
Spatio-Temporal Variations in the Potential Habitat Distribution of Pacific Sardine (Sardinops sagax) in the Northwest Pacific Ocean
Abstract
:1. Introduction
2. Materials and Methods
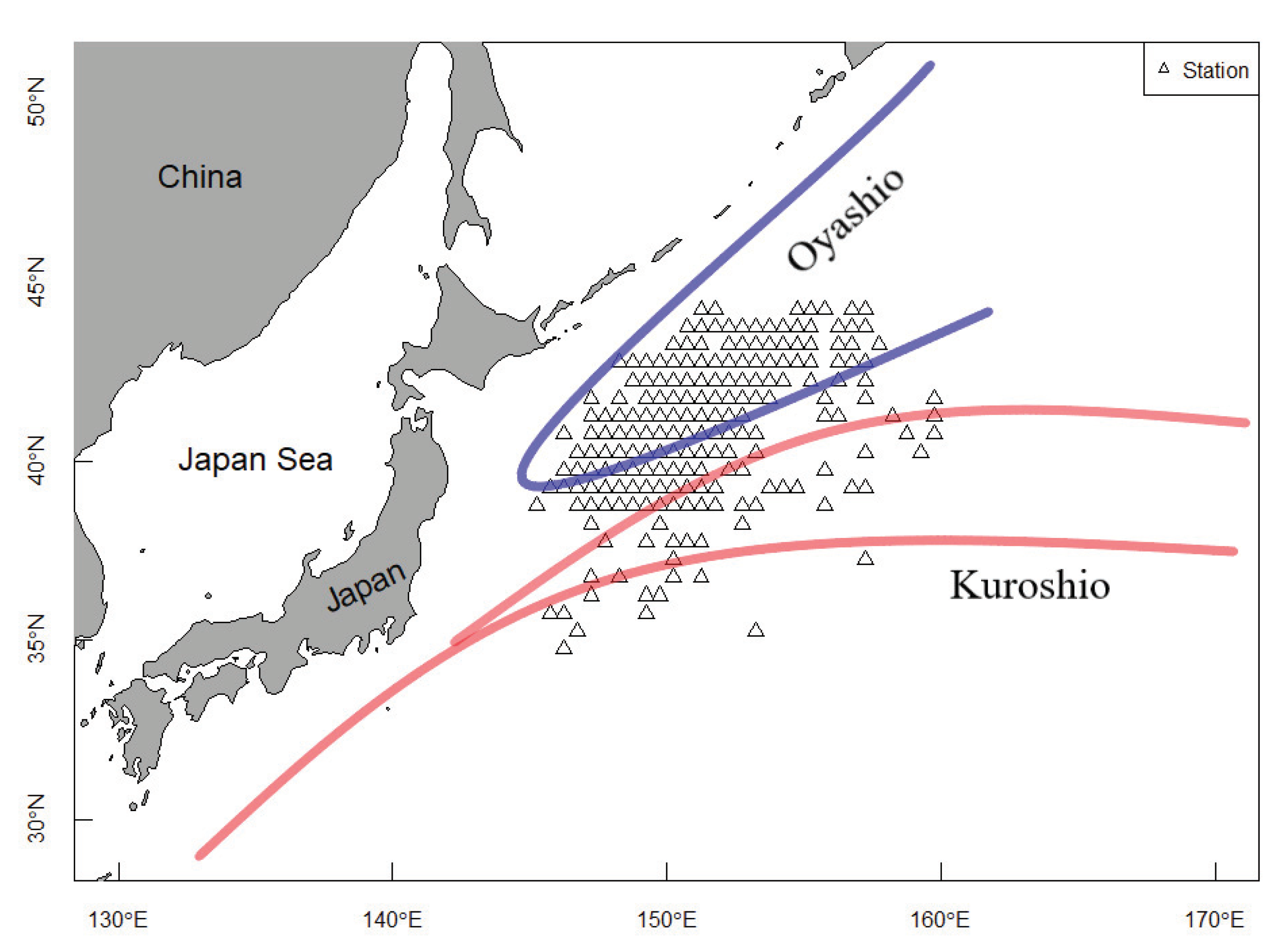
2.1. Study Area
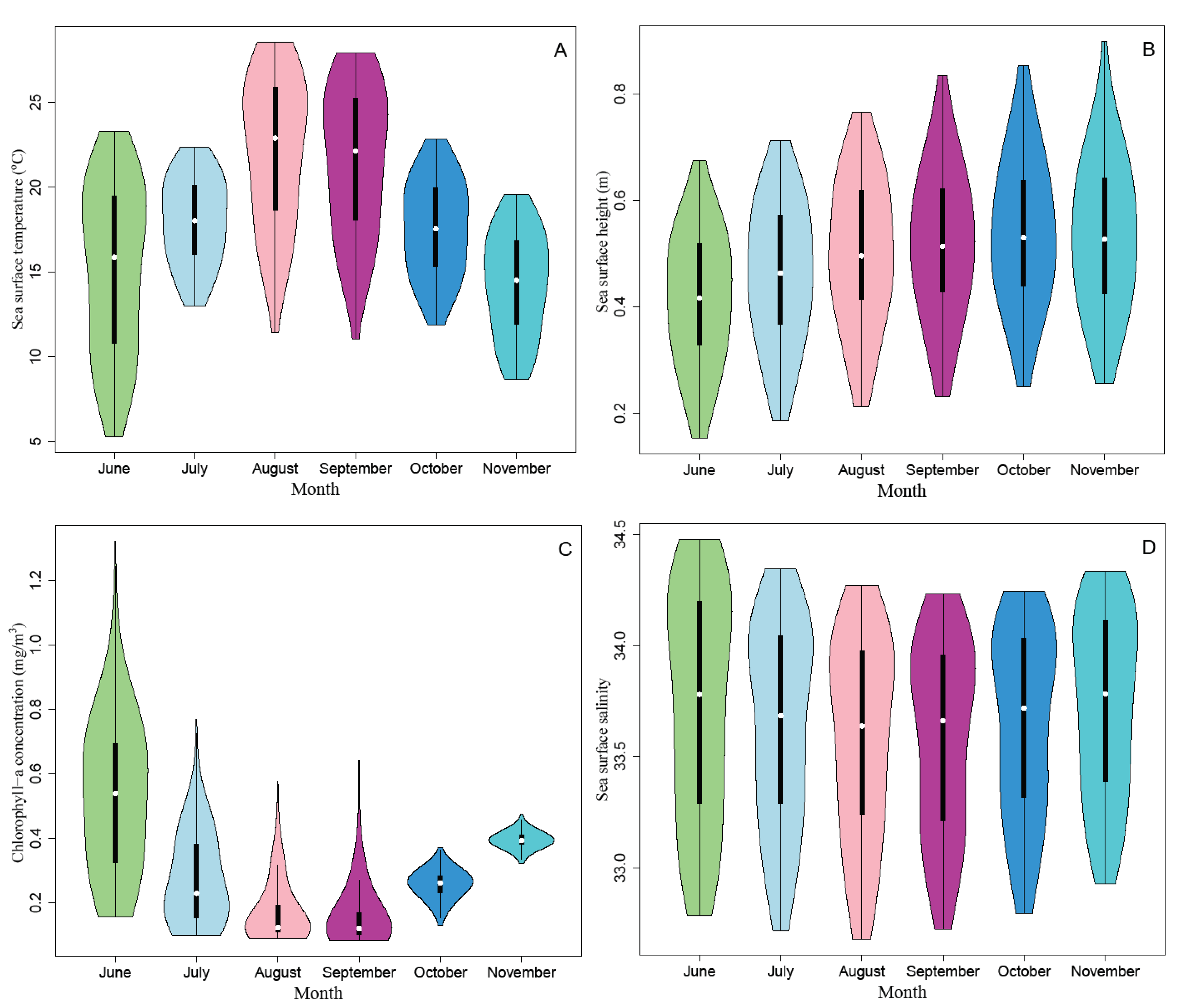
2.2. Data Resources
2.3. Modelling Procedure
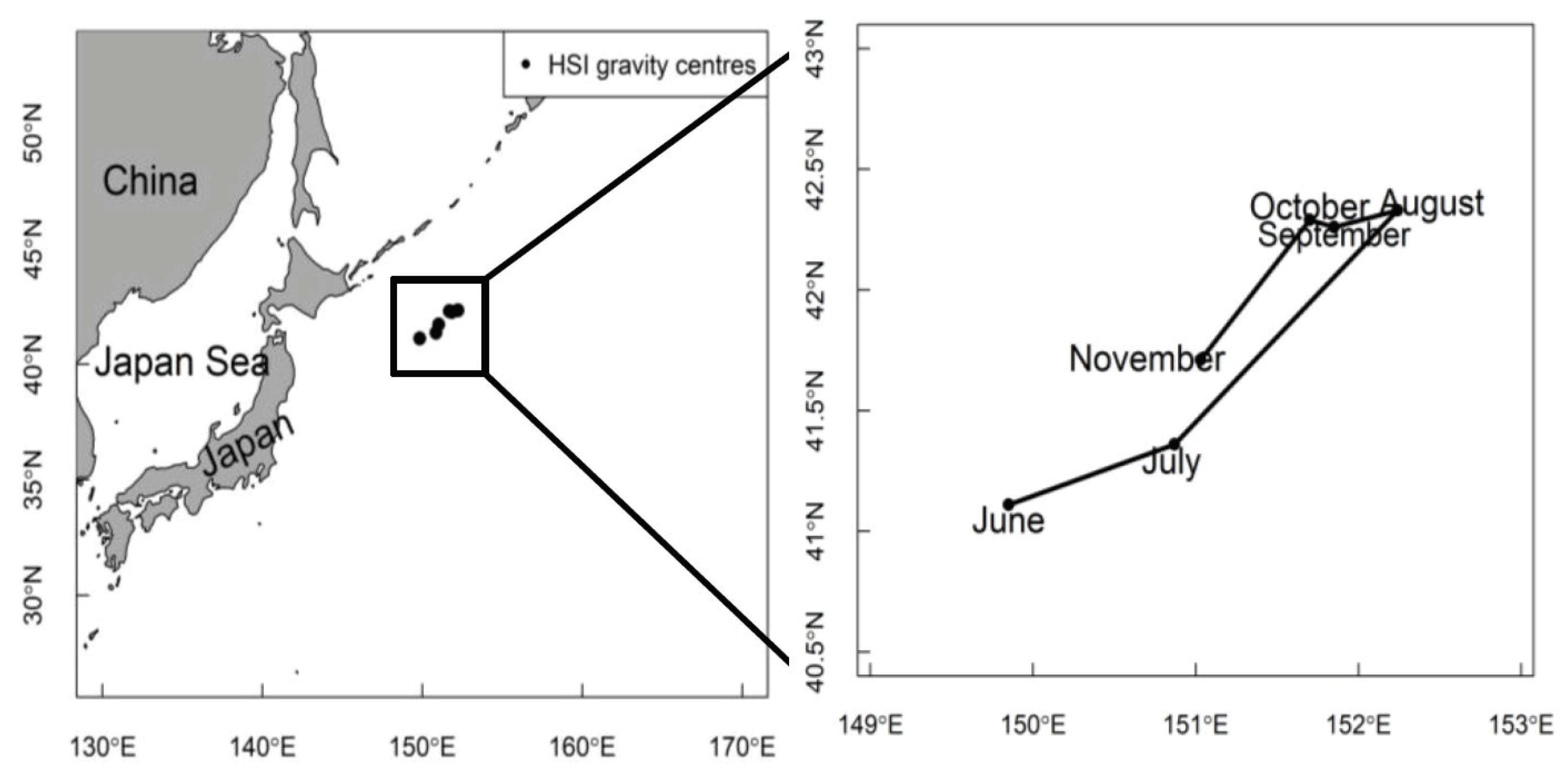
2.4. Centroid Shifts
3. Results
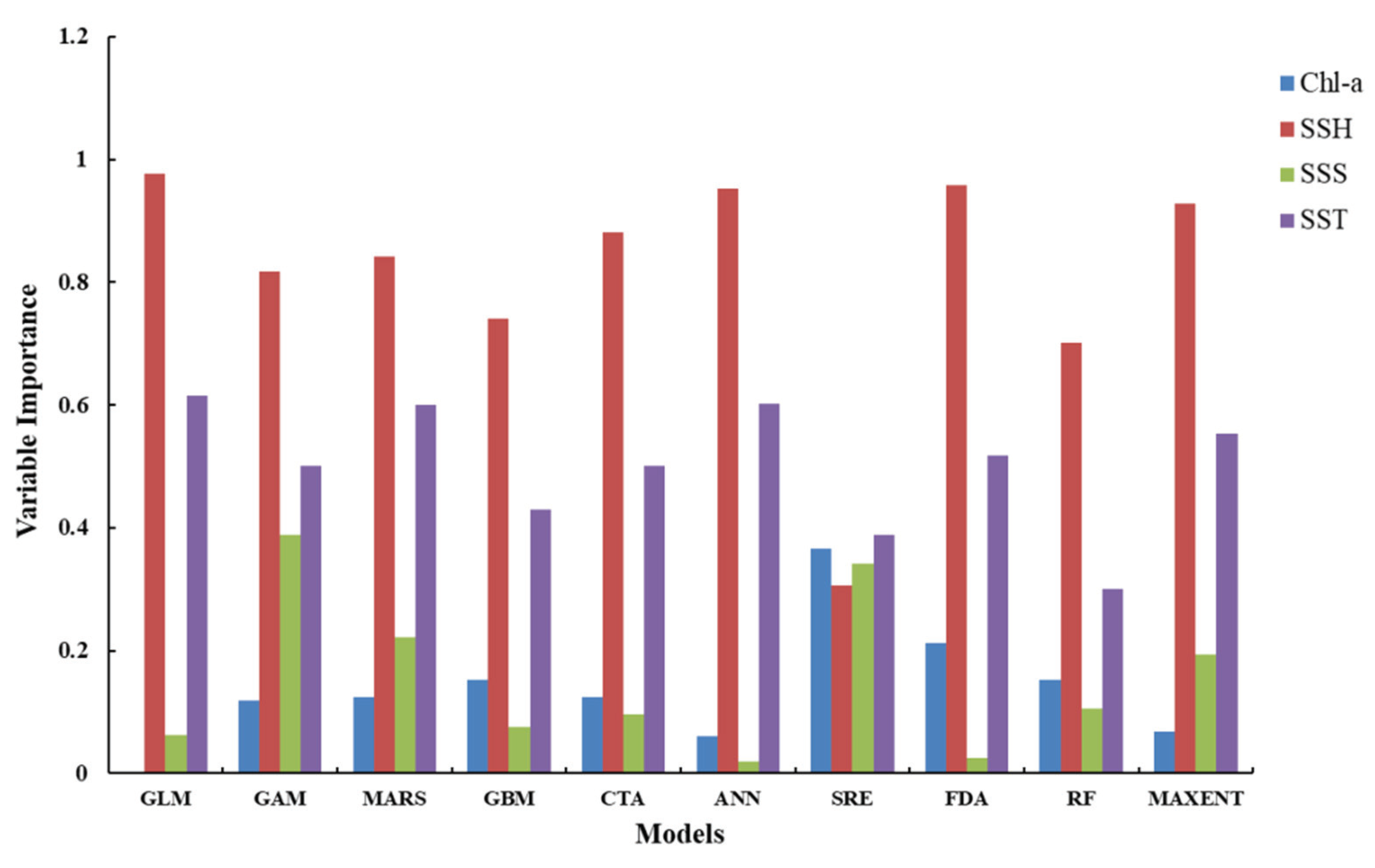
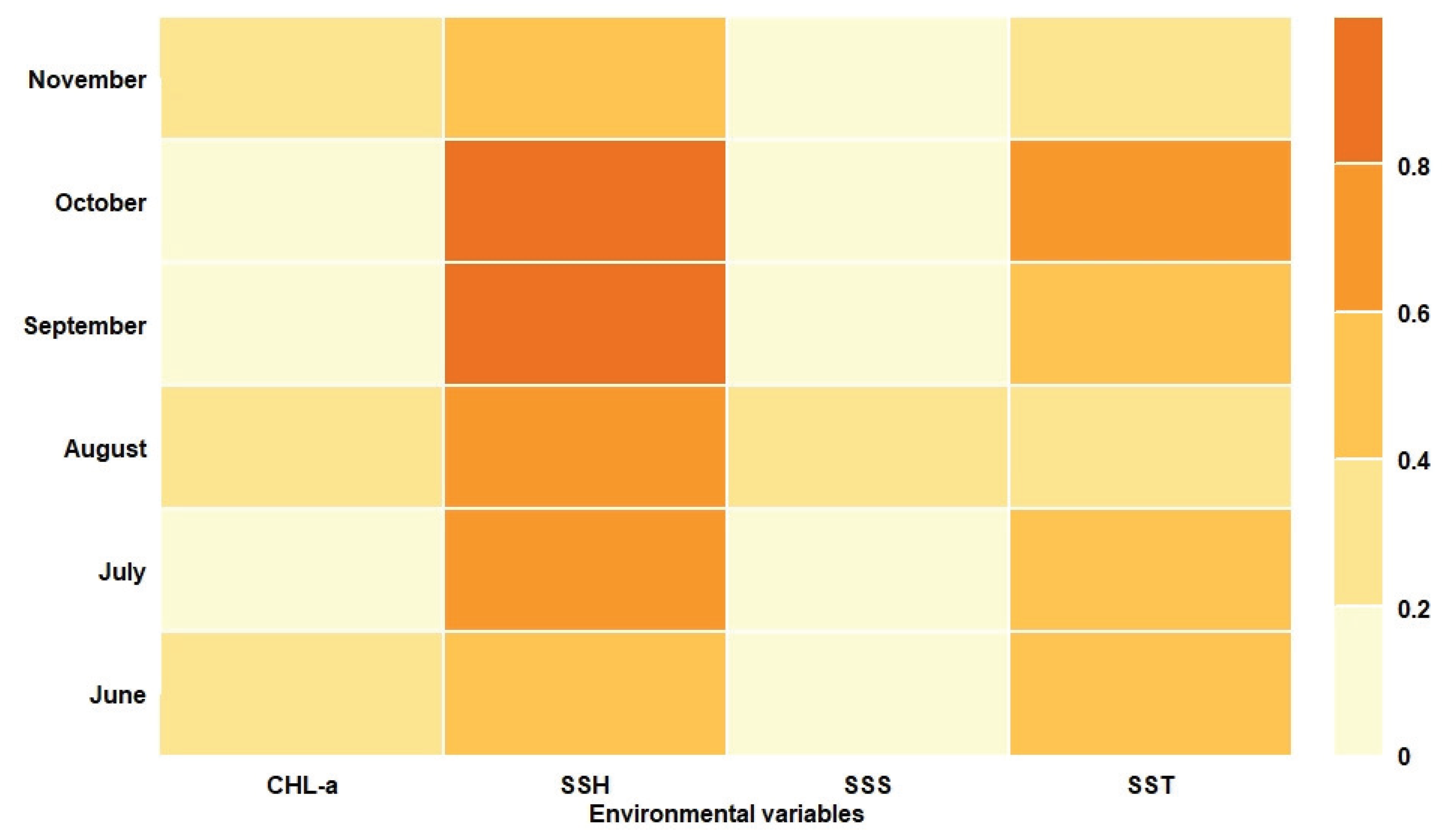
3.1. Single-Algorithm Models Performances
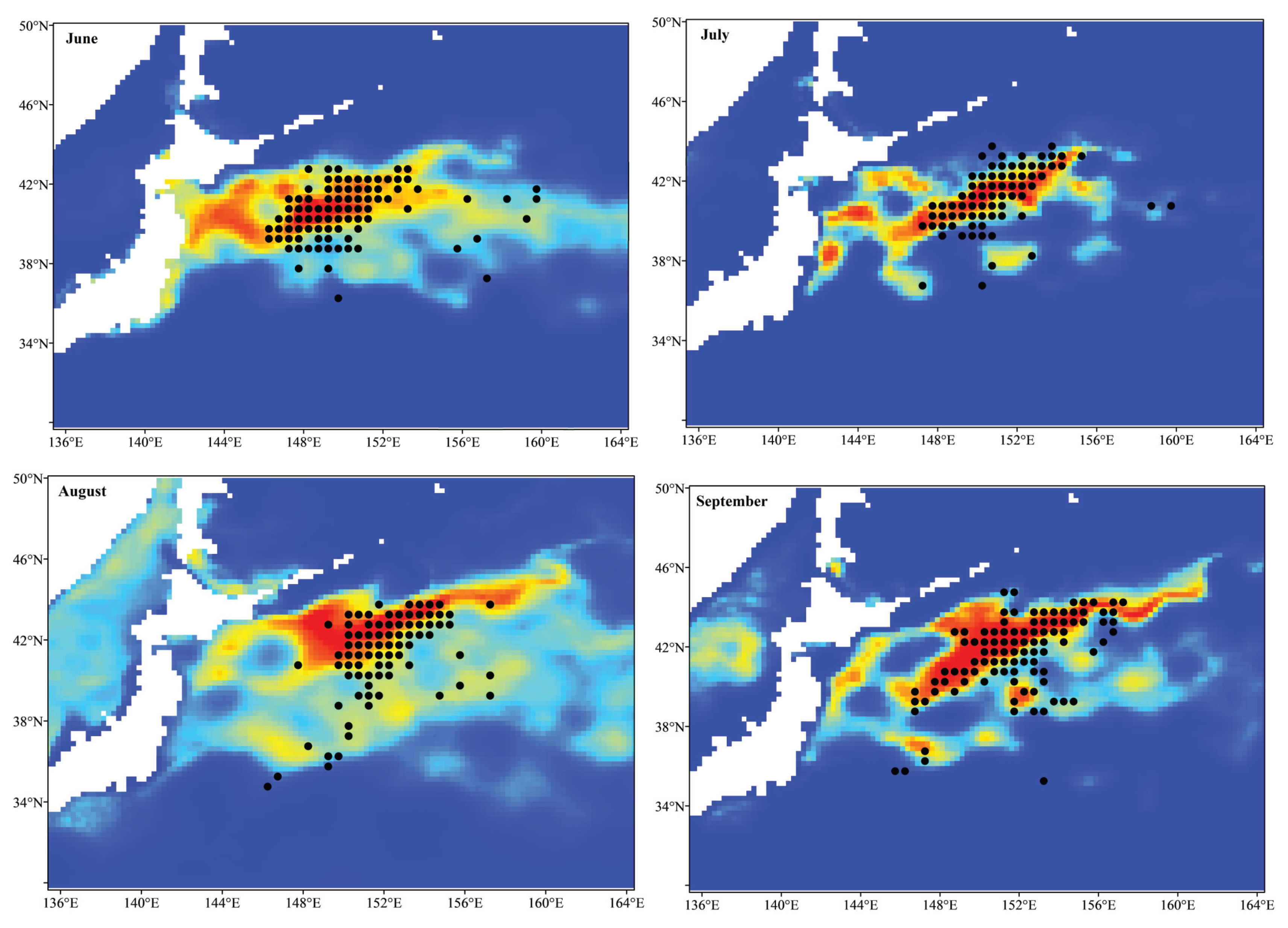
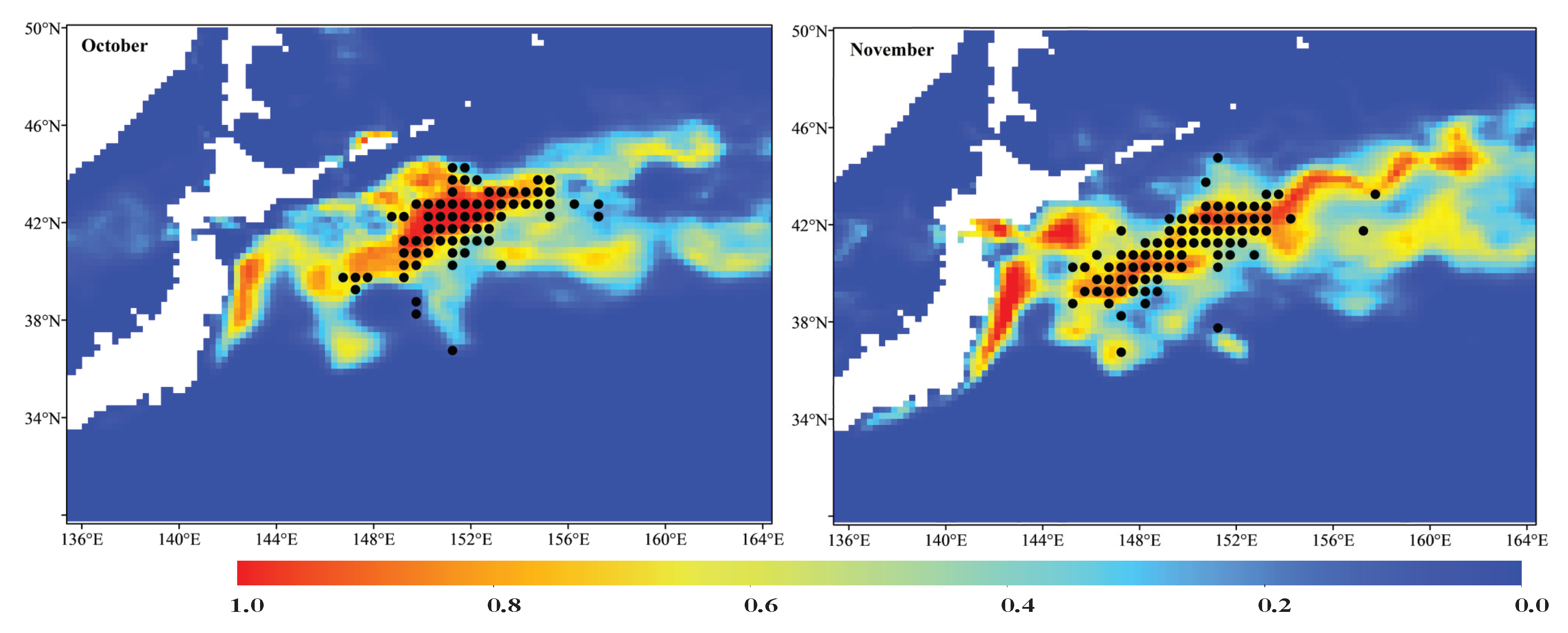
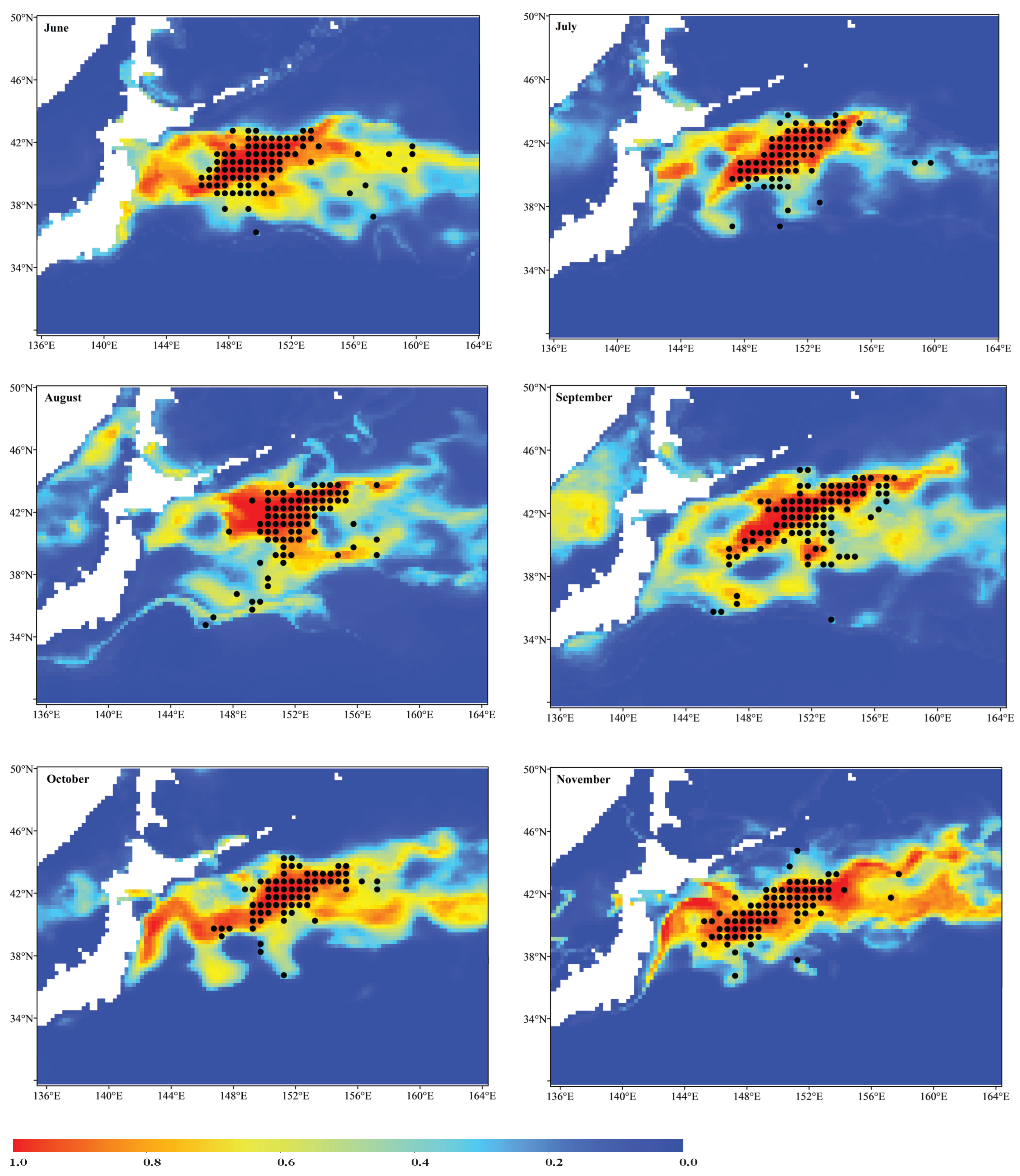
3.2. Ensemble Model Prediction and Potential Habitat Distribution of the Pacific Sardine
3.3. Centroid Shifts of Pacific Sardine Fishing Grounds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Methot, R.D.J.; Wetzel, C. Stock synthesis: A biological and statistical framework for fish stock assessment and fishery management. Fish. Res. 2013, 142, 86–99. [Google Scholar] [CrossRef]
- Cheung, W.W.L.; Lam, V.W.Y.; Sarmiento, J.L.; Kearney, K.; Watson, R.E.G.; Pauly, D. Projecting global marine biodiversity impacts under climate change scenarios. Fish Fish. 2009, 10, 235–251. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Brown, C.J.; Sydeman, W.J.; Kiessling, W.; Burrows, M.T. Global imprint of climate change on marine life. Nat. Clim. Chang. 2013, 3, 919–925. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.J. Ocean warming-induced range shifting of potential habitat for jumbo flying squid Dosidicus gigas in the southeast Pacific Ocean off Peru. Fish. Res. 2018, 204, 137–146. [Google Scholar] [CrossRef]
- Stramma, L.; Johnson, G.C.; Sprintall, J.; Mohrholz, V. Expanding oxygen minimum zones in the tropical oceans. Science 2008, 320, 655–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, N.D.; Levin, L.A. Fish ecology and evolution in the world’s oxygen minimum zones and implications of ocean deoxygenation. Adv. Mar. Biol. 2016, 74, 117–198. [Google Scholar] [PubMed]
- Cortes, G.G.; Montanez, J.A.D.A.; Sánchez, F.A.; Salas, S.; Balart, E.F. How do environmental factors affect the stock–recruitment relationship? The case of the Pacific sardine (Sardinops sagax) of the northeastern Pacific Ocean. Fish. Res. 2010, 102, 173–183. [Google Scholar] [CrossRef]
- Guan, X.D. Sardinops sagax from the coast of Japan. Mar. Fish. 1985, 4, 187–189, (In Chinese with English Abstract). [Google Scholar]
- Wei, C.; Chen, Y.Z.; Zhou, B.B.; Sun, J.M. Identification of Sardinops sagax populations in the Yellow Sea of China. Mar. Sci. 1989, 4, 55–60, (In Chinese with English Abstract). [Google Scholar]
- Morimoto, H. Age and growth of Japanese sardine (Sardinops melanostictus) in Tosa Bay, southwestern Japan during a period of declining stock size. Fish. Sci. 2003, 69, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Yatsu, A.; Kawabata, A. Reconsidering Trans-Pacific “synchrony” in population fluctuations of sardines. Bulletin J. Soc. Fish. Oceanogr. 2017, 81, 271–283. [Google Scholar]
- Suda, M.; Kishida, T. A spatial model of population dynamics of the early life stages of Japanese sardine, Sardinops melanostictus, off the Pacific coast of Japan. Fish. Oceanogr. 2003, 12, 85–99. [Google Scholar] [CrossRef]
- Michio, Y.; Tanaka, H.; Honda, S.; Nishida, H.; Nashida, K.; Hirota, Y.; Ishida, M.; Ohshimo, S.; Miyabe, S.; Ito, H. Sex-ual maturation, spawning period and batch fecundity of Japanese sardine (Sardinops melanostictus) in the coastal waters of western Japan in 2008–2010. Bull. Jpn. Soc. Fish. Oceanogr. 2013, 77, 59–67. [Google Scholar]
- Nyuji, M.; Hongo, Y.; Yoneda, M.; Nakamura, M. Transcriptome characterization of BPG axis and expression profiles of ovarian steroidogenesis-related genes in the Japanese sardine. BMC Genom. 2020, 21, 668. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Zenitani, H.; Kimura, R. Population decline of the Japanese sardine Sardinops melanostictus owing to recruitment failures. Can. J. Fish. Aquat. Sci. 1995, 52, 1609–1616. [Google Scholar] [CrossRef]
- Kinoshita, T. Northward migrating juveniles in the Kuroshio Extension area. In Stock Fluctuations and Ecological Changes of the Japanese Sardine; Watanabe, Y., Wada, T., Eds.; Koseisyakoseikaku: Tokyo, Japan, 1998; pp. 84–92, (In Japanese with English Abstract). [Google Scholar]
- Kuroda, K. Studies on the recruitment process focusing on the early life history of the Japanese sardine, Sardinops melanostictus (Schelegen). Bull. Natl. Res. Inst. Fish. Sci. 1991, 3, 25–278, (In Japanese with English Abstract). [Google Scholar]
- Wang, M.Y.; Dong, W.D. Development and utilization of the Sardinops sagax. Fish. Sci. 1992, 7, 14–16, (In Chinese with English Abstract). [Google Scholar]
- Shi, Y.C.; Zhang, X.M.; He, Y.R.; Fan, W.; Tang, F.H. Stock Assessment Using Length-Based Bayesian Evaluation Method for Three Small Pelagic Species in the Northwest Pacific Ocean. Front. Mar. Sci. 2022, 9, 775180. [Google Scholar] [CrossRef]
- Zhao, G.Q.; Shi, Y.C.; Fan, W.; Cui, X.S.; Tang, F.H. Study on main catch composition and fishing ground change of light purse seine in Northwest Pacific. South China Fish. Sci. 2022, 18, 33–42, (In Chinese with English Abstract). [Google Scholar]
- North Pacific Fisheries Commission. NPFC Yearbook 2017. 2017, p. 385. Available online: www.npfc.int (accessed on 21 November 2017).
- Brochier, T.; Echevin, V.; Tam, J.; Chaigneau, A.; Goubanova, K.; Bertrand, A. Climate change scenarios experiments predict a future reduction in small pelagic fish recruitment in the Humboldt Current system. Glob. Chang. Biol. 2013, 19, 1841–1853. [Google Scholar] [CrossRef]
- Ishimura, G.; Herrick, S.; Sumaila, U.R. Stability of cooperative management of the Pacific sardine fishery under climate variability. Mar. Policy 2013, 39, 333–340. [Google Scholar] [CrossRef]
- Chen, C.S.; Chiu, T.S. Abundance and spatial variation of Ommastrephes bartramii in the eastern North Pacific observed from an exploratory survey. Acta Zool. Taiwan 1999, 10, 135–144. [Google Scholar]
- Lacomte, F.; Grant, W.S.; Dodson, J.J.; Rodriguez-Sanchez, R.; Bowen, B.W. Living with uncertainty; genetic imprints of climate shifts in east Pacific anchovy (Engraulis mordax) and sardine (Sardinops sagax). Mol. Ecol. 2004, 13, 2169–2182. [Google Scholar] [CrossRef]
- Porchas, M.M.; Rodríguez, M.H.; Ramírez, L.F.B. Thermal behavior of the Pacific sardine (Sardinops sagax) acclimated to different thermal cycles. J. Therm. Biol. 2009, 34, 372–376. [Google Scholar] [CrossRef]
- Dudarev, V.A. Oceanological Principles of Distribution, Migration, and Dynamics of Population of Far Eastern Sardine. In Gidrometeorologiya i Gidrokhimiya Morei, Tom 8. Yaponskoe More; Hydrometeorology and Hydrochemistry of the Seas, Vol. 8: Sea of Japan; Gidrometeoizdat: St. Petersburg, Russia, 2004; Volume 8, pp. 229–234. [Google Scholar]
- Vander, L.C.D.; Castro, L.; Drapeau, L.; Checkley, D., Jr. (Eds.) Report of a GLOBEC-SPACC Workshop on Characterizing and Comparing the Spawning Habitats of Small Pelagic Fish; GLOBEC Report 21; GLOBEC: Swindon, UK, 2005; Volume 7, p. 33. [Google Scholar]
- Takasuka, A.; Oozeki, Y.; Aoki, I. Optimal growth temperature hypothesis: Why do anchovy flourish and sardine collapse or vice versa under the same ocean regime? Can. J. Fish. Aquat. Sci. 2007, 64, 768–776. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. S. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, S.; Capinha, C.; Weterings, R.; Gao, T. Using species distribution model to predict the impact of climate change on the potential distribution of Japanese whiting Sillago japonica. Ecol. Indic. 2019, 104, 333–340. [Google Scholar] [CrossRef]
- Gong, C.X.; Chen, X.J.; Gao, F.; Yu, W. The Change Characteristics of Potential Habitat and Fishing Season for Neon Flying Squid in the Northwest Pacific Ocean under Future Climate Change Scenarios. Mar. Coast. Fish. 2021, 13, 450–462. [Google Scholar] [CrossRef]
- Guénard, G.; Morin, J.; Matte, P.; Secretan, Y.; Valiquette, E.; Mingelbier, M. Deep learning habitat modeling for moving organisms in rapidly changing estuarine environments: A case of two fishes. Estuar. Coast. Shelf Sci. 2020, 238, 106713. [Google Scholar] [CrossRef]
- Zhang, X.M.; Shi, Y.C.; Li, F.; Zhu, M.M.; Wei, Z.H. Prediction of potential fishing ground for Pacific saury (Cololabis saira) based on MAXENT model. J. Shanghai Ocean Univ. 2020, 29, 280–286, (In Chinese with English Abstract). [Google Scholar]
- Hao, T.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. Testing whether ensemble modelling is advantageous for maximising predictive performance of species distribution models. Ecography 2020, 43, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Ahmed, S.E.; McInerny, G.; O’Hara, K.; Harper, R.; Salido, L.; Emmott, S.; Joppa, L.N. Scientists and software-surveying the species distribution modeling community. Divers. Distrib. 2015, 21, 258–267. [Google Scholar] [CrossRef]
- Thuiller, W. Biomod: Optimizing predictions of species distributions and projecting potential future shifts under global change. Glob. Chang. Biol. 2003, 9, 1353–1362. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. Biomod: A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Alabia, I.D.; Saitoh, S.I.; Igarashi, H.; Ishikawa, Y.; Usui, N.; Kamachi, M.; Awaji, T.; Seito, M. Ensemble squid habitat model using three-dimensional ocean data. ICES J. Mar. Sci. 2016, 73, 1863–1874. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Huang, Y.; Zhao, H.; Yang, M.; Zhuang, Y.; Ye, X. Modelling the Effects of Climate Change on the Distribution of Endangered Cypripedium japonicum in China. Forests 2021, 12, 429. [Google Scholar] [CrossRef]
- Zhao, G.H.; Cui, X.Y.; Sun, J.J.; Li, T.T.; Wang, Q.; Ye, X.Z.; Fan, B.G. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Indic. 2021, 132, 108256. [Google Scholar] [CrossRef]
- Logerwell, E.A.; Smith, P.E. Mesoscale eddies and survival of late stage Pacific sardine (Sardinops sagax) larvae. Fish. Oceanogr. 2010, 10, 13–25. [Google Scholar] [CrossRef]
- Ma, C.; Zhuang, Z.D.; Liu, Y.; Xu, C.Y.; Cai, J.D.; Shen, C.C. Preliminary study on catch composition and biological characteristics of main species of light-lift net in the Northwest Pacific Ocean. J. Fish. Res. 2018, 40, 141–147, (In Chinese with English Abstract). [Google Scholar]
- Assis, J.; Tyberghein, L.; Bosh, S.; Verbruggen, H.; Serrão, E.A.; Clerck, D.O. Bio-ORACLE v2.0: Extending marine data layers for bioclimatic modelling. Glob. Ecol. Biogeogr. 2017, 27, 277–284. [Google Scholar] [CrossRef]
- Petatán, R.D.; Ojeda, R.M.Á.; Sánchez, V.L.; Rivas, D.; Reyes, B.H.; Cruz, P.G.; Morzaria, L.H.N.; Cisneros, M.A.M.; Cheung, W.; Salvadeo, C. Potential changes in the distribution of suitable habitat for Pacific sardine (Sardinops sagax) under climate change scenarios. Deep Sea Res. Part II 2019, 169–170, 104632. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carr’e, G.; Marqu´ez, J.R.G.; Gruber, B.; Lafourcade, B.; Leitao, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Tien, B.D.; Lofman, O.; Revhaug, I.; Dick, O. Landslide susceptibility analysis in the Hoa Binh province of Vietnam using statistical index and logistic regression. Nat. Hazards 2011, 59, 1413–1444. [Google Scholar]
- Chen, Y.L.; Shan, X.J.; Ovando, D.; Yang, T.; Dai, F.Q.; Jin, X.S. Predicting current and future global distribution of black rockfish (Sebastes schlegelii) under changing climate. Ecol. Indic. 2021, 128, 107799. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez, V.A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Pearce, J.; Ferrier, S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Modell. 2000, 133, 225–245. [Google Scholar] [CrossRef] [Green Version]
- Phillips, N.D.; Reid, N.; Thys, T.; Harrod, C.; Payne, N.L.; Morgan, C.A.; White, H.J.; Porter, S.; Houghton, J.D.R. Applying species distribution modelling to a data poor, pelagic fish complex: The ocean sunfishes. J. Biogeogr. 2017, 44, 2176–2187. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.P.; Aguiar, A.G.; Simiao-Ferreira, J. Assessing the distribution and conservation status of a long-horned beetle with species distribution models. J. Insect. Conserv. 2016, 20, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.J.; Tian, S.Q.; Chen, Y.; Liu, B.L. A modeling approach to identify optimal habitat and suitable fishing grounds for neon flying squid (Ommastrephes bartramii) in the Northwest Pacific Ocean. Fish. Bull. 2010, 108, 1–15. [Google Scholar]
- Yu, W.; Wen, J.; Zhang, Z.; Chen, X.J.; Zhang, Y. Spatio-temporal variations in the potential habitat of a pelagic commercial squid. J. Mar. Syst. 2020, 206, 103339. [Google Scholar] [CrossRef]
- Li, G.; Cao, J.; Zou, X.; Chen, X.J.; Runnebaum, J. Modeling habitat suitability index for Chilean jack mackerel (Trachurus murphyi) in the South East Pacific. Fish. Res. 2016, 178, 47–60. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Arroita, G.G.; Monfort, J.J.L.; Elith, J.; Gordon, A.; Kujala, H.; Lentini, P.E.; McCarthy, M.A.; Tingley, R.; Wintle, B.A. Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 2015, 24, 276–292. [Google Scholar] [CrossRef]
- Takasuka, A.; Kuroda, H.; Okunishi, T.; Shimizu, Y.; Hirota, Y.; Kubota, H.; Sakajl, H.; Kimura, R.; Ito, S.I.; Oozeki, Y. Occurrence and density of Pacific saury Cololabis saira larvae and juveniles in relation to environmental factors during the winter spawning season in the Kuroshio Current system. Fish. Oceanogr. 2014, 23, 304–321. [Google Scholar] [CrossRef]
- Mainali, K.P.; Warren, D.L.; Dhileepan, K.; McConnachie, A.; Strathie, L.; Hassan, G.; Karki, D.; Shrestha, B.B.; Parmesan, C. Projecting future expansion of invasive species: Comparing and improving methodologies for species distribution modeling. Glob. Chang. Biol. 2015, 21, 4464–4480. [Google Scholar] [CrossRef] [Green Version]
- Watling, J.I.; Brandt, L.A.; Bucklin, D.N.; Fujisaki, I.; Mazzotti, F.J.; Romañach, S.S.; Speroterra, C. Performance metrics and variance partitioning reveal sources of uncertainty in species distribution models. Ecol. Modell. 2015, 309–310, 48–59. [Google Scholar] [CrossRef]
- Scales, K.L.; Miller, P.I.; Ingram, S.N.; Hazen, E.L.; Bograd, S.J.; Phillips, R.A. Identifying predictable foraging habitats for a wide-ranging marine predator using ensemble ecological niche models. Divers. Distrib. 2016, 22, 212–224. [Google Scholar] [CrossRef] [Green Version]
- Arkhipkin, A.I.; Rodhouse, P.G.K.; Pierce, G.J.; Sauer, W.; Sakai, M.; Allcock, L.; Arguelles, J.; Bower, J.R.; Castillo, G. World squid fisheries. Rev. Fish. Sci. Aquac. 2015, 23, 92–252. [Google Scholar] [CrossRef] [Green Version]
- Weber, E.D.; McClatchie, S. Predictive models of northern anchovy Engraulis mordax and Pacific sardine Sardinops sagax spawning habitat in the California Current. Mar. Ecol. Prog. Ser. 2010, 406, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Ichii, T.; Mahapatra, K.; Sakai, M.; Okada, Y. Life history of the neon flying squid: Effect of the oceanographic regime in the North Pacific Ocean. Mar. Ecol. Prog. Ser. 2009, 378, 1–11. [Google Scholar] [CrossRef]
- Song, H.; Miller, A.J.; McClatchie, S.; Weber, E.D.; Nieto, K.M.; Checkley, D.M.J. Application of a data-assimilation model to variability of Pacific sardine spawning and survivor habitats with ENSO in the California Current System. J. Geophys. Res. 2012, 117, C03009. [Google Scholar] [CrossRef] [Green Version]
- Hua, C.X.; Li, F.; Zhu, Q.C.; Zhu, G.P.; Meng, L.W. Habitat suitability of Pacific saury (Cololabis saira) based on a yield-density model and weighted analysis. Fish. Res. 2020, 221, 105408. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, H.; Han, H.B.; Zhao, G.Q.; Xu, B.; Shi, Y.C.; Yan, Y.Z.; Ge, Y.L. Spatial and temporal distribution and optimum environmental characteristics of the Sardinops sagax in the North Pacific Ocean. Progr. Fish. Sci. 2022. Available online: http://journal.yykxjz.cn/yykxjz/ch/reader/view_abstract.aspx?journal_id=yykxjz&file_no=202203210000001 (accessed on 14 May 2022). (In Chinese with English Abstract).
- Emmett, R.L.; Brodeur, R.D.; Miller, T.W.; Pool, S.S.; Bentley, P.J.; Krutzikowsky, G.K.; McCrae, J. Pacific sardine (Sardinops sagax) abundance, distribution, and ecological relationships in the Pacific Northwest. Cal. Coop. Ocean. Fish. 2005, 46, 122–143. [Google Scholar]
- Nieto, K.; McClatchie, S.; Weber, E.D.; Lennert-Cody, C.E. Effect of mesoscale eddies and streamers on sardine spawning habitat and recruitment success off southern and central California. J. Geophys. Res. Oceans 2014, 119, 6330–6339. [Google Scholar] [CrossRef]
- Yu, W.; Wen, J.; Chen, X.J.; Gong, Y.; Liu, B.L. Trans-Pacific multidecadal changes of habitat patterns of two squid species. Fish. Res. 2021, 233, 105762. [Google Scholar] [CrossRef]
- Zhang, X.; Saitoh, S.I.; Hirawake, T. Predicting potential fishing zones of Japanese common squid (Todarodes pacificus) using remotely sensed images in coastal waters of south-western Hokkaido, Japan. Int. J. Remote Sens. 2016, 1–18. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | June | July | August | September | October | November |
|---|---|---|---|---|---|---|
| Number | 88 | 181 | 81 | 99 | 70 | 80 |
| Environmental Variables | SST | SSH | SSS | Chla |
|---|---|---|---|---|
| Variance inflation factor (VIF) | 1.52 | 1.62 | 3.38 | 2.49 |
| Evaluation Metric | Fail | Bad | Medium | Good | Very Good |
|---|---|---|---|---|---|
| TSS | 0.00–0.40 | 0.40–0.55 | 0.55–0.70 | 0.70–0.85 | 0.85–1.00 |
| Kappa | 0.00–0.40 | 0.40–0.55 | 0.55–0.70 | 0.70–0.85 | 0.85–1.00 |
| AUC | 0.50–0.60 | 0.60–0.70 | 0.70–0.80 | 0.80–0.90 | 0.90–1.00 |
| Month | Evaluation Metric | GLM | GAM | MARS | GBM | CTA | ANN | SRE | FDA | RF | MaxEnt |
|---|---|---|---|---|---|---|---|---|---|---|---|
| June | TSS | 0.812 | 0.786 | 0.794 | 0.792 | 0.737 | 0.851 | 0.681 | 0.770 | 0.791 | 0.856 |
| Kappa | 0.652 | 0.646 | 0.648 | 0.651 | 0.581 | 0.676 | 0.550 | 0.645 | 0.674 | 0.692 | |
| AUC | 0.930 | 0.926 | 0.915 | 0.925 | 0.865 | 0.936 | 0.839 | 0.920 | 0.932 | 0.938 | |
| July | TSS | 0.825 | 0.873 | 0.866 | 0.869 | 0.809 | 0.780 | 0.739 | 0.828 | 0.872 | 0.864 |
| Kappa | 0.796 | 0.855 | 0.845 | 0.866 | 0.803 | 0.729 | 0.713 | 0.817 | 0.876 | 0.844 | |
| AUC | 0.957 | 0.960 | 0.967 | 0.970 | 0.921 | 0.926 | 0.872 | 0.950 | 0.972 | 0.975 | |
| August | TSS | 0.682 | 0.727 | 0.735 | 0.725 | 0.607 | 0.698 | 0.580 | 0.676 | 0.692 | 0.728 |
| Kappa | 0.525 | 0.615 | 0.622 | 0.636 | 0.526 | 0.596 | 0.475 | 0.566 | 0.621 | 0.623 | |
| AUC | 0.856 | 0.889 | 0.906 | 0.907 | 0.830 | 0.873 | 0.737 | 0.865 | 0.887 | 0.910 | |
| September | TSS | 0.705 | 0.696 | 0.722 | 0.726 | 0.596 | 0.704 | 0.521 | 0.667 | 0.704 | 0.712 |
| Kappa | 0.585 | 0.585 | 0.603 | 0.608 | 0.442 | 0.625 | 0.372 | 0.564 | 0.606 | 0.618 | |
| AUC | 0.894 | 0.894 | 0.908 | 0.909 | 0.821 | 0.908 | 0.764 | 0.881 | 0.907 | 0.911 | |
| October | TSS | 0.761 | 0.741 | 0.768 | 0.738 | 0.669 | 0.735 | 0.642 | 0.704 | 0.715 | 0.747 |
| Kappa | 0.602 | 0.605 | 0.611 | 0.604 | 0.455 | 0.601 | 0.456 | 0.526 | 0.547 | 0.615 | |
| AUC | 0.915 | 0.907 | 0.922 | 0.905 | 0.837 | 0.909 | 0.817 | 0.899 | 0.898 | 0.929 | |
| November | TSS | 0.794 | 0.835 | 0.839 | 0.799 | 0.749 | 0.762 | 0.699 | 0.785 | 0.809 | 0.824 |
| Kappa | 0.618 | 0.654 | 0.658 | 0.639 | 0.533 | 0.620 | 0.584 | 0.625 | 0.645 | 0.661 | |
| AUC | 0.930 | 0.939 | 0.943 | 0.936 | 0.875 | 0.917 | 0.852 | 0.931 | 0.937 | 0.946 |
| Month | Evaluation Metric | Ensemble Model | Model Composition |
|---|---|---|---|
| June | TSS | 0.880 | GLM, GAM, MARS, GBM, ANN, FDA, RF, MaxEnt |
| Kappa | 0.718 | ||
| AUC | 0.942 | ||
| July | TSS | 0.886 | GLM, GAM, MARS, GBM, CTA, ANN, FDA, RF, MaxEnt |
| Kappa | 0.881 | ||
| AUC | 0.980 | ||
| August | TSS | 0.748 | MARS, GBM, MaxEnt |
| Kappa | 0.662 | ||
| AUC | 0.929 | ||
| September | TSS | 0.749 | MARS, GBM, ANN, RF, MaxEnt |
| Kappa | 0.661 | ||
| AUC | 0.920 | ||
| October | TSS | 0.771 | GLM, GAM, MARS, GBM, ANN, MaxEnt |
| Kappa | 0.628 | ||
| AUC | 0.938 | ||
| November | TSS | 0.846 | GLM, GAM, MARS, GBM, CTA, ANN, FDA, RF, MaxEnt |
| Kappa | 0.688 | ||
| AUC | 0.962 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Kang, B.; Fan, W.; Xu, L.; Zhang, S.; Cui, X.; Dai, Y. Spatio-Temporal Variations in the Potential Habitat Distribution of Pacific Sardine (Sardinops sagax) in the Northwest Pacific Ocean. Fishes 2023, 8, 86. https://doi.org/10.3390/fishes8020086
Shi Y, Kang B, Fan W, Xu L, Zhang S, Cui X, Dai Y. Spatio-Temporal Variations in the Potential Habitat Distribution of Pacific Sardine (Sardinops sagax) in the Northwest Pacific Ocean. Fishes. 2023; 8(2):86. https://doi.org/10.3390/fishes8020086
Chicago/Turabian StyleShi, Yongchuang, Bo Kang, Wei Fan, Lingling Xu, Shengmao Zhang, Xuesen Cui, and Yang Dai. 2023. "Spatio-Temporal Variations in the Potential Habitat Distribution of Pacific Sardine (Sardinops sagax) in the Northwest Pacific Ocean" Fishes 8, no. 2: 86. https://doi.org/10.3390/fishes8020086
APA StyleShi, Y., Kang, B., Fan, W., Xu, L., Zhang, S., Cui, X., & Dai, Y. (2023). Spatio-Temporal Variations in the Potential Habitat Distribution of Pacific Sardine (Sardinops sagax) in the Northwest Pacific Ocean. Fishes, 8(2), 86. https://doi.org/10.3390/fishes8020086