Behavioural Response of Juvenile Common Carp (Cyprinus carpio) and Juvenile Channel Catfish (Ictalurus punctatus) to Strobe Light


Abstract
:1. Introduction
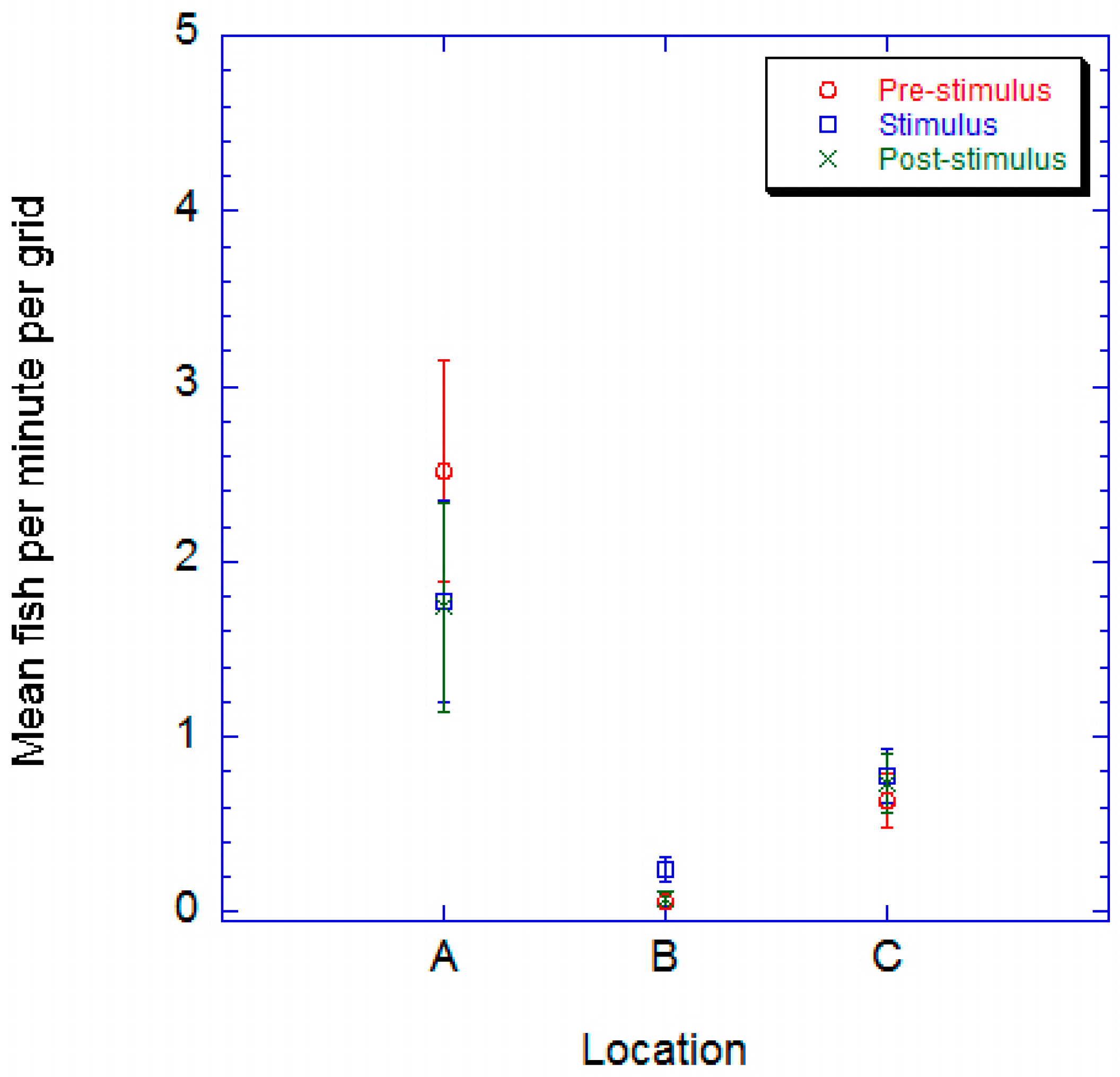
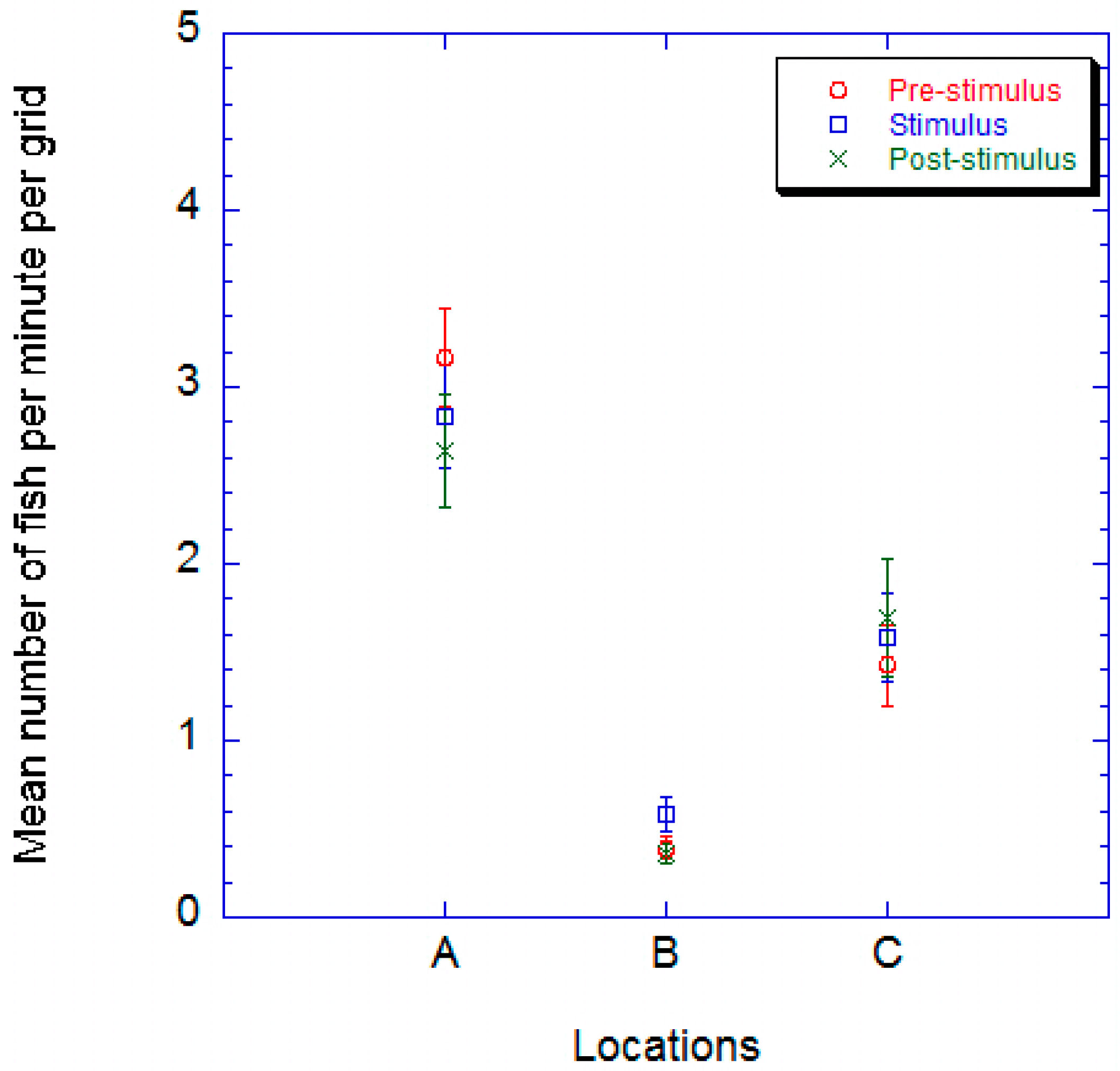
2. Results
2.1. Juvenile Channel Catfish
2.2. Juvenile Common Carp
3. Discussion
4. Materials and Methods
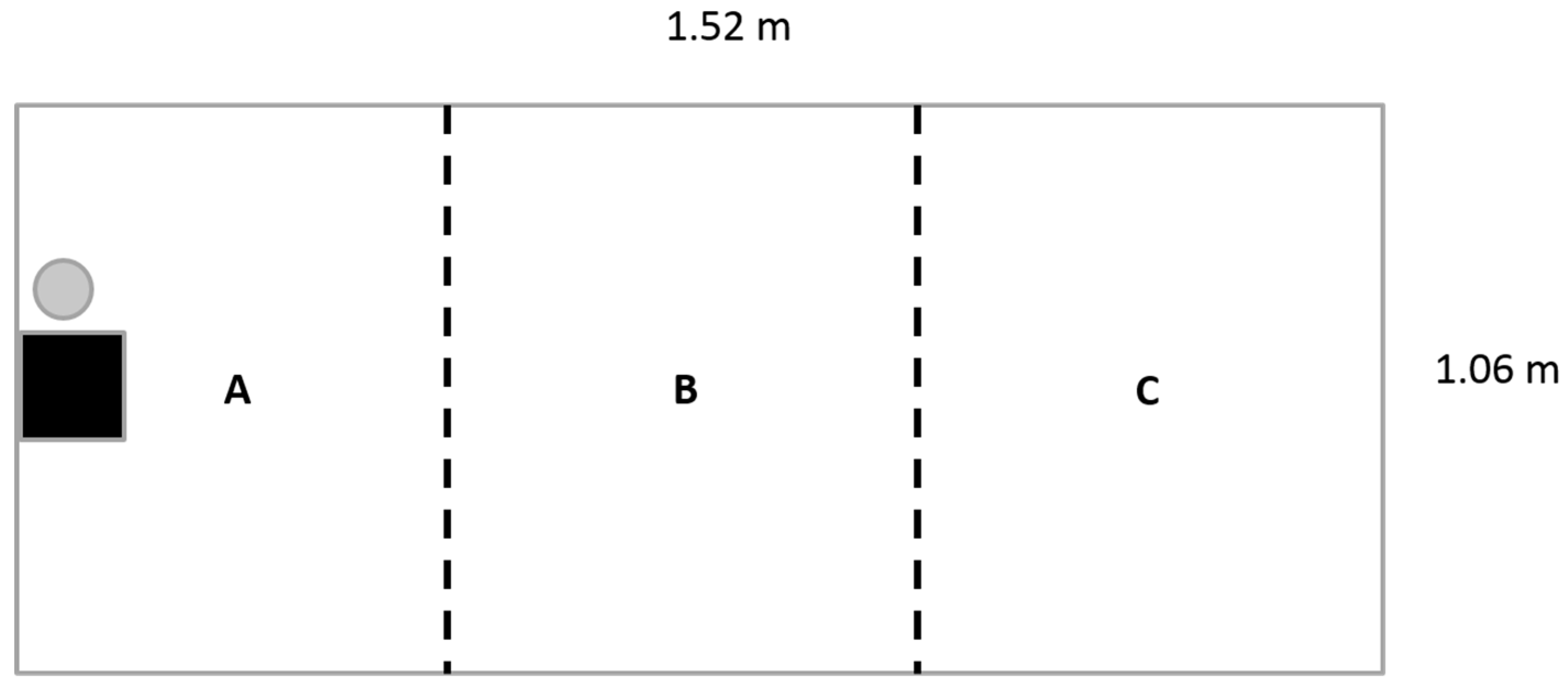
4.1. Experimental Subject and Set-Up
4.2. Data Collection and Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ricciardi, A. Patterns of invasion in the Laurentian Great Lakes in relation to changes in vector activity. Divers. Distrib. 2006, 12, 425–433. [Google Scholar] [CrossRef]
- Currie, W.J.S.; Kim, J.; Koops, M.A.; Mandrak, N.E.; O’Connor, L.M.; Pratt, T.C.; Timusk, E.; Choy, M. Modelling spread and assessing movement of Grass Carp, Ctenopharyngodon idella, in the Great Lakes basin. Can. Sci. Advis. Sec. Res. Doc. 2017. Available online: https://waves-vagues.dfo-mpo.gc.ca/Library/40600555.pdf (accessed on 5 November 2018).
- Cudmore, B.; Jones, L.A.; Mandrak, N.E.; Dettmers, J.M.; Chapman, D.C.; Kolar, C.S.; Conover, G. Ecological risk assessment of Grass Carp (Ctenopharyngodon idella) for the Great Lakes Basin. DFO Can. Sci. Advis. Sec. Res. Doc. 2017. Available online: https://waves-vagues.dfo-mpo.gc.ca/Library/4060116x.pdf (accessed on 1 November 2018).
- Taft, E.P.; Dixon, D.A.; Sullivan, C.W. Electric power research institute’s (EPRI) Research on behavioral technologies. Behav. Technol. Fish Guid. Am. Fish. Soc. Symp. 2001, 26, 115–124. [Google Scholar]
- Flammang, M.K.; Weber, M.J.; Thul, M.D. Laboratory evaluation of a bioacoustic bubble strobe light barrier for reducing walleye escapement. N. Am. J. Fish. Manag. 2014, 34, 1047–1054. [Google Scholar] [CrossRef]
- Noatch, M.R.; Suski, C.D. Non-physical barriers to deter fish movements. Environ. Rev. 2012, 20, 71–82. [Google Scholar] [CrossRef]
- Kim, J.; Mandrak, N.E. Effects of strobe lights on the behaviour of freshwater fishes. Environ. Biol. Fish. 2017, 100, 1427–1434. [Google Scholar] [CrossRef]
- Elvidge, C.K.; Ford, M.I.; Pratt, T.C.; Smokorowski, K.E.; Sills, M.; Patrick, P.H.; Cooke, S.J. Behavioural guidance of yellow-stage American eel Anguilla rostrata with a light-emitting diode device. Endanger. Species Res. 2018, 35, 159–168. [Google Scholar] [CrossRef]
- Vetter, B.J.; Murchy, K.A.; Cupp, A.R.; Amberg, J.J.; Gaikowski, M.P.; Mensinger, A.F. Acoustic deterrence of bighead carp (Hypophthalmichthys nobilis) to a broadband sound stimulus. J. Great Lakes Res. 2017, 43, 163–171. [Google Scholar] [CrossRef]
- Zielinski, D.P.; Sorensen, P.W. Silver, bighead and common carp orient to acoustic particle motion when avoiding a complex sound. PLoS ONE 2017, 12, e0180110. [Google Scholar] [CrossRef]
- Brown, G.E.; Ferrari, M.C.O.; Chivers, D.P. Learning about danger: Chemical alarm cues and threat-sensitive assessment of predation risk by fishes. In Fish Cognition and Behaviour, 2nd ed.; Brown, C., Laland, K.N., Krause, J., Eds.; Blackwell: London, UK, 2011; pp. 59–80. [Google Scholar]
- Kim, J.-W.; Wood, J.L.A.; Grant, J.W.A.; Brown, G.E. Acute and chronic increases in predation risk affect the territorial behaviour of juvenile Atlantic salmon in the wild. Anim. Behav. 2011, 81, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Bui, S.; Oppedal, F.; Korsøen, Ø.J.; Sonny, D.; Dempster, T. Group behavioural responses of Atlantic Salmon (Salmo salar L.) to light, infrasound and sound stimuli. PLoS ONE 2013, 8, e63696. [Google Scholar] [CrossRef]
- Hamel, M.J.; Brown, M.L.; Chipps, S.R. Behavioral responses of rainbow smelt to in situ strobe lights. N. Am. J. Fish. Manag. 2008, 28, 394–401. [Google Scholar] [CrossRef]
- Richards, N.S.; Chipps, S.R.; Brown, M.L. Stress response and avoidance behavior of fishes as influenced by high-frequency strobe lights. N. Am. J. Fish. Manag. 2007, 27, 1310–1315. [Google Scholar] [CrossRef]
- Sager, D.; Hocutt, C.; Stauffer, J. Estuarine fish responses to strobe light, bubble curtains and strobe light/bubble curtain combinations as influenced by water flow rate and flash frequencies. Fish. Res. 1987, 5, 383–399. [Google Scholar] [CrossRef]
- Sullivan, B.G.; Wilson, A.D.M.; Gutowsky, L.F.G.; Patrick, P.H.; Sills, M.; Cooke, S.J. The behavioral responses of a warmwater teleost to different spectra of light-emitting diodes. N. Am. J. Fish. Manag. 2016, 36, 1000–1005. [Google Scholar] [CrossRef]
- Königson, S.; Fjälling, A.; Lunneryd, S. Reactions in individual fish to strobe light. Field and aquarium experiments performed on whitefish (Coregonus lavaretus). Hydrobiologia 2002, 483, 39–44. [Google Scholar] [CrossRef]
- Patrick, P.H.; Christie, A.E.; Sager, D.; Hocutt, C.; Stauffer, J. Responses of fish to a strobe light/air-bubble barrier. Fish. Res. 1985, 3, 157–172. [Google Scholar] [CrossRef]
- Jesus, J.; Teixeira, A.; Natário, S.; Cortes, R. Repulsive effect of stroboscopic light barriers on native salmonid (Salmo trutta) and cyprinid (Pseudochondrostoma duriense and Luciobarbus bocagei) species of Iberia. Sustainability 2019, 11, 1332. [Google Scholar] [CrossRef]
- Mussen, T.D.; Patton, O.; Cocherell, D.; Ercan, A.; Bandeh, H.; Kavvas, M.L.; Post, J. Can behavioral fish-guidance devices protect juvenile Chinook salmon (Oncorhynchus tshawytscha) from entrainment into unscreened water-diversion pipes? Can. J. Fish. Aquat. Sci. 2014, 71, 1209–1219. [Google Scholar] [CrossRef]
- Lodge, D.M.; Williams, S.; MacIsaac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; McMichael, A. Biological invasions: Recommendations for U.S. policy and management. Ecol. Appl. 2006, 16, 2035–2054. [Google Scholar] [CrossRef]
- Britton, J.R.; Boar, R.R.; Grey, J.; Foster, J.; Lugonzo, J.; Harper, D.M. From introduction to fishery dominance: The initial impacts of the invasive carp Cyprinus carpio in Lake Naivasha, Kenya, 1999 to 2006. J. Fish. Biol. 2007, 71, 239–257. [Google Scholar] [CrossRef]
- Weber, M.J.; Brown, M.L. Effects of common carp on aquatic ecosystems 80 years after “carp as a dominant”: Ecological insights for fisheries management. Rev. Fish. Sci. 2009, 17, 524–537. [Google Scholar] [CrossRef]
- Vilizzi, L.; Thwaites, L.A.; Smith, B.B.; Nicol, M.J.; Madden, C.P. Ecological effects of common carp (Cyprinus carpio) in a semi-arid floodplain wetland. Mar. Freshw. Res. 2014, 65, 802–817. [Google Scholar] [CrossRef]
- Scott, W.B.; Crossman, E.J. Freshwater fishes of Canada. Fish. Res. Board Can. Bull. 1973, 184, 1–966. [Google Scholar]
- McLean, E.B.; Godin, J.-G.J. Distance to cover and fleeing from predators in fish with different amounts of defensive armour. Oikos 1989, 55, 281–290. [Google Scholar] [CrossRef]
- Venter, O.; Grant, J.W.A.; Noël, M.V.; Kim, J.-W. Mechanisms underlying the increase in young-of-the-year Atlantic salmon (Salmo salar) density with habitat complexity. Can. J. Fish. Aquat. Sci. 2008, 65, 1956–1964. [Google Scholar] [CrossRef]
- Mesquita, F.de.O.; Godinho, H.P.; Azevedo, P.G.de.; Young, R.J. A preliminary study into the effectiveness of stroboscopic light as an aversive stimulus for fish. Appl. Anim. Behav. Sci. 2008, 111, 402–407. [Google Scholar] [CrossRef]
- Milinski, M.; Lüthi, J. Cooperation under predation risk: Experiments on costs and benefits. Proc. R. Soc. Lond. B 1997, 264, 831–837. [Google Scholar] [CrossRef]
- Wrona, F.J.; Dixon, R.W.J. Group size and predation risk: A field analysis of encounter and dilution effects. Am. Nat. 1991, 137, 186–201. [Google Scholar] [CrossRef]
- Stewart, H.A.; Wolter, M.H.; Wahl, D.H. Laboratory investigations on the use of strobe lights and bubble curtains to deter dam escapes of age-0 muskellunge. N. Am. J. Fish. Manag. 2014, 34, 571–575. [Google Scholar] [CrossRef]
- McMahon, T.E.; Holanov, S.H. Foraging success of largemouth bass at different light intensities: Implications for time and depth of feeding. J. Fish. Biol. 1995, 46, 759–767. [Google Scholar] [CrossRef]
- McFarland, W.N.; Loew, E.R. Wave produced changes in underwater light and their relations to vision. Environ. Biol. Fish. 1983, 8, 173–184. [Google Scholar] [CrossRef]
- Chivers, D.P.; Brown, G.E.; Smith, R.J.F. Familiarity and shoal cohesion in fathead minnows (Pimephales promelas): Implications for antipredator behaviour. Can. J. Zool. 1995, 73, 955–960. [Google Scholar] [CrossRef]
- Hoare, D.J.; Couzin, I.D.; Godin, J.-G.J.; Krause, J. Context-dependent group size choice in fish. Anim. Behav. 2004, 67, 155–164. [Google Scholar] [CrossRef]
- Blumstein, D.T.; Evans, C.S.; Daniel, J.C. JWatcher 1.0; Animal Behaviour Laboratory Macquarie University: Sydney, Australia, 2000. [Google Scholar]
- Blumstein, D.T.; Evans, C.S.; Sunderland, M.A. Quantifying behavior the JWatcher Way; Sinauer Associates Inc.: Sunderland, MA, USA, 2007; 211p. [Google Scholar]
- Herbert-Read, J.E.; Rosén, E.; Szorkovszky, A.; Ioannou, C.C.; Rogell, B.; Perna, A.; Ramnarine, I.W.; Kotrschal, A.; Kolm, N.; Krause, J.; et al. How predation shapes the social interaction rules of shoaling fish. Proc. R. Biol. Soc. B 2017, 284, 20171126. [Google Scholar] [CrossRef] [Green Version]
- Imada, H.; Hoki, M.; Suehiro, Y.; Okuyama, T.; Kurabayashi, D.; Shimada, A.; Naruse, K.; Takeda, H.; Kubo, T.; Takeuchi, H. Coordinated and cohesive movement of two small conspecific fish induced by eliciting a simultaneous optomotor response. PLoS ONE 2010, 5, e11248. [Google Scholar] [CrossRef]
- Kim, J.-W.; Grant, J.W.A. Effects of patch shape and group size on the effectiveness of defence by juvenile convict cichlids. Anim. Behav. 2007, 73, 275–280. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cohesion Group and Points | Number of Fish | Example Configurations |
|---|---|---|
| 0 | 1 |  |
| 1 | 2 |  |
| 2 | 3 in a line |  |
| 3 | 3 in tight group or 4 in a line |  |
| 4 | 4 in tight group or 5 in a line |  |
| 5 | 5 in tight group |  |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Bondy, C.; Chandler, C.M.; Mandrak, N.E. Behavioural Response of Juvenile Common Carp (Cyprinus carpio) and Juvenile Channel Catfish (Ictalurus punctatus) to Strobe Light. Fishes 2019, 4, 29. https://doi.org/10.3390/fishes4020029
Kim J, Bondy C, Chandler CM, Mandrak NE. Behavioural Response of Juvenile Common Carp (Cyprinus carpio) and Juvenile Channel Catfish (Ictalurus punctatus) to Strobe Light. Fishes. 2019; 4(2):29. https://doi.org/10.3390/fishes4020029
Chicago/Turabian StyleKim, Jaewoo, Caitlyn Bondy, Catherine M. Chandler, and Nicholas E. Mandrak. 2019. "Behavioural Response of Juvenile Common Carp (Cyprinus carpio) and Juvenile Channel Catfish (Ictalurus punctatus) to Strobe Light" Fishes 4, no. 2: 29. https://doi.org/10.3390/fishes4020029
APA StyleKim, J., Bondy, C., Chandler, C. M., & Mandrak, N. E. (2019). Behavioural Response of Juvenile Common Carp (Cyprinus carpio) and Juvenile Channel Catfish (Ictalurus punctatus) to Strobe Light. Fishes, 4(2), 29. https://doi.org/10.3390/fishes4020029