pheromones are chemical cues released by animals and many other organisms that drive behavioral and/or physiological changes in conspecifics in adaptive manners [
1,
2,
3]. Living in aquatic environments that typically offer few visual cues, teleost fishes have commonly evolved to employ pheromones to mediate many aspects of their life histories [
4]. Sex pheromones are particularly important in the regulation of reproduction and can serve both priming (physiological) and/or releasing (behavioral) roles [
5,
6]. With only one possible exception [
7], all teleost sex pheromones described to date have been found to contain sex hormones and their derivatives [
5,
6]. Hormonal sex pheromones have now been identified in about half a dozen model fish species including two Eurasian Carps (Goldfish,
Carassius auratus, and Common Carp,
Cyprinus carpio), a few Cichlids including the tilapias,
Oreochromis sp., some gobies, and salmonids [
6,
8]. Being potent attractants, species-specific, and easily added to the water, sex pheromones are candidates for possible application as attractants for invasive fishes to facilitate their removal or quantification [
9].
Sex pheromones in teleost fishes are perhaps best understood in two Eurasian Carps, the Goldfish and its relative the Common Carp [
6,
10,
11]. Females of these seasonal scramble spawning Cyprinid carps [
10] employ a mixture of several sex steroid hormones and their metabolites as a priming pheromone to synchronize egg and sperm maturity during the perio-ovulatory period, and then a mixture of prostaglandin F
2α (PGF
2α) and its metabolites, 15keto-prostaglandin F
2α (15K-PGF
2α) and 13,14dihydro-15keto-PGF
2α (dh15K-PGF
2α) as a spawning (releasing) pheromone [
6,
10,
12]. PGF
2α is known to be produced by ovulated Goldfish as a hormone to stimulate female sexual behavior before functioning as a sex pheromone that is released within minutes of ovulation and spawning [
12,
13,
14]. The PGF pheromone both attracts males and signals that females are sexually receptive and carrying eggs, stimulating courtship. Both field and laboratory studies using Goldfish and Common Carp have shown that PGF-pheromone release can also be induced in immature fish by injecting them with PGF
2α or implanting them with osmotic pumps containing PGF
2α, which is then metabolized and released along with other natural products including two metabolites, 15K-PGF
2α and dh15K-PGF
2α [
12,
13]. Notably, PGF
2α-implants mimic natural circulating levels of PGF
2α, eliciting normal pheromone release for days and making this technique very useful as well as biologically relevant [
12]. Electrophysiological experiments have also shown that the olfactory systems of both the Goldfish and Common Carp and a variety of other fishes detect PGF
2α and its metabolites, 15K-PGF
2α and dh15K-PGF
2α, at low concentrations [
13,
15] in a sexually dimorphic manner that is inducible by androgen treatment [
16,
17,
18]. Studies of the PGF spawning pheromone show that Goldfish and Common Carp discern each other’s odors and that a variety of PGFs as well as other metabolites are responsible while specific ratios of PGFs may not be especially important [
11]. Finally, the activity of the PGF sex pheromone has been confirmed in the field using PGF
2α-implanted Common Carp odor [
12], suggesting this technique might be used as a tool to facilitate attracting, monitoring, and trapping these invasive fishes [
9].
Two congeneric species of carp from Asia, the Silver Carp (
Hypophthalmichthys molitrix) and the Bighead Carp (
H. nobilis), together known as Bigheaded Carps, and globally some of the most commonly cultured species [
19], were introduced in the United States in the late 1970s, escaped, and have become highly invasive in the Mississippi River [
20]. Although both of these Asian Carp species share many similarities with the Common Carp, they represent a distinctly different evolutionary branch that diverged several million years ago [
21,
22]. These naturally sympatric Asian Carps are now found in great abundance throughout most of the Mississippi River, and techniques to monitor their distribution and control them are sought. Both Bigheaded Carp species typically live in productive, turbid river waters and appear to have very well developed olfactory and internal gustatory systems [
23,
24] and reproduce episodically in a highly synchronized fashion in flowing turbid waters [
25,
26], suggestive of a pheromonally-mediated reproductive strategy. This suggests that they, like the Eurasian carps, employ hormonal sex pheromones, and PGFs in particular, to promote reproductive synchrony and that this trait could be exploited in their control. However, the possibility that Asian carps use F-series prostaglandins (PGFs) or other hormonal products as pheromones has not yet been examined.
The present study asked the question of whether the Silver Carp might also use PGFs as a sex pheromone to attract and discern conspecifics. Because this species naturally matures at a large size (about 60 cm) and is very difficult to study in the field, we conducted a proof-of-concept laboratory study with small (juvenile) fish using the same techniques that we have developed to artificially masculinize and study reproductive behaviors in the Goldfish as well as some other cyprinids [
17,
18]. We asked five questions. First, is the Silver Carp olfactory system acutely and specifically sensitive to PGFs and can it be increased by masculinization with androgens (as expected of a sex pheromone)? Second, are these individual PGFs highly attractive to masculinized fish? Third, when implanted with PGF
2α, do Silver Carp release a mixture of PGF
2α and its metabolites, and is this mixture profile distinctive, as might be expected if these PGFs function together as a mixture (blend)? Fourth, is the odor of PGF
2α-implanted Silver Carp and blend of PGFs they release attractive to male (masculinized) conspecifics and in a species-specific manner? Fifth, do the mixture profiles of PGFs released by the PGF
2α-implanted carps explain the behavioral specificity their pheromonal odor might have?
2. Results
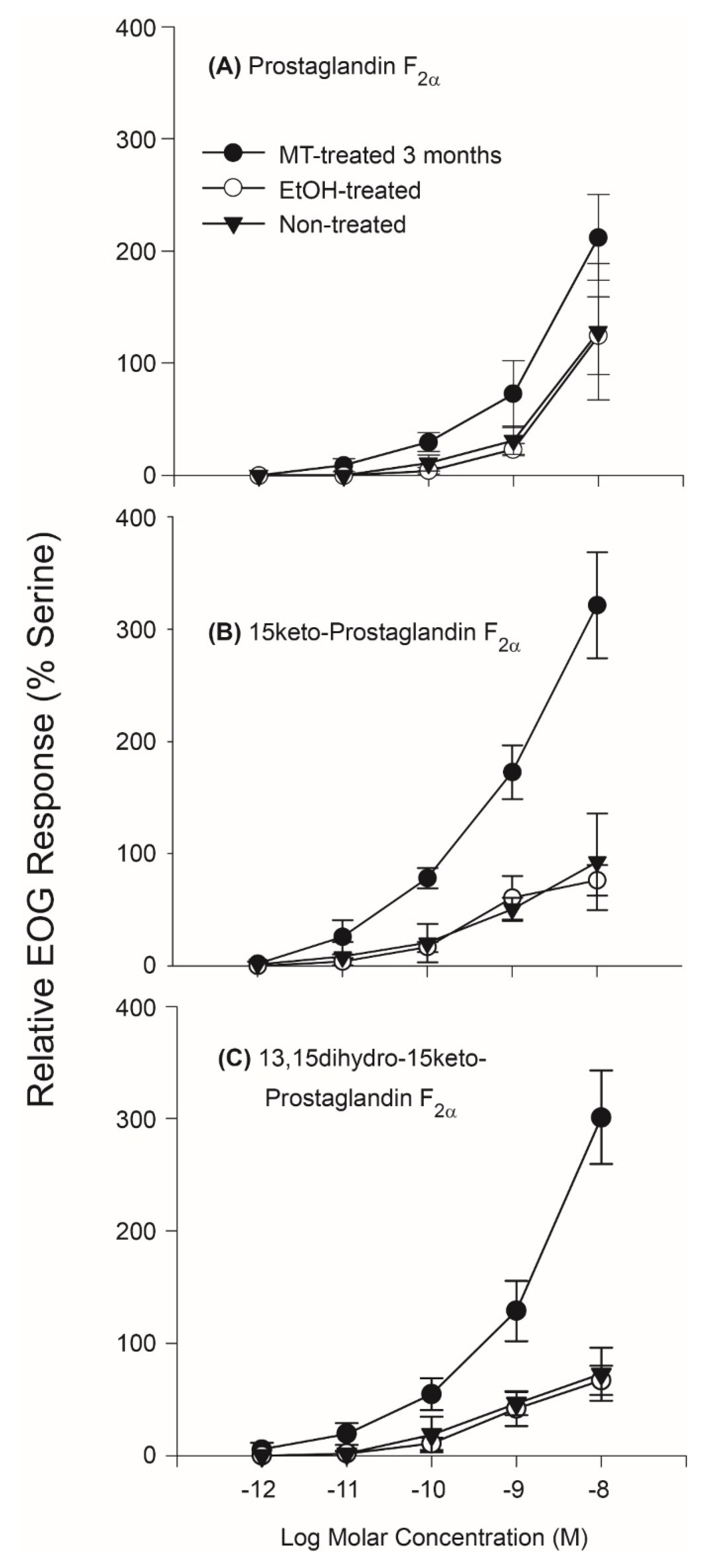
Electro-olfactogram (EOG) recording from the olfactory systems of Silver Carp established that they are highly and specifically sensitive to PGFs and that this sensitivity is influenced by androgens and thus likely sexually-dimorphic. Initial studies of the olfactory sensitivity of untreated juvenile Silver Carp established that while these fish are insensitive to E series prostaglandins (
Supplementary Data, Supplemental Figure S1), they detected all three F series prostaglandins previously identified as pheromones in the Goldfish: PGF
2α, 15K-PGF
2α, and dh15K-PGF
2α with high sensitivity. Further, while there was no apparent difference between the olfactory sensitivities of untreated juvenile fish and ethanol-treated control fishes to PGFs (their 95% confidence intervals overlapped), the olfactory sensitivity (EOG responses) of masculinized Silver Carp treated with methyltestosterone (MT) were greater than controls to 10
−8 Molar (M) PGF
2α (
p = 0.05; df = 2), 10
−8 M 15K-PGF
2α (
p = 0.02; df = 2), and 10
−8 M dh15K-PGF
2α (
p = 0.03, df = 2) (
Figure 1). Specifically, MT-treated fish detected PGF
2α down to a concentration of approximately 10
−11 M (i.e., their confidence intervals did not overlap the y axis) with responses rising to about twice that of the standard, 10
−5 M L-Serine (a food odor) at 10
−8 M, nearly twice the size response noted in the control carp to this concentration. Responses to 15K-PGF
2α were even larger; MT-treated carp detected this PGF down to approximately 10
−12 M and showed a response nearly three times that of the standard at 10
−8 M. Responses to dh15K-PGF
2α were similar to those elicited to 15K-PGF
2α: MT-treated carp detected it down to approximately 10
−12–10
−11 M and also responded to 10
−8 M with an EOG about three times the size of the standard. EOG tests of whether binary mixtures of PGFs are detected by more than one type of olfactory receptor suggested that PGF
2α is detected by its own independent olfactory receptor (the mean mixture discrimination index [MDI] for a mixture of PGF
2α and 15K-PGF
2α was 1.39 ± 0.7—different from 1.0 (
p = 0.02, df = 2), the expected MDI if they shared a receptor—and the MDI for PGF
2α and 13dh15K-PGF
2α was 1.34 ± 0.12 vs. 1.0 (
p = 0.05, df = 2). Conversely, it was not clear whether 15K-PGF
2α and 13dh15K-PGF
2α have their own receptors (the MDI for a mixture of PGF
2α and dh15K-PGF
2α was 1.12 ± 0.12 vs. 1.0;
p > 0.05, df = 2). When EOG responses to the PGFs were measured after a year of MT treatment and at the conclusion of behavioral experiments (see below), they were found to be about twice the size they were after 3 months of MT treatment (
Supplementary Data, Supplemental Figure S2).
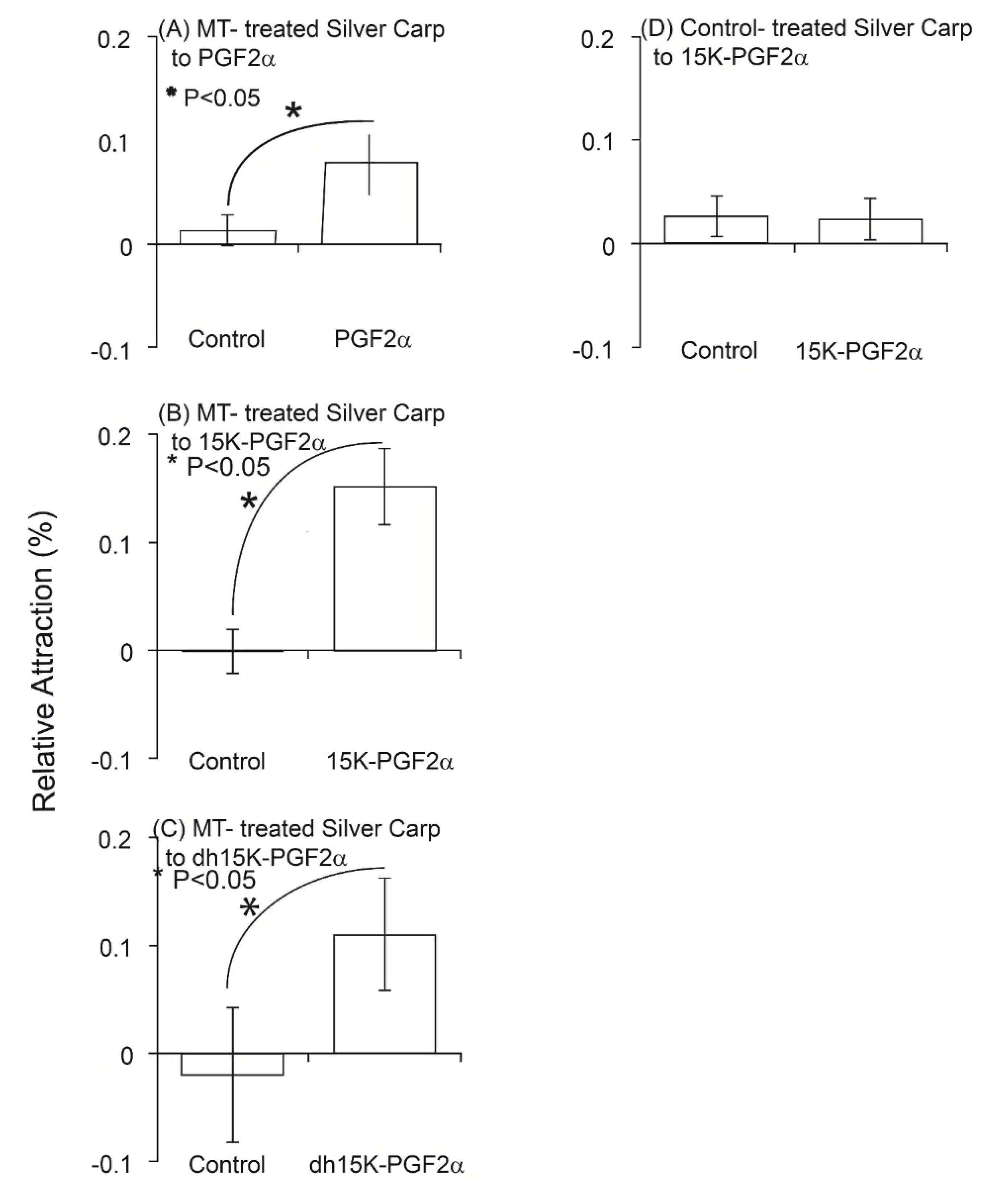
Tests of the behavioral activity individual PGFs in a circular maze showed that 10 nanomolar (nm) concentrations of each of the three PGFs attracted MT-treated Silver Carp (
p = 0.02,
p = 0.1,
p = 0.01, df = 9) when tested on their own and that control (EtOH-treated) Silver Carp were not attracted to 15K-PGF
2α (
p > 0.05; df = 10), further suggesting these compounds function as a sex pheromone(s) (
Figure 2). Nevertheless, these responses were very small (only about 10% of the fish were measurably attracted), suggesting that on their own, these individual PGFs might not be the entire pheromonal signal.
Further investigation of PGF
2α metabolism and release in Bigheaded carps to determine if mixtures of PGFs might instead function as the sex pheromone showed that they, like the Goldfish and Common Carp, release substantial but different quantities of all three PGFs (PGF
2α, 15K-PGF
2α, and dh15K-PGF
2α) to the water when implanted with PGF
2α (
Table 1). Notably, Silver Carp released approximately a third less PGF
2α than either the Bighead or Common Carp. Nevertheless, PGF
2α dominated all three mixtures with Silver Carp water containing more 15K-PGF
2α and dh15K-PGF
2α than the other two species’ release waters. Chi-square analysis showed that the quantities of the three PGFs varied between species (X
2 = 35.2;
p = 0.01), but that the relative amounts of the PGFs released by Common Carp and Bighead Carp did not differ from each other (X
2 = 0.426;
p > 0.05). When corrected for mass (molarity), the Silver Carp PGF release ratio (PGF
2α:15K-PGF
2α:dh15K-PGF
2α) was 94.7:2.3:2.7; Bighead Carp was 99.0:0.1:0.8; and Common Carp was 99.4:0.1:0.5 (after rounding).
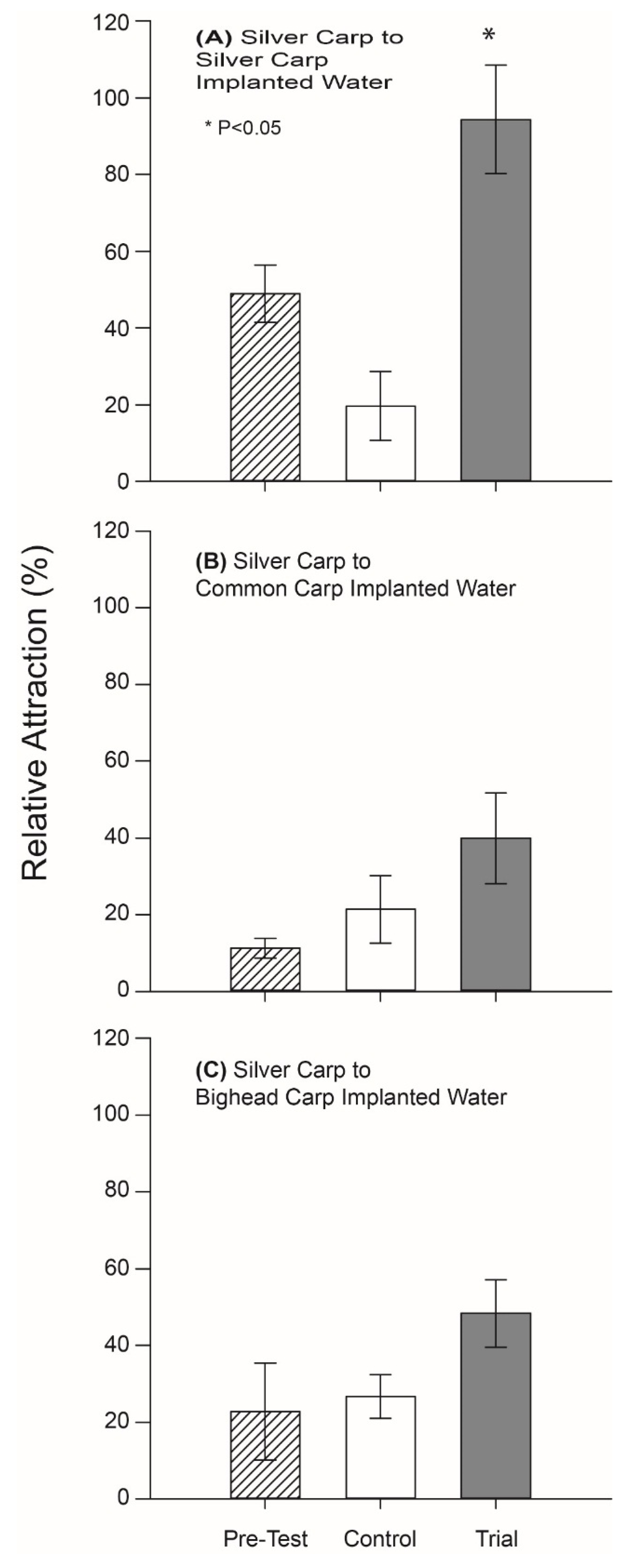
When the behavioral activity of PGF
2α-implanted carp odor was next tested in a rectangular maze designed to facilitate natural shoaling behavior using masculinized Silver Carp, we found that fish were strongly attracted (spending 97% of their time) to the holding water (odor) of PGF
2α-implanted conspecifics (
p = 0.03, df = 6) but ignored control well water (
Figure 3A). In contrast, Silver Carp did not appear to respond to either the odor of PGF
2α-implanted Bighead Carp odor, or the odor of PGF
2α-implanted Common Carp water (or well water controls) (
p > 0.05; df = 6;
Figure 3B,C).
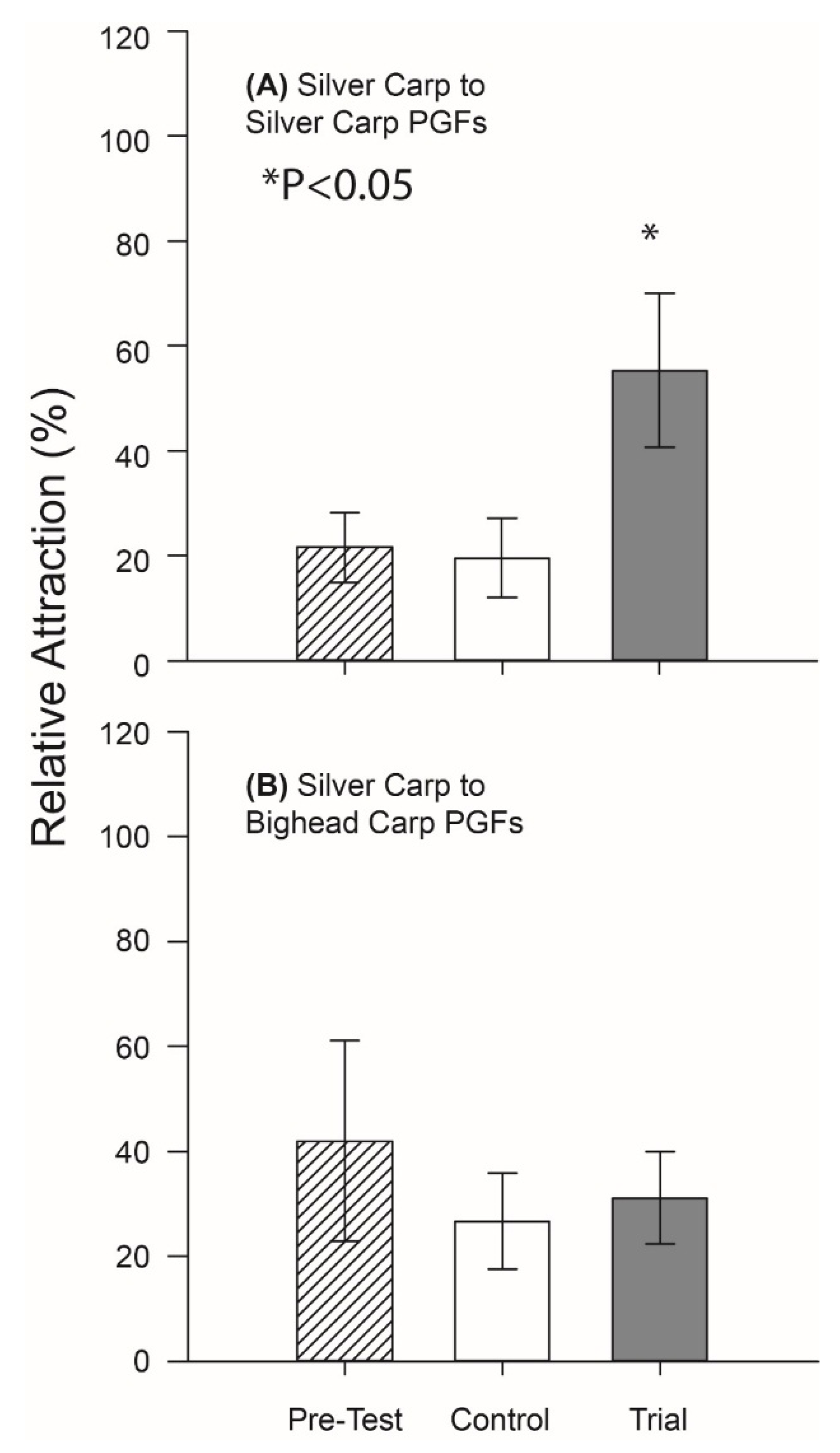
Finally, we confirmed that much of the behavioral activity of PGF
2α-implanted Silver Carp was associated with the blend of PGFs it contained; masculinized Silver Carp were strongly attracted to the mixture of PGFs found in the holding water of PGF
2α-implanted conspecifics (
p = 0.01, df = 6), but not the mixture of PGFs found in the holding water of implanted Bighead Carp (
Figure 4). Well water again had no apparent effect (
p > 0.05, df = 6).
3. Discussion
Using several well-established approaches, our proof-of concept laboratory study demonstrated that similar to the Eurasian carps, the Silver Carp likely uses a mixture of F prostaglandins as a sex pheromone which almost certainly is associated with spawning. In particular, we found that the olfactory systems of juvenile Silver Carp are remarkably sensitive to three F prostaglandins and that both this physiological and behavioral sensitivity is increased by androgen treatment. Further, at least two of the three PGFs are discriminated from each other. Additionally, masculinized fish were attracted to a mixture of the three PGFs, which we also showed to be released by conspecifics treated with PGF
2α following established techniques known to mimic natural pheromone release in Eurasian Carps [
11,
12]. Very likely, this sex pheromone is normally released in the urine by ovulated, sexually receptive Silver Carp females, as is the case for the Goldfish and Common Carp [
27], for which PGF
2α is also intimately linked to the process of ovulation and spawning behavior [
5,
14]. As with the Common Carp, the Silver Carp pheromone signal appears to be specific, although in the case of the Silver Carp the ratio (blend) of PGFs seems to be important. Nevertheless, because ours is a relatively small proof-of-concept laboratory study, this sex pheromone’s precise identity and function must now be confirmed in wild, naturally-ovulated free-ranging fish. This sex pheromone could be a very useful tool in Bigheaded Carp management and control as these fish are usually very difficult to locate and track in large flowing rivers [
9].
The Silver Carp joins approximately a dozen other fish species for which there is now evidence that PGF-based sex pheromones mediate reproductive behaviors, opening new doors for managing these invasive carp. It, however, is only one of a handful of species for which olfactory sensitivity, behavioral responsiveness, and PGF release have all been demonstrated and is the first Asian Carp species to be so described. Several apparent similarities and differences with Eurasian Carps are evident in this proof-of-concept study. First, while the overall olfactory sensitivities of the Bighead and Eurasian Carps to the three PGFs are similar, differences exist in the way they detect each of them. In particular, the concentration-response relationship to PGF
2α appears different between the Eurasian and the Asian (Bigheaded) species because, while it does not saturate in the former species, it does appear to in the latter species. Differences are also seen in responses to 15K-PGF
2α and dh15K-PGF
2α. Silver Carp respond to both of these PGFs in a very similar manner and possess a shared olfactory receptor(s) for them, while the Goldfish (the best studied example), responded to these PGFs in different ways. These observations both suggest that these species likely use PGF mixtures in slightly different ways and that the Silver Carp olfactory neurons may use different secondary messenger systems [
28]. This would be interesting and important to determine in the future with more definitive studies of receptor and species-specificity that test natural odors in naturally mature fishes. Pilot EOG studies show that the Bighead Carp also detects PGFs well (Lim and Sorensen, in preparation). Second, seemingly identical to the Goldfish, the Silver Carp olfactory system is quickly and specifically masculinized to PGFs using MT treatment and only these fish respond behaviorally, strongly suggesting that, as established in the Goldfish, PGFs are indeed sex pheromones [
13,
18]. Of course, it is important to confirm sexual dimorphism in wild-caught sexually mature male and female Silver Carps as has been done for Eurasian Carps because while MT treatment stimulates normal courtship (and presumably attraction), differences in other aspects of behavior could exist [
15,
18].
As well established in the Eurasian Carps [
11,
12,
13], PGF
2α-treatment clearly evokes the production of a species-specific PGF-based sex pheromone in the Silver Carp. However, unlike for the Common Carp [
11,
12], this blend seems to drive pheromonal activity in the Silver Carp. This may not be unreasonable because pheromonal blends are important in many sympatric species including many insects [
29,
30]. Notably, the Silver Carp seemingly evolved in sympatry with other carps including the Bighead Carp, while the Common Carp and Goldfish did not [
21], so Bigheaded Carps may have needed to evolve more precise species-isolation mechanisms to mediate reproductive behavior. The PGF pheromone is likely specifically associated with the act of spawning as in the Eurasian Carps. Because immature Bighead and Silver readily intermix [
31], common body odors probably do not serve a species-isolation function as in Eurasian carps [
30]. Sex pheromonally-based isolation mechanisms in Bigheaded Carps cannot be perfect given the observation of hybrid Silver-Bighead Carp in North America but not in China. Perhaps natural habitat features may help completely isolate these species when spawning in China, complimenting the actions of the pheromone [
32].
Our study has several notable strengths and a few weaknesses. Using well established electrophysiological and endocrinological techniques we clearly show for the first time that a relatively unstudied and important species, the Silver Carp, detects PGFs with high sensitivity using multiple receptors, and that this sensitivity is androgen dependent. Using two different behavioral tests, one of which has been used many times, we also show that androgen-treated carp respond to these PGFs, whose release we also show can be induced using established implant techniques that also stimulate release distinctive odors and mixtures of PGFs. The strongest behavioral responses were clearly seen to the blends of PGFs released by Silver Carp which also would have, been exposed to largely undiluted odors streams in the rectangular tank, suggesting a failure to detect 15K-PGF
2α in the Bigheaded Carp odor was unlikely to be the cause of its mixture’s lack of potency. Nevertheless, while these laboratory techniques and assays have been shown to mimic behaviors and release in a closely-related model cyprinid, the Goldfish, there is always a possibility Silver Carp may be different. Additionally, this study measured slightly different ratios of PGFs released by juvenile Common Carp than we saw in previous work using much larger individuals [
11], suggesting more study of PGF metabolism and release in fish is needed to fully understand the causes and significance of PGF metabolism. Accordingly, specific PGF ratios measured by small implanted laboratory fishes should be interpreted with caution. Whether or not 15K-PGF
2α and dh15K-PGF
2α play different roles in the Silver Carp pheromone must also be examined in greater detail, ideally with naturally mature and ovulated Silver Carp. Ideally larger, more natural test arenas or ponds might be used too and many more individuals than we were able to manage in this multiple-year laboratory-based study that was limited by ability to treat more than a few dozen fish with androgen for extended times. Nevertheless, given the fact that PGF
2α production, metabolism, and release is now known to be intimately associated with both ovulation and the expression of female sexual behavior (they increase within minutes of each other in the Goldfish [
18]), and the fact that the Goldfish has proven to be a powerful and reasonable model of many cyprinids [
5,
6], it seems highly likely that the Bigheaded Carps use also PGFs as female spawning pheromone as seen in many other cyprinids.
In conclusion, while our study, a relatively simple test of PGF sensitivity, release, and responses in the laboratory, cannot be considered definitive, when taken as a whole it appears to confirm that the Silver Carp and presumably its congener the Bighead Carp use PGF-based sex pheromones to attract each other and mediate spawning. These carps thus resemble dozens of other fish species, including the invasive Common Carp, in their use of this class of pheromones, opening new doors for invasive carp management. The seeming potency and specificity of their pheromone signal(s) are especially notable. A PGF-pheromone in the Silver Carp might have several uses. For example, given its high potency, it could be easily used as an attractant to bring males to specific locations for removal or to be counted [
9]. By including feeding attractants outside of the mating season [
33,
34,
35], an attractant strategy using different types of attractants including sex pheromones might be especially valuable [
9]. It seems especially notable that PGF
2α-implanted Silver Carps could be used an inexpensive platform to produce and release pheromones in wild fishes, as has been shown in the invasive Common Carp [
12]. Additionally, because hormonal PGF
2α is known to function as a female reproductive hormone that drives female behavior [
14], it creates the opportunity to use PGF
2α-implanted carps as Judas fish, which can lead managers to shoals of mature fish [
34,
35]. Lastly, recent studies have shown that PGF can be measured in waterways, making it possible to track PGFs together with eDNA to survey for Bigheaded Carps, while also identifying Carp gender and health [
9,
35]. Other studies now hint that Bigheaded Carps also use sex steroids as priming pheromones (Lim and Sorensen, in preparation) similar to the Goldfish [
5,
6,
10], suggesting a reliance on sex pheromones for many aspects of reproduction. We hope our study will serve as a foundation for future studies of pheromones in wild, free-ranging fish that will both examine and develop the basic science and application of sex pheromones in the Bigheaded Carps.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}