Seasonal Variation of Captive Meagre Acoustic Signalling: A Manual and Automatic Recognition Approach

Abstract
:1. Introduction
2. Results
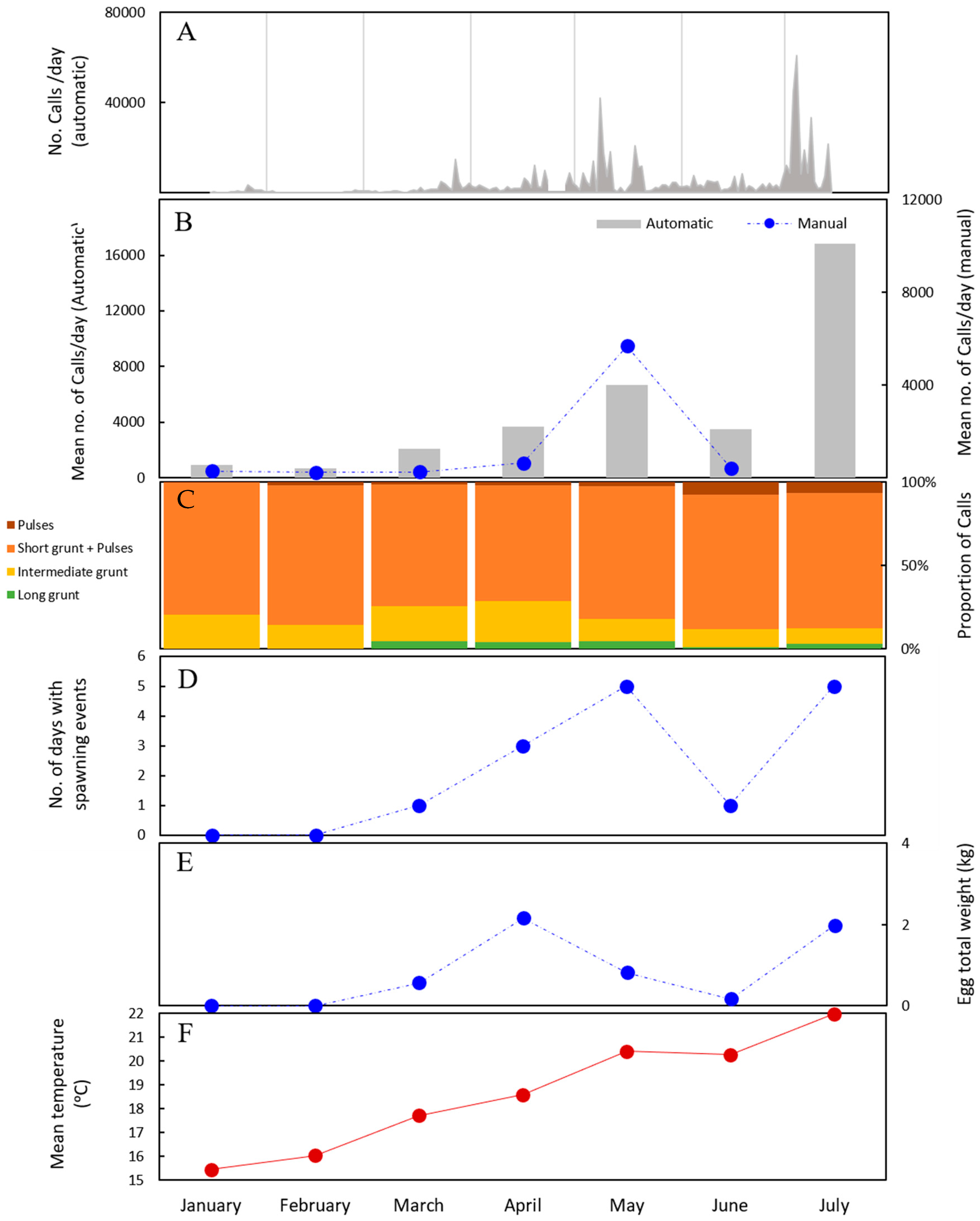
2.1. Validation of the Automatic Recognition System: Automatic Versus Manual Detection
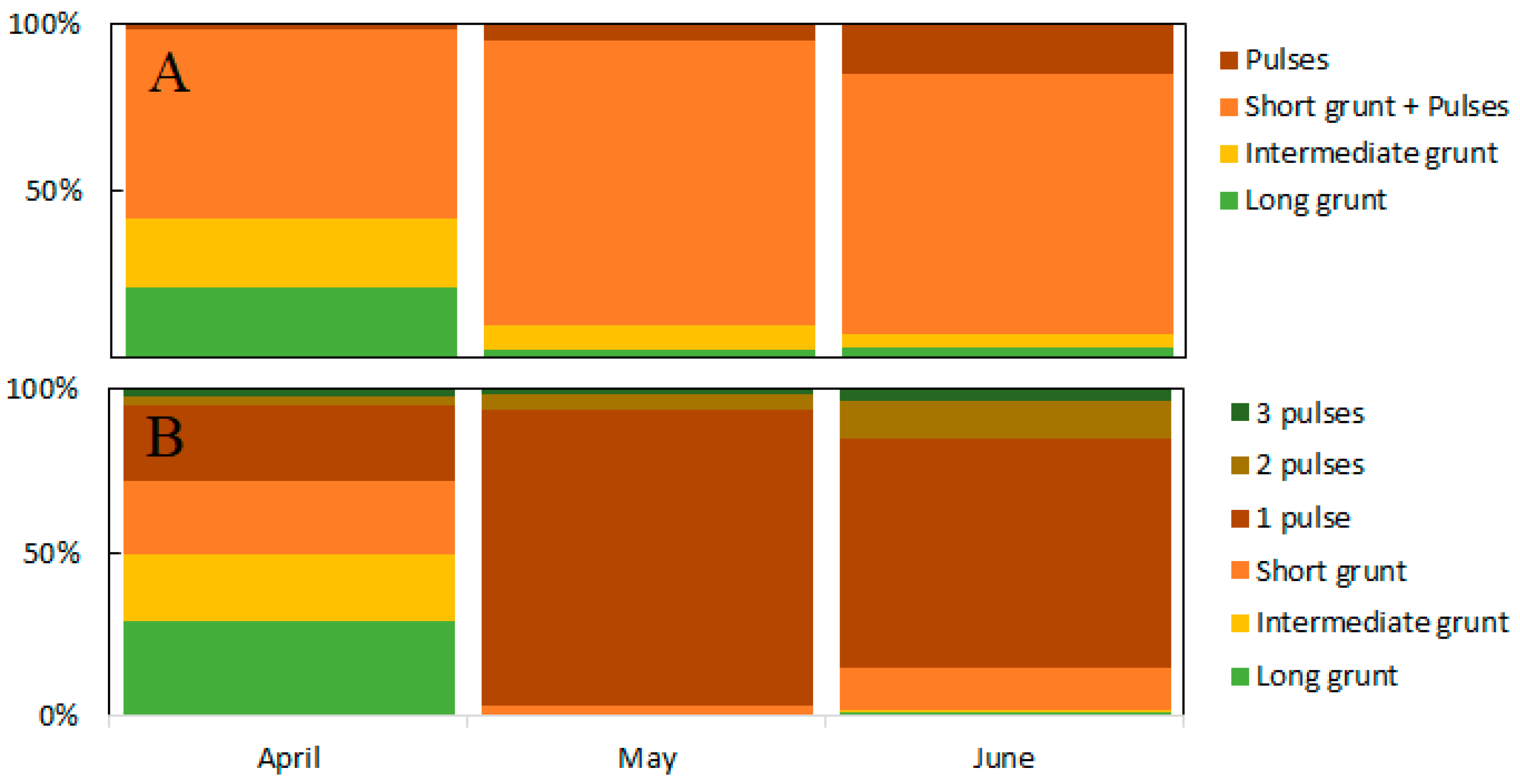
2.2. Seasonal Changes in Calling Rate
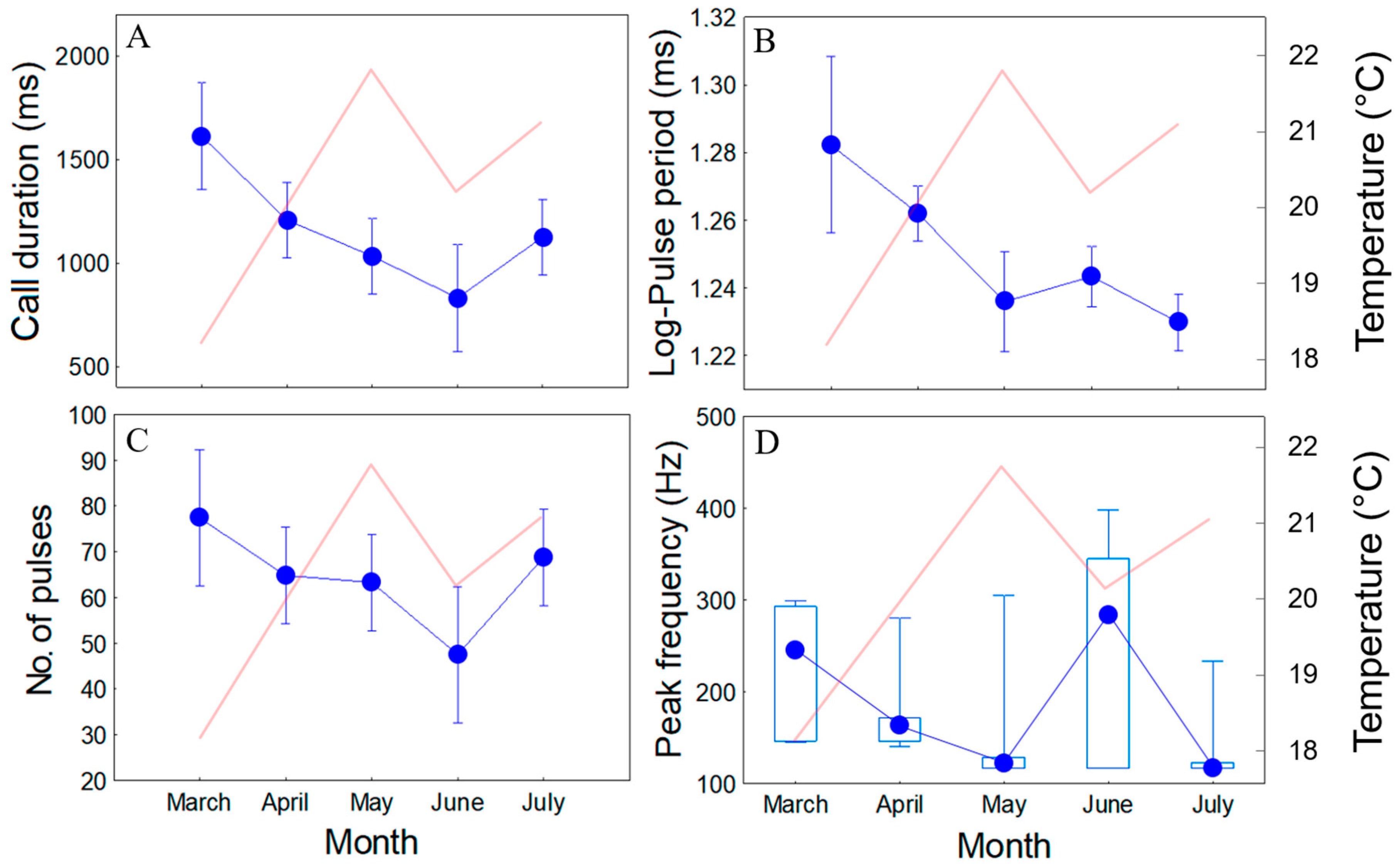
2.3. Seasonal Changes in Sound Features
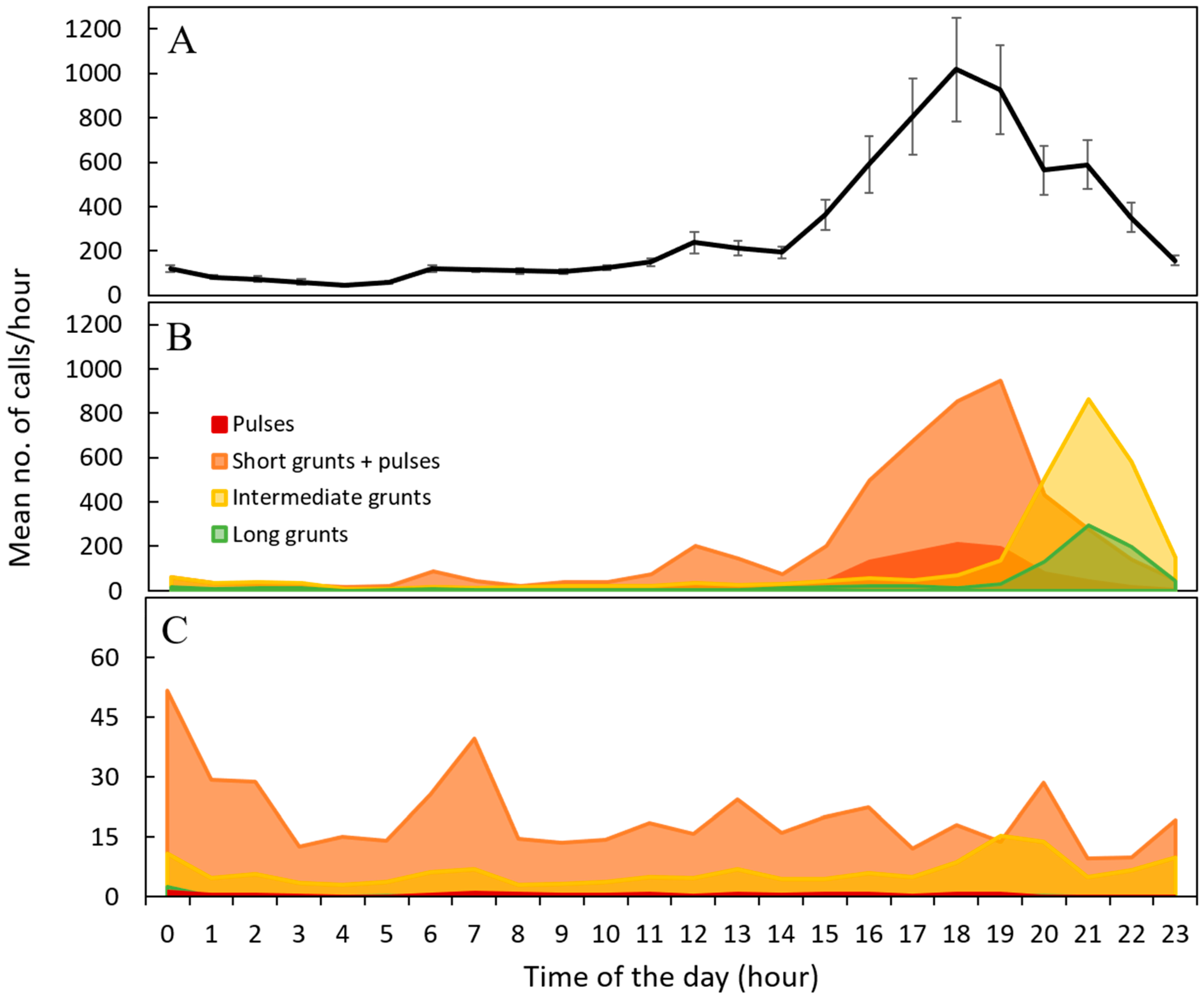
2.4. Diel Changes in Calling Rate
3. Discussion
4. Materials and Methods
4.1. Fish Maintenance
4.2. Data Collection
4.2.1. Acoustic Recordings
4.2.2. Detection of Spawning Events
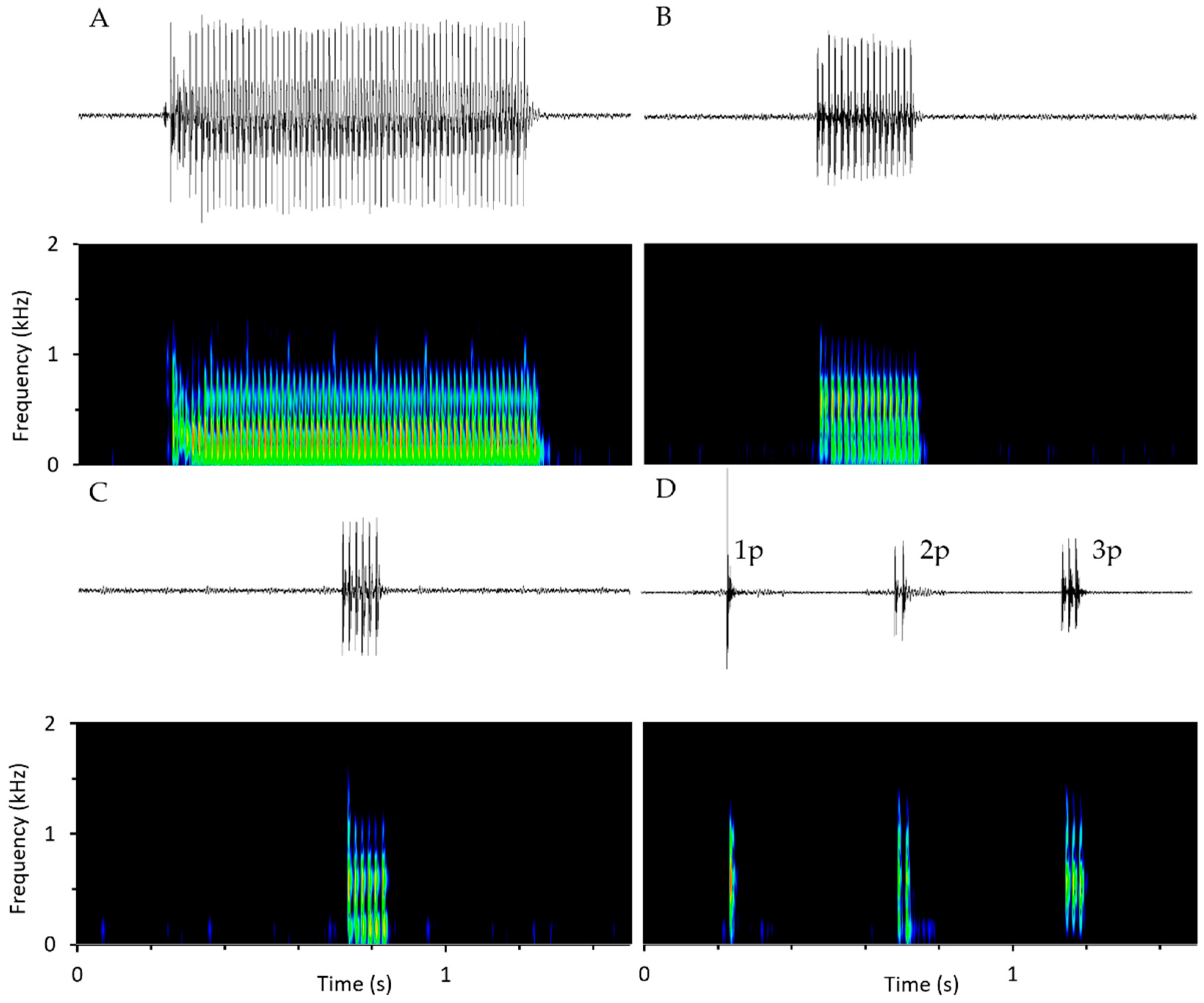
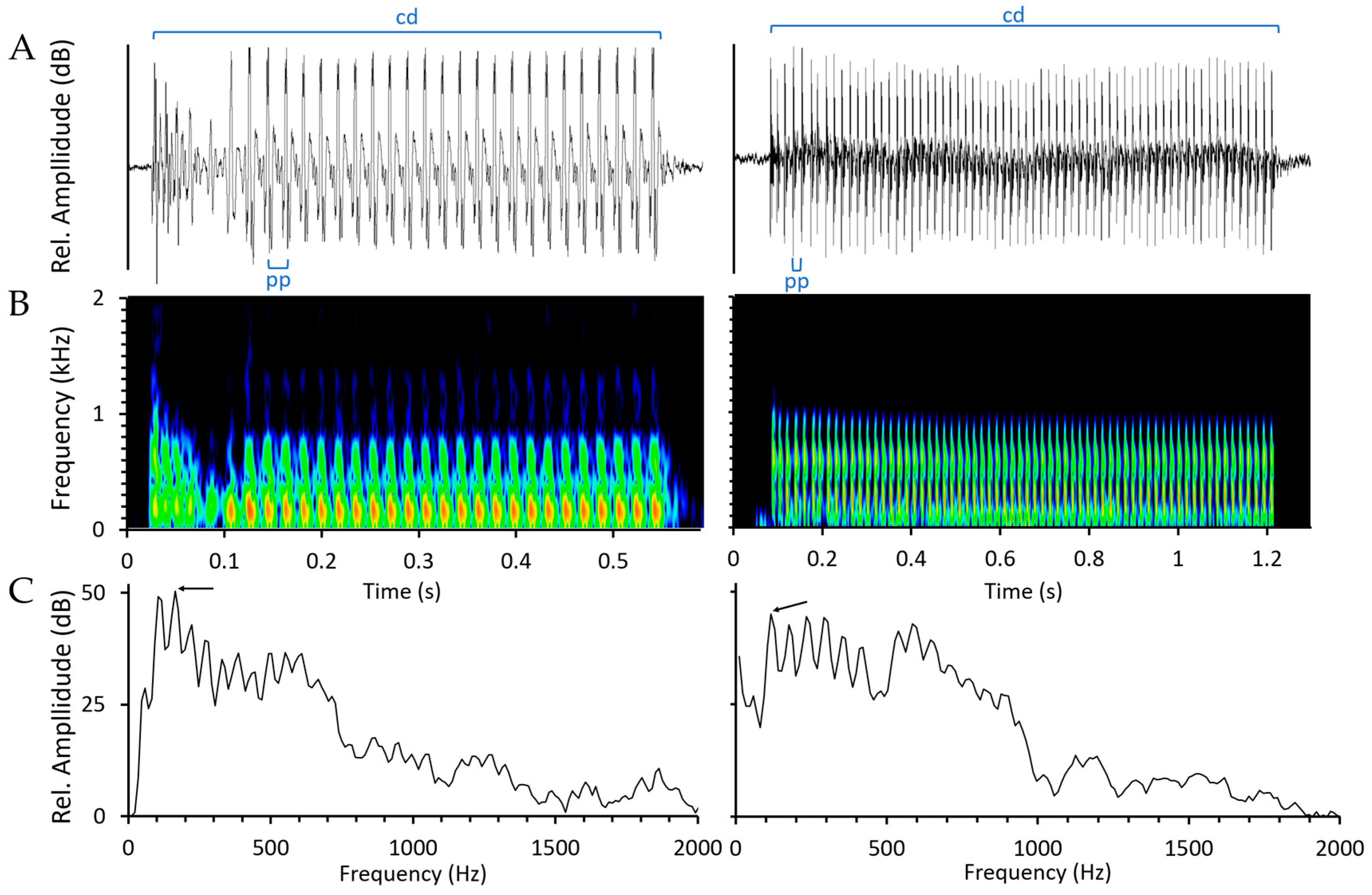
4.3. Manual Sound Detection, Classification and Feature Measurement
4.3.1. Sound Detection and Classification
4.3.2. Acoustic Features Measurements
4.4. Automatic Recognition
4.4.1. Signal Processing
4.4.2. The HMM Time Alignment Structure
4.4.3. System Training and Testing Process
4.4.4. Evaluation of the Recognition System
4.5. Analysis
4.5.1. Seasonal Changes in Calling Rate
4.5.2. Seasonal Changes in Calls Features
4.5.3. Diel Changes in Calling Rate
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Winn, H.E. The biological significance of fish sounds. Mar. Bio-Acoust. 1964, 2, 213–231. [Google Scholar]
- Connaughton, M.A.; Taylor, M.H. Seasonal and daily cycles in sound production associated with spawning in the weakfish, Cynoscion regalis. Environ. Biol. Fishes 1995, 42, 233–240. [Google Scholar] [CrossRef]
- McCauley, R.D. Biological Sea Noise in Northern Australia: Patterns of Fish Calling. Ph.D. Thesis, James Cook University, Douglas, Australia, 2001. [Google Scholar]
- Amorim, M.C.P. Diversity of sound production in fish. Commun. Fishes 2006, 1, 71–104. [Google Scholar]
- McWilliam, J.N.; Hawkins, A.D. A comparison of inshore marine soundscapes. J. Exp. Mar. Biol. Ecol. 2013, 446, 166–176. [Google Scholar] [CrossRef]
- Radford, C.A.; Jeffs, A.G.; Tindle, C.T.; Montgomery, J.C. Temporal patterns in ambient noise of biological origin from a shallow water temperate reef. Oecologia 2008, 156, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Radford, C.A.; Stanley, J.A.; Tindle, C.T.; Montgomery, J.C.; Jeffs, A.G. Localised coastal habitats have distinct underwater sound signatures. Mar. Ecol. Prog. Ser. 2010, 401, 21–29. [Google Scholar] [CrossRef]
- Hawkins, A.D. The use of passive acoustics to identify a haddock spawning area. In Proceedings of the International Workshop on the Applications of Passive Acoustics to Fisheries, Cambridge, MA, USA, 8–10 April 2002. [Google Scholar]
- Putland, R.L.; Ranjard, L.; Constantine, R.; Radford, C.A. A hidden Markov model approach to indicate Bryde’s whale acoustics. Ecol. Indic. 2018, 84, 479–487. [Google Scholar] [CrossRef]
- Miller, B.S.; Miller, E.J. The seasonal occupancy and diel behaviour of Antarctic sperm whales revealed by acoustic monitoring. Sci. Rep. 2018, 8, 5429. [Google Scholar] [CrossRef] [PubMed]
- Marques, T.A.; Thomas, L.; Martin, S.W.; Mellinger, D.K.; Ward, J.A.; Moretti, D.J.; Harris, D.; Tyack, P.L. Estimating animal population density using passive acoustics. Biol. Rev. Camb. Philos. Soc. 2013, 88, 287–309. [Google Scholar] [CrossRef] [PubMed]
- Putland, R.L.; Mackiewicz, A.G.; Mensinger, A.F. Localizing individual soniferous fish using passive acoustic monitoring. Ecol. Inform. 2018, 48, 60–68. [Google Scholar] [CrossRef]
- Parsons, M.J.; McCauley, R.D.; Mackie, M.C.; Siwabessy, P.; Duncan, A.J. Localization of individual mulloway (Argyrosomus japonicus) within a spawning aggregation and their behaviour throughout a diel spawning period. ICES J. Mar. Sci. 2009, 66, 1007–1014. [Google Scholar] [CrossRef]
- Parsons, M.J.G.; McCauley, R.D.; Mackie, M.C.; Duncan, A.J. A Comparison of techniques for ranging close-proximity mulloway (Argyrosomus japonicus) calls with a single hydrophone. Acoust. Aust. 2010, 38, 145–151. [Google Scholar]
- Vieira, M.; Fonseca, P.J.; Amorim, M.C.P.; Teixeira, C.J.C. Call recognition and individual identification of fish vocalizations based on automatic speech recognition: An example with the Lusitanian toadfish. J. Acoust. Soc. Am. 2015, 138, 3941–3950. [Google Scholar] [CrossRef]
- Clemins, P.J.; Johnson, M.T.; Leong, K.M.; Savage, A. Automatic classification and speaker identification of African elephant (Loxodonta africana) vocalizations. J. Acoust. Soc. Am. 2005, 117, 956. [Google Scholar] [CrossRef]
- Chou, C.H.; Lee, C.H.; Ni, H.W. Bird species recognition by comparing the HMMs of the syllables. In Proceedings of the Second International Conference on Innovative Computing, Informatio and Control (ICICIC 2007), Kumamoto, Japan, 5–7 September 2007; p. 143. [Google Scholar]
- Parsons, M.J.G.; McCauley, R.D.; Mackie, M.C. Spawning sounds of the mulloway (Argyrosomus japonicus). In Proceedings of the ACOUSTICS, Christchurch, New Zealand, 20–22 November 2006. [Google Scholar]
- Connaughton, M.A.; Taylor, M.H. Drumming, courtship, and spawning behavior in captive weakfish, Cynoscion regalis. Copeia 1996, 1996, 195–199. [Google Scholar] [CrossRef]
- Lagardère, J.P.; Mariani, A. Spawning sounds in meagre Argyrosomus regius recorded in the Gironde estuary, France. J. Fish Biol. 2006, 69, 1697–1708. [Google Scholar] [CrossRef]
- Montie, E.W.; Kehrer, C.; Yost, J.; Brenkert, K.; O’Donnell, T.; Denson, M.R. Long-term monitoring of captive red drum Sciaenops ocellatus reveals that calling incidence and structure correlate with egg deposition. J. Fish Biol. 2016, 88, 1776–1795. [Google Scholar] [CrossRef] [PubMed]
- Montie, E.W.; Hoover, M.; Kehrer, C.; Yost, J.; Brenkert, K.; O’Donnell, T.; Denson, M.R. Acoustic monitoring indicates a positive relationship between calling frequency and spawning in captive spotted seatrout (Cynoscion nebulosus). PeerJ 2017, 5, e2944. [Google Scholar] [CrossRef]
- Mok, H.-K.; Gilmore, R.G. Analysis of sound production in estuarine aggregations of Pogonias cromis, Bairdiella chrysoura, and Cynoscion nebulosus (Sciaenidae). Bull. Inst. Zool. Acad. Sin. 1983, 22, 157–186. [Google Scholar]
- Saucier, M.H.; Baltz, D.M. Spawning site selection by spotted seatrout, Cynoscion nebulosus, and black drum, Pogonias cromis, in Louisiana. Environ. Biol. Fishes 1993, 36, 257–272. [Google Scholar] [CrossRef]
- Luczkovich, J.J.; Daniel, H.J.; Sprague, M.W.; Johnson, S.E. Characterization of Critical Spawning Habitats of Weakfish, Spotted Seatrout and Red Drum In Pamlico Sound Using Hydrophone Surveys; North Carolina Department of Environment and Natural Resources: Morehead City, NC, USA, 1999. [Google Scholar]
- Quéméner, L.; Suquet, M.; Mero, D.; Gaignon, J.-L. Selection method of new candidates for finfish aquaculture: The case of the French Atlantic, the Channel and the North Sea coasts. Aquat. Living Resour. 2002, 15, 293–302. [Google Scholar] [CrossRef]
- Reynolds, D.; Rose, R. Robust text-independent speaker identification using Gaussian mixture speaker models. IEEE Trans. Speech Audio Proc. 1995, 3, 72–83. [Google Scholar] [CrossRef]
- Lippmann, R.P. An introduction to computing with neural nets. IEEE ASSP Mag. 1987, 4, 4–22. [Google Scholar] [CrossRef]
- Yu, H.; Oh, Y. A neural network for 500 vocabulary word spotting using acoustic sub-word units. In Proceedings of the 1997 IEEE International Conference on Acoustics, Speech, and Signal Processing, Munich, Germany, 21–24 April 1997; pp. 3277–3280. [Google Scholar]
- Baker, J. The DRAGON system—An overview. Acoust. Speech Signal Process. IEEE Trans. 1975, 23, 24–29. [Google Scholar] [CrossRef]
- Jelinek, F. Continuous speech recognition by statistical methods. Proc. IEEE 1976, 64, 532–556. [Google Scholar] [CrossRef]
- Jelinek, F.; Bahl, L.; Mercer, R. Design of a linguistic statistical decoder for the recognition of continuous speech. IEEE Trans. Inf. Theory 1975, 21, 250–256. [Google Scholar] [CrossRef]
- Rabiner, L.R. A tutorial on hidden Markov models and selected applications in speech recognition. Proc. IEEE 1989, 77, 257–286. [Google Scholar] [CrossRef]
- Young, S.; Bloothooft, G. Corpus-Based Methods in Language and Speech Processing; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997. [Google Scholar]
- Tellechea, J.S.; Fine, M.L.; Norbis, W. Passive acoustic monitoring, development of disturbance calls and differentiation of disturbance and advertisement calls in the A rgentine croaker U mbrina canosai (S ciaenidae). J. Fish Biol. 2017, 90, 1631–1643. [Google Scholar] [CrossRef]
- Sattar, F.; Cullis-Suzuki, S.; Jin, F. Identification of fish vocalizations from ocean acoustic data. Appl. Acoust. 2016, 110, 248–255. [Google Scholar] [CrossRef]
- Ibrahim, A.K.; Chérubin, L.M.; Zhuang, H.; Schärer Umpierre, M.T.; Dalgleish, F.; Erdol, N.; Ouyang, B.; Dalgleish, A. An approach for automatic classification of grouper vocalizations with passive acoustic monitoring. J. Acoust. Soc. Am. 2018, 143, 666–676. [Google Scholar] [CrossRef]
- Monczak, A.; Ji, Y.; Soueidan, J.; Montie, E.W. Automatic detection, classification, and quantification of sciaenid fish calls in an estuarine soundscape in the Southeast United States. PLoS ONE 2019, 14, e0209914. [Google Scholar] [CrossRef]
- Noda, J.; Travieso, C.; Sánchez-Rodríguez, D. Automatic taxonomic classification of fish based on their acoustic signals. Appl. Sci. 2016, 6, 443. [Google Scholar] [CrossRef]
- Lin, T.-H.; Tsao, Y.; Akamatsu, T. Comparison of passive acoustic soniferous fish monitoring with supervised and unsupervised approaches. J. Acoust. Soc. Am. 2018, 143, EL278–EL284. [Google Scholar] [CrossRef]
- Harakawa, R.; Ogawa, T.; Haseyama, M.; Akamatsu, T. Automatic detection of fish sounds based on multi-stage classification including logistic regression via adaptive feature weighting. J. Acoust. Soc. Am. 2018, 144, 2709–2718. [Google Scholar] [CrossRef] [PubMed]
- Malfante, M.; Mars, J.I.; Dalla Mura, M.; Gervaise, C. Automatic fish sounds classification. J. Acoust. Soc. Am. 2018, 143, 2834–2846. [Google Scholar] [CrossRef] [PubMed]
- Potter, J.; Mellinger, D.; Clark, C. Marine mammal call discrimination using artificial neural networks. J. Acoust. Soc. Am. 1994, 96, 1255. [Google Scholar] [CrossRef]
- Murray, S.; Mercado, E.; Roitblat, H. The neural network classification of false killer whale (Pseudorca crassidens) vocalizations. J. Acoust. Soc. Am. 1998, 104, 3626. [Google Scholar] [CrossRef]
- Schaar, M. van der Neural network-based sperm whale click classification. J. Mar. Biol. Assoc. UK 2007, 87, 35. [Google Scholar] [CrossRef]
- Pace, F.; White, P.; Adam, O. Hidden Markov Modeling for humpback whale (Megaptera Novaeanglie) call classification. Proc. Meetings Acoust. 2012, 17, 070046. [Google Scholar]
- Gillespie, D.; Caillat, M.; Gordon, J.; White, P. Automatic detection and classification of odontocete whistles. J. Acoust. Soc. Am. 2013, 134, 2427–2437. [Google Scholar] [CrossRef]
- Peso Parada, P.; Cardenal-López, A. Using Gaussian mixture models to detect and classify dolphin whistles and pulses. J. Acoust. Soc. Am. 2014, 135, 3371–3380. [Google Scholar] [CrossRef]
- Lin, T.-H.; Chou, L.-S. Automatic classification of delphinids based on the representative frequencies of whistles. J. Acoust. Soc. Am. 2015, 138, 1003–1011. [Google Scholar] [CrossRef]
- Erbs, F.; Elwen, S.H.; Gridley, T. Automatic classification of whistles from coastal dolphins of the southern African subregion. J. Acoust. Soc. Am. 2017, 141, 2489–2500. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Bu, L.; Wang, X.; Li, C.; Sun, Z.; Yan, H.; Hua, B.; Duan, F.; Yang, J. Clicks classification of sperm whale and long-finned pilot whale based on continuous wavelet transform and artificial neural network. Appl. Acoust. 2018, 141, 26–34. [Google Scholar] [CrossRef]
- Campbell, G.; Gisiner, R.; David, A.; Milette, L. Acoustic identification of female Steller sea lions (Eumetopias jubatus). J. Acoust. Soc. Am. 2002, 101, 2920–2928. [Google Scholar] [CrossRef]
- Guest, W.C.; Lasswell, J.L. A note on courtship behavior and sound production of red drum. Copeia 1978, 1978, 337–338. [Google Scholar] [CrossRef]
- Luczkovich, J.J.; Sprague, M.W.; Johnson, S.E.; Pullinger, R.C. Delimiting spawning areas of weakfish Cynoscion regalis (family Sciaenidae) in Pamlico Sound, North Carolina using passive hydroacoustic surveys. Bioacoustics 1999, 10, 143–160. [Google Scholar] [CrossRef]
- McIver, E.L.; Marchaterre, M.A.; Rice, A.N.; Bass, A.H. Novel underwater soundscape: Acoustic repertoire of plainfin midshipman fish. J. Exp. Biol. 2014, 217, 2377–2389. [Google Scholar] [CrossRef] [PubMed]
- Fish, J.F.; Cummings, W.C. A 50-dB increase in sustained ambient noise from fish (Cynoscion xanthulus). J. Acoust. Soc. Am. 1972, 52, 1266–1270. [Google Scholar] [CrossRef]
- Takemura, A.; Takita, T.; Mizue, K. Studies on the underwater sound, 7: Underwater calls of the Japanese marine drum fishes (Sciaenidae). Bull. Jpn. Soc. Sci. Fish. 1978, 54, 21–27. [Google Scholar]
- Parsons, M.J.G.; McCauley, R.D.; Mackie, M.C. Characterisation of mulloway argyrosomus japonicus advertisement sounds. Acoust. Aust. 2013, 41, 196–201. [Google Scholar]
- Borie, A.; Mok, H.-K.; Chao, N.L.; Fine, M.L. Spatiotemporal variability and sound characterization in Silver Croaker Plagioscion squamosissimus (Sciaenidae) in the Central Amazon. PLoS ONE 2014, 9, e99326. [Google Scholar] [CrossRef]
- Feher, J.J.; Waybright, T.D.; Fine, M.L. Comparison of sarcoplasmic reticulum capabilities in toadfish (Opsanus tau) sonic muscle and rat fast twitch muscle. J. Muscle Res. Cell Motil. 1998, 19, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Ladich, F. Acoustic communication in fishes: Temperature plays a role. Fish Fish. 2018, 19, 598–612. [Google Scholar] [CrossRef]
- Vicente, J.R.; Fonseca, P.J.; Amorim, M.C.P. Effects of temperature on sound production in the painted goby Pomatoschistus pictus. J. Exp. Mar. Biol. Ecol. 2015, 473, 1–6. [Google Scholar] [CrossRef]
- Bass, A.H.; Baker, R. Sexual dimorphisms in the vocal control system of a teleost fish: Morphology of physiologically identified neurons. J. Neurobiol. 1990, 21, 1155–1168. [Google Scholar] [CrossRef]
- Fine, M.L. Seasonal and geographical variation of the mating call of the oyster toadfish Opsanus tau L. Oecologia 1978, 36, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Torricelli, P.; Lugli, M.; Pavan, G. Analysis of sounds produced by male Padogobius martensi (Pisces, Gobiidae) and factors affecting their structural properties. Bioacoustics 1990, 2, 261–275. [Google Scholar] [CrossRef]
- Brantley, R.K.; Bass, A.H. Alternative male spawning tactics and acoustic signals in the plainfin midshipman fish Porichthys notatus Girard (Teleostei, Batrachoididae). Ethology 1994, 96, 213–232. [Google Scholar] [CrossRef]
- Connaughton, M.A.; Taylor, M.H.; Fine, M.L. Effects of fish size and temperature on weakfish disturbance calls: Implications for the mechanism of sound generation. J. Exp. Biol. 2000, 203, 1503–1512. [Google Scholar] [PubMed]
- Holt, S.A.; Holt, G.J. Effects of variable salinity on reproduction and early life stages of spotted seatrout. In Biology of the Spotted Seatrout; CRC Press: Boca Raton, FL, USA, 2002; pp. 140–150. [Google Scholar]
- Holt, G.J.; Holt, S.A.; Arnold, C.R. Diel periodicity of spawning in sciaenids. Mar. Ecol. Prog. Ser. 1985, 27, 7. [Google Scholar] [CrossRef]
- Montie, E.W.; Vega, S.; Powell, M. Seasonal and spatial patterns of fish sound production in the May River, South Carolina. Trans. Am. Fish. Soc. 2015, 144, 705–716. [Google Scholar] [CrossRef]
- Middaugh, D.P. Reproductive ecology and spawning periodicity of the Atlantic silverside, Menidia menidia (Pisces: Atherinidae). Copeia 1981, 766–776. [Google Scholar] [CrossRef]
- Doherty, P.J. Diel, lunar and seasonal rhythms in the reproduction of two tropical damselfishes: Pomacentrus flavicauda and P. wardi. Mar. Biol. 1983, 75, 215–224. [Google Scholar] [CrossRef]
- Hobson, E.S.; Chess, J.R. Trophic relationships among fishes and plankton in the lagoon at Enewetak Atoll, Marshall Island. Fish. Bull 1978, 76, 133–153. [Google Scholar]
- Lobel, P.S. Diel, lunar, and seasonal periodicity in the reproductive behavior of the pomacanthid fish, Centropyge potteri, and some other reef fishes in Hawaii. Pac. Sci. 1978, 32, 193–207. [Google Scholar]
- Ferraro, S.P. Daily time of spawning of 12 fishes in the Peconic Bays, New York, USA. Fish. Bull. US 1980, 78, 455–464. [Google Scholar]
- Robertson, D.R. On the spawning behavior and spawning cycles of eight surgeonfishes (Acanthuridae) from the Indo-Pacific. Environ. Biol. Fishes 1983, 9, 193–223. [Google Scholar] [CrossRef]
- Amorim, M.C.P.; Hawkins, A.D. Growling for food: Acoustic emissions during competitive feeding of the streaked gurnard. J. Fish Biol. 2000, 57, 895–907. [Google Scholar] [CrossRef]
- Fonseca, P.J.; Maia Alves, J. Electret Capsule Hydrophone: A New Underwater Sound Detector. Patent Application PT105, 2011. [Google Scholar]
- O’shaughnessy, D. Speech Communication: Human and Machine; Addison-Wesley: Boston, MA, USA, 1987. [Google Scholar]
- McDermott, E.; Iwamida, H.; Katagiri, S.; Tohkura, Y. Shift-tolerant LVQ and hybrid LVQ-HMM for phoneme recognition. In Readings in Speech Recognition; Morgan Kaufmann Publishers: San Mateo, CA, USA, 1990; pp. 425–438. [Google Scholar]
- Baum, L.; Petrie, T.; Soules, G.; Weiss, N. A maximization technique occurring in the statistical analysis of probabilistic functions of Markov chains. Ann. Math. Stat. 1970, 41, 164–171. [Google Scholar] [CrossRef]
- Forney, G. The viterbi algorithm. Proc. IEEE 1973, 6, 268–278. [Google Scholar] [CrossRef]
- Young, S.; Evermann, G.; Gales, M. The HTK Book Version 3.4; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Zar, J.H. Biostatistical Analysis Englewood Cliffs; Prentice-Hall: New York, NY, USA, 1984; p. 360. [Google Scholar]
- Amorim, M.C.P.; Vasconcelos, R.O.; Marques, J.F.; Almada, F. Seasonal variation of sound production in the Lusitanian toadfish Halobatrachus didactylus. J. Fish Biol. 2006, 69, 1892–1899. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Objective | System | Feature | Reference | |
|---|---|---|---|---|---|
| Fish | Meagre | C | HMM | MFCC a | Present study |
| Lusitanian toadfish | I,C | HMM | MFCC, LPC, a | [15] | |
| Plainfin midshipman | C | SVM | MRAF | [36] | |
| 4 grouper species | S | KNN, SVM, Sparse | (W)MFCC, (W)MRAF | [37] | |
| 4 sciaenid species | S | f | SPL | [38] | |
| On-line fishes sound database | S | SVM | MFCC, LFCC, SE, SL | [39] | |
| Unknown fishes | Fish sound detection | PC-NMF, GMM | [40] | ||
| SVM, KNN, NLR | MFCC,a | [41] | |||
| C | RF, SVM | MFCC,a | [42] | ||
| Mammals | Cetaceans | S | ANN | SCF | [43] |
| C | ANN | b | [44] | ||
| C | ANN | c | [45] | ||
| C | HMM | LPCC, MFCC | [46] | ||
| S | g | [47] | |||
| C,S | GMM | a, i | [48] | ||
| S | h | a | [49] | ||
| C | HMM | [9] | |||
| S | g | [50] | |||
| S | ANN | e | [51] | ||
| Sea lions | I | ANN | d | [52] |
| Predicted Group Membership | |||||||
|---|---|---|---|---|---|---|---|
| Sound type | LG | IG | SG | Pulses | False negative | Correct identification rate (%) | |
| Long grunt (LG) | 39 | 108 | 0 | 0 | 1 | 26.4 | 95.0 |
| Intermediate grunt (IG) | 1 | 345 | 20 | 0 | 4 | 93.2 | |
| Short grunt (SG) | 0 | 20 | 1485 | 12 | 30 | 96.0 | 76.8 |
| Pulses | 0 | 0 | 1568 | 175 | 915 | 6.6 | |
| False positive | 0 | 3 | 121 | 7 | |||
| Overall mean | 43.3 % | 78.8 % | |||||
| Sound Parameters | March | April | May | June | July | |
|---|---|---|---|---|---|---|
| Call duration (ms) | Mean ± SD | 1613 ± 376 | 1208 ± 405 | 1034 ± 436 | 831 ± 227 | 1127 ± 468 |
| Range | 1112–2230 | 524–1835 | 449–2176 | 517–1348 | 646–2480 | |
| No. of pulses | Mean ± SD | 77 ± 18 | 65 ± 21 | 63 ± 26 | 48 ± 13 | 69 ± 30 |
| Range | 54–109 | 30–95 | 30–130 | 30–78 | 39–160 | |
| Pulse period (ms) | Mean ± SD | 21 ± 1 | 19 ± 1 | 16 ± 1 | 18 ± 0.3 | 17 ± 0.4 |
| Range | 21–23 | 18–20 | 15–17 | 17–18 | 15–17 | |
| Peak frequency (Hz) | Mean ± SD | 230 ± 62 | 200 ± 100 | 151 ± 64 | 258 ± 104 | 124 ± 26 |
| Range | 146–299 | 141–563 | 117–305 | 117–398 | 117–234 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, M.; Pereira, B.P.; Pousão-Ferreira, P.; Fonseca, P.J.; Amorim, M.C.P. Seasonal Variation of Captive Meagre Acoustic Signalling: A Manual and Automatic Recognition Approach. Fishes 2019, 4, 28. https://doi.org/10.3390/fishes4020028
Vieira M, Pereira BP, Pousão-Ferreira P, Fonseca PJ, Amorim MCP. Seasonal Variation of Captive Meagre Acoustic Signalling: A Manual and Automatic Recognition Approach. Fishes. 2019; 4(2):28. https://doi.org/10.3390/fishes4020028
Chicago/Turabian StyleVieira, Manuel, Beatriz P. Pereira, Pedro Pousão-Ferreira, Paulo J. Fonseca, and M. Clara P. Amorim. 2019. "Seasonal Variation of Captive Meagre Acoustic Signalling: A Manual and Automatic Recognition Approach" Fishes 4, no. 2: 28. https://doi.org/10.3390/fishes4020028
APA StyleVieira, M., Pereira, B. P., Pousão-Ferreira, P., Fonseca, P. J., & Amorim, M. C. P. (2019). Seasonal Variation of Captive Meagre Acoustic Signalling: A Manual and Automatic Recognition Approach. Fishes, 4(2), 28. https://doi.org/10.3390/fishes4020028