
Depth Partitioning and Diel Movement of Two Large Carcharhinid Sharks in Extremely Shallow Waters

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
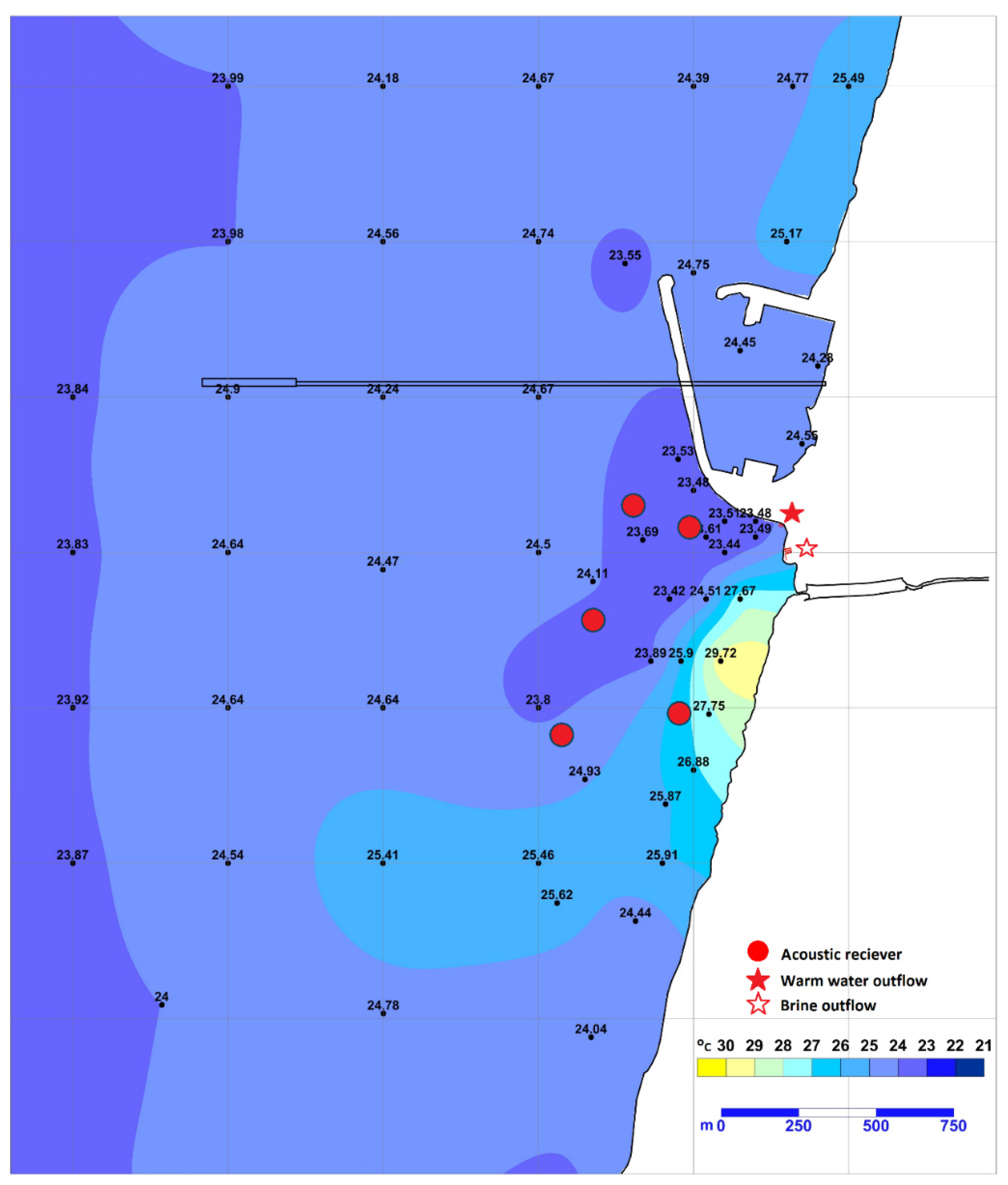
2.1. Study Site
2.2. Shark Tagging and Receivers’ Deployment
2.3. Data Analysis
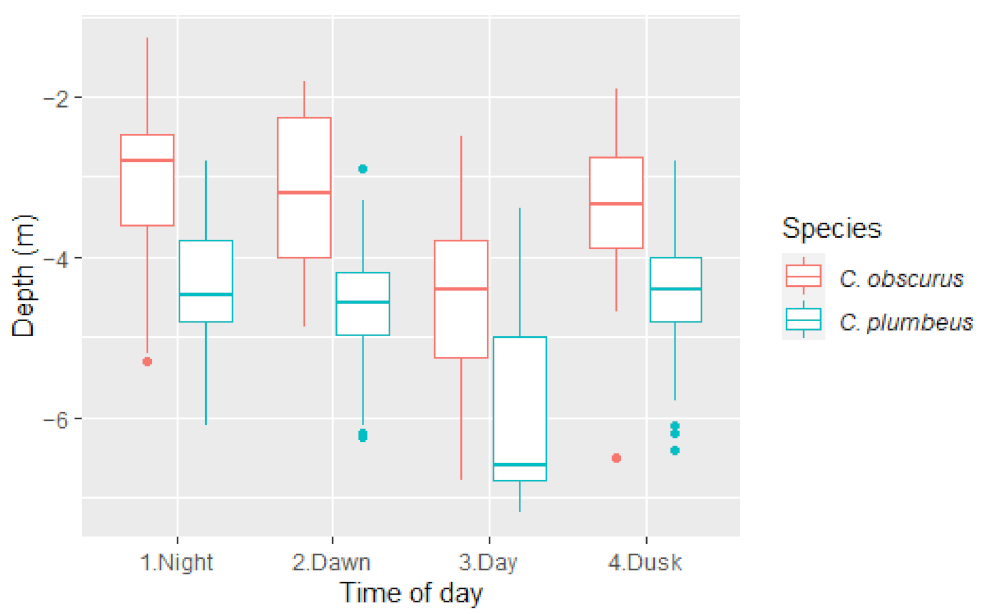
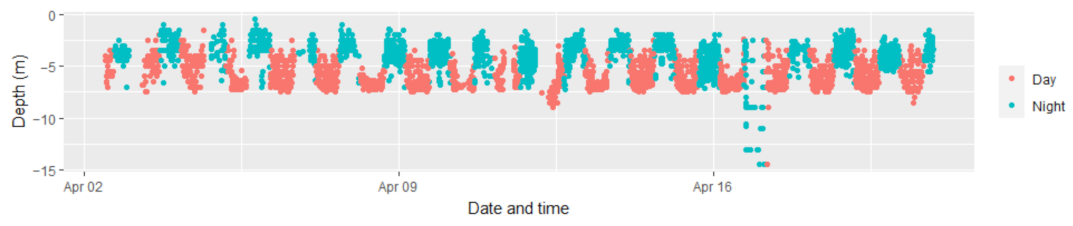
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Catch Date | TL (cm) | Sex |
|---|---|---|---|
| C. obscurus | 25 February 2016 | 322 | Female |
| C. obscurus | 25 February 2016 | 328 | Female |
| C. obscurus | 23 March 2016 | 309 | Female |
| C. obscurus | 23 March 2016 | 325 | Female |
| C. obscurus | 23 March 2016 | 299 | Female |
| C. obscurus | 17 January 2017 | 200 | Female |
| C. obscurus | 20 February 2017 | 250 | Female |
| C. obscurus | 21 February 2017 | 290 | Female |
| C. obscurus | 23 February 2017 | 280 | Female |
| C. obscurus | 6 March 2017 | 280 | Female |
| C. obscurus | 8 March 2017 | 390 | Female |
| C. obscurus | 28 March 2017 | 320 | Female |
| C. obscurus | 19 December 2017 | 283 | Female |
| C. obscurus | 9 January 2018 | 303 | Female |
| C. plumbeus | 8 March 2017 | 170 | Male |
| C. plumbeus | 23 February 2017 | 177 | Male |
| C. plumbeus | 6 April 2017 | 198 | Male |
| C. plumbeus | 6 April 2017 | 179 | Male |
| C. plumbeus | 1 May 2018 | 178 | Male |

References
- Barash, A.; Pickholtz, R.; Pickholtz, E.; Blaustein, L.; Rilov, G. Seasonal Aggregations of Sharks near Coastal Power Plants in Israel: An Emerging Phenomenon. Mar. Ecol. Prog. Ser. 2018, 590, 145–154. [Google Scholar] [CrossRef]
- Weigmann, S. Annotated Checklist of the Living Sharks, Batoids and Chimaeras (Chondrichthyes) of the World, with a Focus on Biogeographical Diversity. J. Fish Biol. 2016, 88, 837–1037. [Google Scholar] [CrossRef] [PubMed]
- Cortés, E. Standardized Diet Compositions and Trophic Levels of Sharks. ICES J. Mar. Sci. 1999, 56, 707–717. [Google Scholar] [CrossRef]
- Finke, D.L.; Snyder, W.E. Niche Partitioning Increases Resource Exploitation by Diverse Communities. Science 2008, 321, 1488–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwyer, R.G.; Campbell, H.A.; Cramp, R.L.; Burke, C.L.; Micheli-Campbell, M.A.; Pillans, R.D.; Lyon, B.J.; Franklin, C.E. Niche Partitioning between River Shark Species Is Driven by Seasonal Fluctuations in Environmental Salinity. Funct. Ecol. 2020, 34, 2170–2185. [Google Scholar] [CrossRef]
- Klimley, A.P.; Ketchum, J.T.; Lara-Lizardi, F.; Papastamatiou, Y.P.; Hoyos-Padilla, E.M. Evidence for Spatial and Temporal Resource Partitioning of Sharks at Roca Partida, an Isolated Pinnacle in the Eastern Pacific. Environ. Biol. Fishes 2022, 105, 1963–1974. [Google Scholar] [CrossRef]
- Lear, K.O.; Whitney, N.M.; Morris, J.J.; Gleiss, A.C. Temporal Niche Partitioning as a Novel Mechanism Promoting Co-Existence of Sympatric Predators in Marine Systems. Proc. R. Soc. B 2021, 288, 20210816. [Google Scholar] [CrossRef] [PubMed]
- Besnard, L.; le Croizier, G.; Galván-Magaña, F.; Point, D.; Kraffe, E.; Ketchum, J.; Rincon, R.O.M.; Schaal, G. Foraging Depth Depicts Resource Partitioning and Contamination Level in a Pelagic Shark Assemblage: Insights from Mercury Stable Isotopes. Environ. Pollut. 2021, 283, 117066. [Google Scholar] [CrossRef] [PubMed]
- Whitney, N.M.; Lear, K.O.; Gleiss, A.C.; Payne, N.; White, C.F. Advances in the Application of High-Resolution Biologgers to Elasmobranch Fishes. In Shark Research: Emerging Technologies and Applications for the Field and Laboratory; Routledge: London, UK, 2018; pp. 45–70. [Google Scholar]
- Thieurmel, B.; Elmarhraoui, A.; Thieurmel, M.B. Package ‘Suncalc’. 2019. Available online: https://cran.r-project.org/web/packages/suncalc/suncalc.pdf (accessed on 8 January 2021).
- Papastamatiou, Y.P.; Wetherbee, B.M.; Lowe, C.G.; Crow, G.L. Distribution and Diet of Four Species of Carcharhinid Shark in the Hawaiian Islands: Evidence for Resource Partitioning and Competitive Exclusion. Mar. Ecol. Prog. Ser. 2006, 320, 239–251. [Google Scholar] [CrossRef]
- Gelsleichter, J.; Musick, J.A.; Nichols, S. Food Habits of the Smooth Dogfish, Mustelus canis, Dusky Shark, Carcharhinus obscurus, Atlantic Sharpnose Shark, Rhizoprionodon terraenovae, and the Sand Tiger, Carcharias taurus, from the Northwest Atlantic Ocean. Environ. Biol. Fishes 1999, 54, 205–217. [Google Scholar] [CrossRef]
- Simpfendorfer, C.A.; Goodreid, A.; McAuley, R.B. Diet of Three Commercially Important Shark Species from Western Australian Waters. Mar. Freshw. Res. 2001, 52, 975–985. [Google Scholar] [CrossRef]
- Smale, M.J. Occurrence and Feeding of Three Shark Species, Carcharhinus brachyurus, C. obscurus and Sphyrna zygaena, on the Eastern Cape Coast of South Africa. South. Afr. J. Mar. Sci. 1991, 11, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Saïdi, B.; Bradaï, M.N.; Bouaïn, A.; Capapé, C. Feeding Habits of the Sandbar Shark Carcharhinus plumbeus (Chondrichthyes: Carcharhinidae) from the Gulf of Gabès, Tunisia. Cah. Biol. Mar. 2007, 48, 139–144. [Google Scholar]
- Vögler, R.; Milessi, A.C.; Duarte, L.O. Changes in Trophic Level of Squatina guggenheim with Increasing Body Length: Relationships with Type, Size and Trophic Level of Its Prey. Environ. Biol. Fishes 2009, 84, 41–52. [Google Scholar] [CrossRef]
- Malpica-Cruz, L.; Herzka, S.Z.; Sosa-Nishizaki, O.; Escobedo-Olvera, M.A. Tissue-specific Stable Isotope Ratios of Shortfin Mako (Isurus oxyrinchus) and White (Carcharodon carcharias) Sharks as Indicators of Size-based Differences in Foraging Habitat and Trophic Level. Fish Oceanogr. 2013, 22, 429–445. [Google Scholar] [CrossRef]
- Romanuk, T.N.; Hayward, A.; Hutchings, J.A. Trophic Level Scales Positively with Body Size in Fishes. Glob. Ecol. Biogeogr. 2011, 20, 231–240. [Google Scholar] [CrossRef]
- Andrews, K.S.; Williams, G.D.; Farrer, D.; Tolimieri, N.; Harvey, C.J.; Bargmann, G.; Levin, P.S. Diel Activity Patterns of Sixgill Sharks, Hexanchus griseus: The Ups and Downs of an Apex Predator. Anim. Behav. 2009, 78, 525–536. [Google Scholar] [CrossRef]



| Shark Serial | Species | Sex | TL (cm) | Detections | Tagging Date | Catch Time | Last Detected | Min Depth (m) | Max Depth (m) | Days Tracked | Detection Rate |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CO 21 | C. obscurus | F | 289 | 318 | 27 November 2017 | 10:30 | 11 March 2018 | 1 | 6.8 | 105 | 3.2 |
| CO 23 | C. obscurus | F | 276 | 737 | 12 December 2017 | 14:34 | 24 April 2018 | 1 | 15 | 134 | 6.3 |
| CO 22 | C. obscurus | F | 315 | 482 | 27 December 2017 | 7:17 | 2 April 2018 | 1 | 7 | 97 | 6.1 |
| CO 14 | C. obscurus | F | 355 | 424 | 27 December 2017 | 10:43 | 13 March 2018 | 1 | 7.4 | 77 | 7.4 |
| CO 20 | C. obscurus | F | 300 | 267 | 2 January 2018 | 13:00 | 8 May 2018 | 0 | 9.2 | 127 | 3.1 |
| CO 26 | C. obscurus | F | 275 | 1051 | 5 February 2018 | NA | 22 April 2018 | 1 | 17.6 | 77 | 8.2 |
| CP 15 | C. plumbeus | M | 169 | 17117 | 12 March 2018 | 13:00 | 14 May 2018 | 0.6 | 13.8 | 64 | 53.5 |
| CO 25 | C. obscurus | F | 280 | 63 | 12 March 2018 | 13:00 | 23 March 2018 | 1.6 | 7.4 | 12 | 1.1 |
| CP 10 | C. plumbeus | M | 191 | 17231 | 14 March 2018 | 10:55 | 10 May 2018 | 0.6 | 17 | 58 | 59.4 |
| CO 11 | C. obscurus | F | 294 | 969 | 28 March 2018 | 8:52 | 27 April 2018 | 1 | 38.6 | 31 | 6.3 |
| CP 17 | C. plumbeus | M | 180 | 4706 | 28 March 2018 | 11:59 | 14 May 2018 | 0 | 11.2 | 48 | 19.6 |
| CO 12 | C. obscurus | F | 300 | 1895 | 2 April 2018 | 11:33 | 2 June 2018 | 0.8 | 7.6 | 62 | 6.1 |
| CP 27 | C. plumbeus | M | 180 | 4348 | 2 April 2018 | 13:49 | 21 April 2018 | 1.4 | 10.8 | 20 | 43.5 |
| Model Formula | AICc | ∆AICc | df | Log Likelihood | adjR2 |
|---|---|---|---|---|---|
| DM ~ Species + TOD + (1|Shark) | 1562.2 | 0 | 7 | −774.010 | 0.579 |
| DM ~ Species + TOD + DIS + (1|Shark) | 1563.1 | 0.9 | 8 | −773.453 | 0.583 |
| DM ~ Species × TOD + (1|Shark) | 1564.1 | 1.9 | 10 | −771.883 | 0.585 |
| Species | Test | Difference (m) | 95 CL | Sig |
|---|---|---|---|---|
| C. obscurus | Dawn (n = 21)—Night (n = 95) | 0.0689 | [−0.382; 0.549] | − |
| C. obscurus | Day (n = 87)—Night (n = 95) | 1.39 | [1.05; 1.73] | + |
| C. obscurus | Dusk (n = 24)—Night (n = 95) | 0.312 | [−0.152; 0.774] | − |
| C. plumbeus | Dawn (n = 66)—Night (n = 125) | 0.351 | [0.0516; 0.639] | + |
| C. plumbeus | Day (n = 116)—Night (n = 125) | 2 | [1.67; 2.34] | + |
| C. plumbeus | Dusk (n = 82)—Night (n = 125) | 0.166 | [−0.122; 0.434] | − |
| TOD | Test | Difference (m) | 95 CL | Sig |
| Night | C. plumbeus (n = 125)—C. obscurus (n = 95) | 1.48 | [1.11; 1.84] | + |
| Dawn | C. plumbeus (n = 66)—C. obscurus (n = 21) | 1.80 | [1.15; 2.42] | + |
| Day | C. plumbeus (n = 116)—C. obscurus (n = 87) | 1.51 | [1.19; 1.85] | + |
| Dusk | C. plumbeus (n = 82)—C. obscurus (n = 24) | 1.26 | [0.556; 1.87] | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barash, A.; Scheinin, A.; Bigal, E.; Zemah Shamir, Z.; Martinez, S.; Tchernov, D. Depth Partitioning and Diel Movement of Two Large Carcharhinid Sharks in Extremely Shallow Waters. Fishes 2023, 8, 85. https://doi.org/10.3390/fishes8020085
Barash A, Scheinin A, Bigal E, Zemah Shamir Z, Martinez S, Tchernov D. Depth Partitioning and Diel Movement of Two Large Carcharhinid Sharks in Extremely Shallow Waters. Fishes. 2023; 8(2):85. https://doi.org/10.3390/fishes8020085
Chicago/Turabian StyleBarash, Adi, Aviad Scheinin, Eyal Bigal, Ziv Zemah Shamir, Stephane Martinez, and Dan Tchernov. 2023. "Depth Partitioning and Diel Movement of Two Large Carcharhinid Sharks in Extremely Shallow Waters" Fishes 8, no. 2: 85. https://doi.org/10.3390/fishes8020085
APA StyleBarash, A., Scheinin, A., Bigal, E., Zemah Shamir, Z., Martinez, S., & Tchernov, D. (2023). Depth Partitioning and Diel Movement of Two Large Carcharhinid Sharks in Extremely Shallow Waters. Fishes, 8(2), 85. https://doi.org/10.3390/fishes8020085