
The Effect of Artificial Substrate and Carbon Source Addition on Bacterial Diversity and Community Composition in Water in a Pond Polyculture System

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites and Design
2.2. Experimental Fish and Culture Management
2.3. Water Quality Indicators
2.4. Bacterial DNA Extraction and 16S rRNA Sequencing
2.5. Statistical Analysis
3. Results
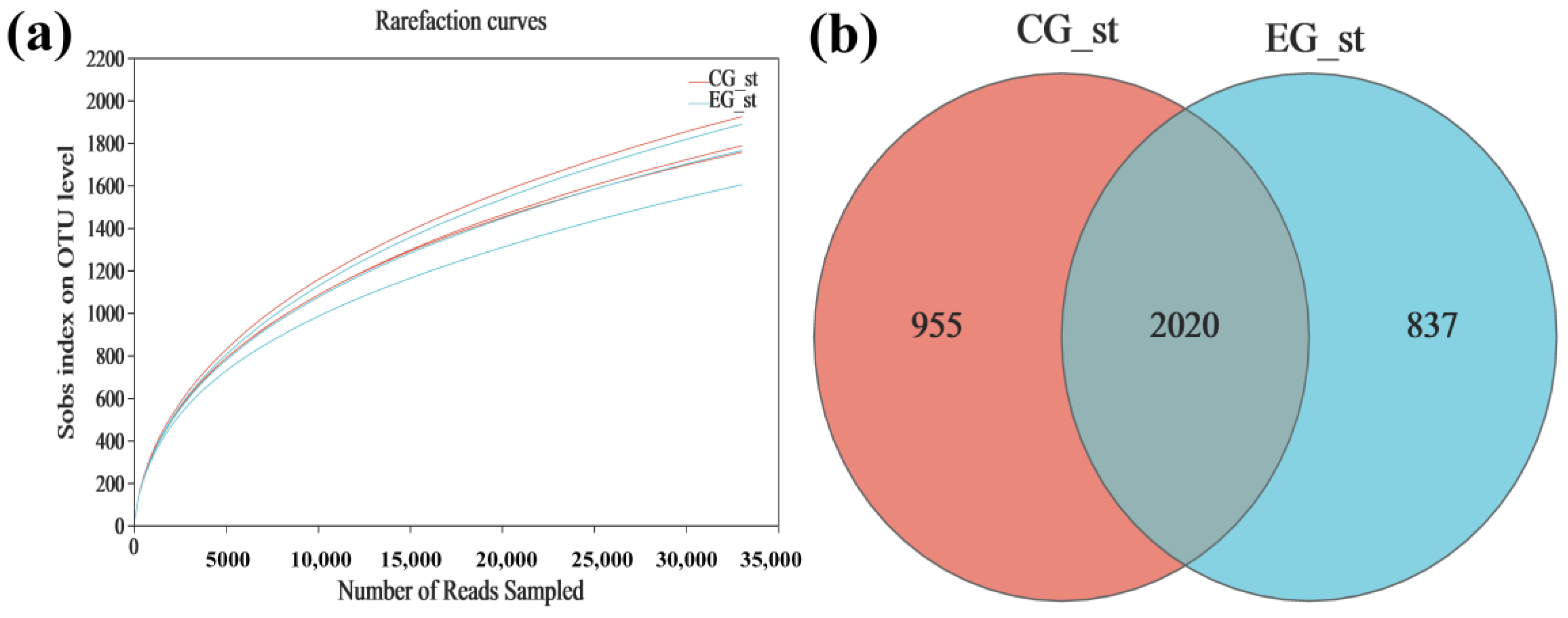
3.1. Bacterial 16S rRNA Sequencing Data and OTUs
3.2. Analysis of Alpha Diversity and Beta Diversity
3.3. Bacterial Community Composition
3.4. Taxonomic Composition Analysis
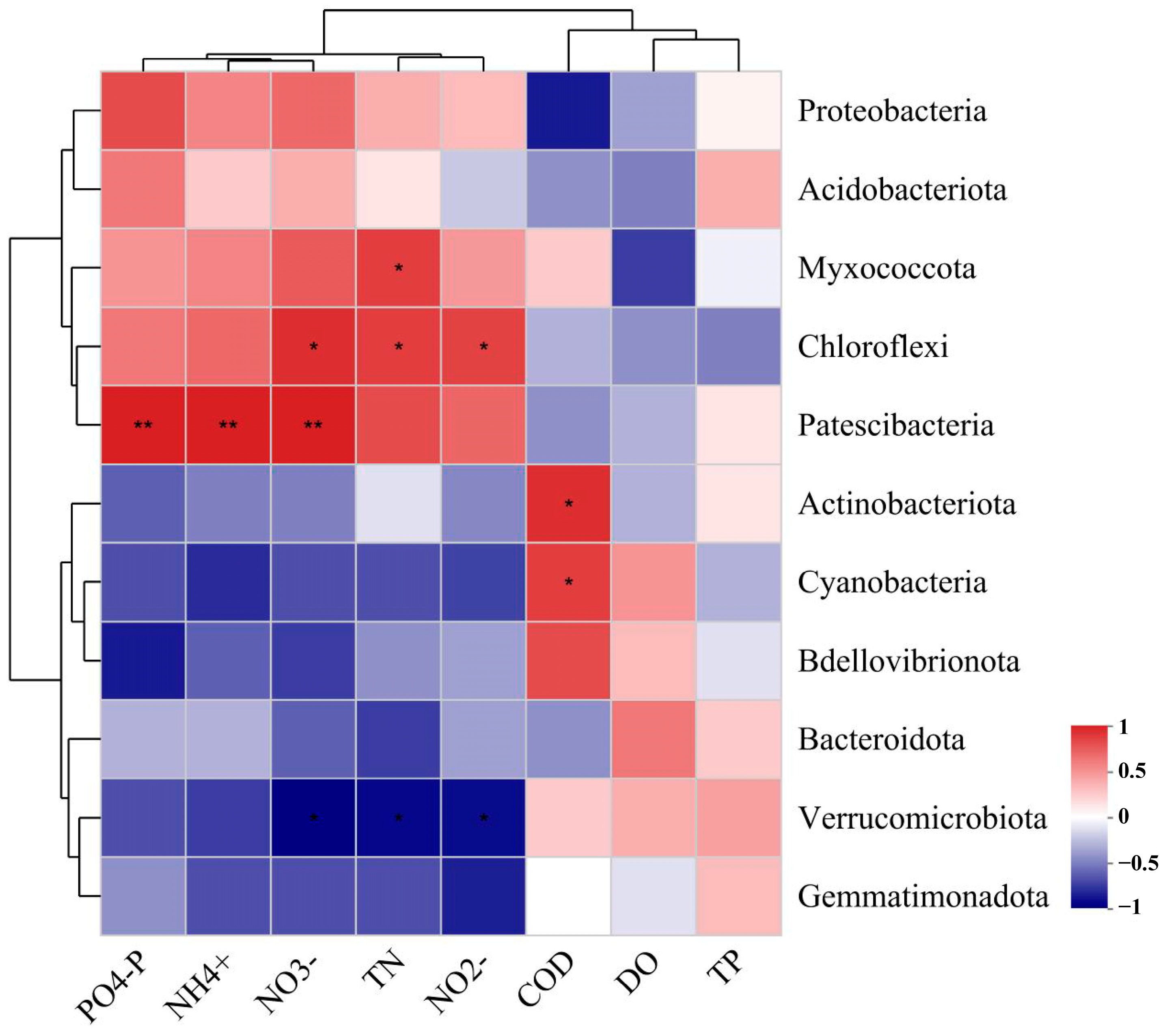
3.5. Correlation Analysis of Water Quality and Microbial Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parkos, J.; Wahl, D. Effects of Common Carp (Cyprinus carpio), an Exotic Fish, on Aquatic Ecosystems; Ilinois Natural History Survey Report of January/February 2000. University of Illinois Board of Trustees; Center for Aquatic Ecology: Raleigh, NC, USA, 2014. [Google Scholar]
- Fisheries and Fisheries Administration of the Ministry of Agriculture and Rural Affairs of China. China Fisheries Statistcal Yearbook; China Agriculture Press: Beijing, China, 2022. (In Chinese) [Google Scholar]
- Zhao, Y.J.; Zhang, J.; Qiao, D.; Gao, F.; Gu, Y.L.; Jiang, X.Y.; Zhu, L.; Kong, X.H. CcGSDMEa functions the pore-formation in cytomembrane and the regulation on the secretion of IL-lb in common carp (Cyprinus carpio haematopterus). Front. Immunol. 2023, 13, 1110322. [Google Scholar] [CrossRef]
- Zhu, S.-M.; Deng, Y.-L.; Ruan, Y.-J.; Guo, X.-S.; Shi, M.-M.; Shen, J.-Z. Biological denitrification using poly(butylene succinate) as carbon source and biofilm carrier for recirculating aquaculture system effluent treatment. Bioresour. Technol. 2015, 192, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Barraza-Guardado, R.H.; Arreola-Lizárraga, J.A.; López-Torres, M.A.; Casillas-Hernández, R.; Miranda-Baeza, A.; Magallón-Barrajas, F.; Ibarra-Gámez, C. Effluents of shrimp farms and its influence on the coastal ecosystems of Bahía de Kino, Mexico. Sci. World J. 2013, 2013, 306370. [Google Scholar] [CrossRef]
- Notice of the General Office of the Ministry of Agriculture and Rural Affairs on the Implementing the “Five Major Actions” of Green and Healthy Aquaculture in 2020; Gazette of the Ministry of Agriculture and Rural Affairs of the People’s Republic of China: Beijing, China, 2020; pp. 86–87.
- Yu, Y.-B.; Choi, J.-H.; Lee, J.-H.; Jo, A.-H.; Lee, K.M.; Kim, J.-H. Biofloc Technology in Fish Aquaculture: A Review. Antioxidants 2023, 12, 398. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.F.; Pinto, R.C.; Nunes, A.J. Nutrient value and contribution of microbial floc to the growth performance of juvenile shrimp, Litopenaeus vannamei, fed fatty acid and amino acid-restrained diets under a zero-water exchange intensive system. Aquaculture 2021, 531, 735789. [Google Scholar] [CrossRef]
- Wei, Y.; Liao, S.-A.; Wang, A.-L. The effect of different carbon sources on the nutritional composition, microbial community and structure of bioflocs. Aquaculture 2016, 465, 88–93. [Google Scholar] [CrossRef]
- Uddin, M.; Azim, M.; Wahab, M.; Verdegem, M. Effects of substrate addition and supplemental feeding on plankton composition and production in tilapia (Oreochromis niloticus) and freshwater prawn (Macrobrachium rosenbergii) polyculture. Aquaculture 2010, 297, 99–105. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Rahman, M.; Azim, M.; Islam, M.A.; Wahab, M.; Verdegem, M.; Verreth, J. Effects of C/N ratio and substrate addition on natural food communities in freshwater prawn monoculture ponds. Aquaculture 2010, 306, 127–136. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Wahab, M.; Verdegem, M.; Benerjee, S.; Akter, T.; Hasan, M.; Azim, M. Effects of addition of tilapia Oreochromis niloticus, and substrates for periphyton developments on pond ecology and production in C/N-controlled freshwater prawn Macrobrachium rosenbergii, farming systems. Aquaculture 2009, 287, 371–380. [Google Scholar] [CrossRef]
- Huggett, M.J.; Nedved, B.T.; Hadfield, M.G. Effects of initial surface wettability on biofilm formation and subsequent settlement of Hydroides elegans. Biofouling 2009, 25, 387–399. [Google Scholar] [CrossRef]
- Schveitzer, R.; Arantes, R.; Baloi, M.F.; Costódio, P.F.S.; Arana, L.V.; Seiffert, W.Q.; Andreatta, E.R. Use of artificial substrates in the culture of Litopenaeus vannamei (Biofloc System) at different stocking densities: Effects on microbial activity, water quality and production rates. Aquac. Eng. 2013, 54, 93–103. [Google Scholar] [CrossRef]
- Ballester, E.L.C.; Wasielesky, W.; Cavalli, R.O.; Abreu, P.C. Nursery of the pink shrimp Farfantepenaeus paulensis in cages with artificial substrates: Biofilm composition and shrimp performance. Aquaculture 2007, 269, 355–362. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Wahab, M.; Verdegem, M.; Mondal, M.; Azim, M. Effects of stocking density of freshwater prawn Macrobrachium rosenbergii and addition of different levels of tilapia Oreochromis niloticus on production in C/N controlled periphyton based system. Aquaculture 2009, 286, 72–79. [Google Scholar] [CrossRef]
- AftabUddin, S.; Siddique, M.A.M.; Sein, A.; Dey, P.K.; Rashed-Un-Nabi, M.; Haque, M.A. First use of biofloc technology for Penaeus monodon culture in Bangladesh: Effects of stocking density on growth performance of shrimp, water quality and bacterial growth. Aquac. Rep. 2020, 18, 100518. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Wahab, M.A.; Verdegem, M.C.J.; Azim, M.E.; Haque, S.; Salam, M.A. C/N ratio control and substrate addition for periphyton development jointly enhance freshwater prawn Macrobrachium rosenbergii production in ponds. Aquaculture 2008, 280, 117–123. [Google Scholar] [CrossRef]
- Guo, K.; Zhao, Z.; Xie, J.; Luo, L.; Wang, S.; Zhang, R.; Xu, W.; Huang, X. Combined effects of eco-substrate and carbon addition on water quality, fish performance and nutrient budgets in the pond polyculture system. Fishes 2022, 7, 212. [Google Scholar] [CrossRef]
- Song, C.; Zhong, L.; Liu, Y.; Yin, Y.; Zhang, C.; Zhang, X.; Chen, J. Spatial and temporal succession of bacterial communities in three artificial fishponds. Aquac. Res. 2019, 50, 2793–2801. [Google Scholar] [CrossRef]
- Mohapatra, S.; Chakraborty, T.; Kumar, V.; DeBoeck, G.; Mohanta, K.N. Aquaculture and stress management: A review of probiotic intervention. J. Anim. Physiol. Anim. Nutr. 2013, 97, 405–430. [Google Scholar] [CrossRef]
- Ray, A.J.; Lotz, J.M. Shrimp (Litopenaeus vannamei) production and stable isotope dynamics in clear-water recirculating aquaculture systems versus biofloc systems. Aquac. Res. 2017, 48, 4390–4398. [Google Scholar] [CrossRef]
- Ebeling, J.M.; Timmons, M.B. Carbon: Nitrogen balance impacts nitrogen removal process in microbial-based aquaculture systems. Glob. Aquac. Advocate 2008, 11, 38–40. [Google Scholar]
- Singh, T.; Mandal, A.; Holeyappa, S.A.; Khairnar, S.O.; Tyagi, A. Growth performance and physiological responses of striped catfish (Pangasianodon hypophthalmus) under different carbohydrates supplemented biofloc aquaculture systems. Aquaculture 2024, 579, 740252. [Google Scholar] [CrossRef]
- Hu, X.; Meng, L.-J.; Liu, H.-D.; Guo, Y.-S.; Liu, W.-C.; Tan, H.-X.; Luo, G.-Z. Impacts of Nile Tilapia (Oreochromis niloticus) exposed to microplastics in bioflocs system. Sci. Total Environ. 2023, 901, 165921. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Xu, J.; Meng, H. Nitrate accumulation in biofloc aquaculture systems. Aquaculture 2020, 520, 734675. [Google Scholar] [CrossRef]
- Morais, A.P.M.; Abreu, P.C.; Wasielesky, W., Jr.; Krummenauer, D. Effect of aeration intensity on the biofilm nitrification process during the production of the white shrimp Litopenaeus vannamei (Boone, 1931) in Biofloc and clear water systems. Aquaculture 2020, 514, 734516. [Google Scholar] [CrossRef]
- Keshavanath, P.; Gangadhar, B.; Ramesh, T.J.; Van Rooij, J.M.; Beveridge, M.C.M.; Baird, D.J.; Verdegem, M.C.J.; Van Dam, A.A. Use of artificial substrates to enhance production of freshwater herbivorous fish in pond culture. Aquac. Res. 2001, 32, 189–197. [Google Scholar] [CrossRef]
- Carlos, L.C.; Paniagua-Michel, J. Effects of constructed microbial mats on water quality and performance of Litopenaeus vannamei, post-larvae. Aquac. Eng. 2010, 42, 75–81. [Google Scholar]
- Houdt, R.V.; Michiels, C.W. Role of bacterial cell surface structures in Escherichia coli, biofilm formation. Res. Microbiol. 2005, 156, 626–633. [Google Scholar] [CrossRef]
- Gullian-Klanian, M.; Quintanilla-Mena, M.; Hau, C.P. Influence of the biofloc bacterial community on the digestive activity of Nile tilapia (Oreochromis niloticus). Aquaculture 2023, 562, 738774. [Google Scholar] [CrossRef]
- Bratvold, D.; Browdy, C.L. Effects of sand sediment and vertical surfaces (AquaMatsTM) on production, water quality, and microbial ecology in an intensive Litopevaeus vamvamei culture system. Aquaculture 2001, 195, 81–94. [Google Scholar] [CrossRef]
- Ray, A.J.; Seaborn, G.; Leffler, J.W.; Wilde, S.B.; Lawson, A.; Browdy, C.L. Characterization of microbial communities in minimal-exchange, intensive aquaculture systems and the effects of suspended solids management. Aquaculture 2010, 310, 130–138. [Google Scholar] [CrossRef]
- Cardona, E.; Gueguen, Y.; Magré, K.; Lorgeoux, B.; Piquemal, D.; Pierrat, F.; Noguier, F.; Saulnier, D. Bacterial community characterization of water and intestine of the shrimp Litopenaeus stylirostris in a biofloc system. BMC Microbiol. 2016, 16, 157. [Google Scholar] [CrossRef]
- Kruglova, A.; Gonzalez-Martinez, A.; Kråkström, M.; Mikola, A.; Vahala, R. Bacterial diversity and population shifts driven by spotlight wastewater micropollutants in low-temperature highly nitrifying activated sludge. Sci. Total Environ. 2017, 605–606, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, Y.; Zhao, L.; Li, Y.; Xie, S.; Liu, Y. Distribution of sediment bacterial and archaeal communities in plateau freshwater lakes. Appl. Microbiol. Biotechnol. 2015, 99, 3291–3302. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Barry, K.; Hu, G.; Meng, S.; Song, C.; Qiu, L.; Zheng, Y.; Wu, W.; Qu, J.; Chen, J.; et al. Characterizing bacterial communities in tilapia pond surface sediment and their responses to pond differences and temporal variations. World J. Microbiol. Biotechnol. 2017, 33, 1. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.-K.; Sun, Q.; Fang, P.-H.; Li, X.-Q.; Zeng, R.; Wang, H.-C.; Wang, A.-J. Integrated constructed wetland and bioelectrochemistry system approach for simultaneous enhancment of p-chloronitrobenzene and nitrogen transformations performance. Water Res. 2022, 217, 118433. [Google Scholar] [CrossRef] [PubMed]
- Klase, G.; Lee, S.; Liang, S.; Kim, J.; Zo, Y.-G.; Lee, J. The microbiome and antibiotic resistance in integrated fishfarm water: Implications of environmental public health. Sci. Total Environ. 2019, 649, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Paniagua-Michel, J. Wastewater treatment using phototrophic–heterotrophic biofilms and microbial mats. In Prospects and Challenges in Algal Biotechnology; Tripathi, B.N., Kumar, D., Eds.; Springer: Singapore, 2017; pp. 257–275. [Google Scholar]
- Lv, Y.; Wan, C.; Lee, D.-J.; Tay, J.-H. Microbial communities of aerobic granules: Granulation mechanisms. Bioresour. Technol. 2014, 169, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhao, L.; Cheng, Y.; Yang, Y.; Sun, Y.; Liu, Q. Control of cyanobacterial blooms in different polyculture patterns of filter feeders and effects of these patterns on water quality and microbial community in aquacultural ponds. Aquaculture 2021, 542, 736913. [Google Scholar] [CrossRef]
- Kiersztyn, B.; Chróst, R.; Kalinski, T.; Siuda, W.; Bukowska, A.; Kowalczyk, G.; Grabowska, K. Structural and functional microbial diversity along a eutrophication gradient of interconnected lakes undergoing anthropopressure. Sci. Rep. 2019, 9, 111444. [Google Scholar] [CrossRef]
- Das, S.; Ward, L.R.; Burke, C. Prospects of using marine actinobacteria as probiotics in aquaculture. Appl. Microbiol. Biotechnol. 2008, 81, 419–429. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Coverage | Richness | Diversity | ||
|---|---|---|---|---|---|
| ACE | Chao | Shannon | Simpson | ||
| CGˍst | 0.979 ± 0.001 | 3055 ± 389 | 2720 ± 172 | 5.455 ± 0.048 | 0.0170 ± 0.009 |
| EGˍst | 0.978 ± 0.001 | 3247 ± 90 | 2678 ± 154 | 5.318 ± 0.076 | 0.0266 ± 0.0070 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, K.; Shi, M.; Huang, X.; Luo, L.; Wang, S.; Zhang, R.; Xu, W.; Ruan, G.; Zhao, Z. The Effect of Artificial Substrate and Carbon Source Addition on Bacterial Diversity and Community Composition in Water in a Pond Polyculture System. Fishes 2024, 9, 80. https://doi.org/10.3390/fishes9030080
Guo K, Shi M, Huang X, Luo L, Wang S, Zhang R, Xu W, Ruan G, Zhao Z. The Effect of Artificial Substrate and Carbon Source Addition on Bacterial Diversity and Community Composition in Water in a Pond Polyculture System. Fishes. 2024; 9(3):80. https://doi.org/10.3390/fishes9030080
Chicago/Turabian StyleGuo, Kun, Mumin Shi, Xiaoli Huang, Liang Luo, Shihui Wang, Rui Zhang, Wei Xu, Guoliang Ruan, and Zhigang Zhao. 2024. "The Effect of Artificial Substrate and Carbon Source Addition on Bacterial Diversity and Community Composition in Water in a Pond Polyculture System" Fishes 9, no. 3: 80. https://doi.org/10.3390/fishes9030080
APA StyleGuo, K., Shi, M., Huang, X., Luo, L., Wang, S., Zhang, R., Xu, W., Ruan, G., & Zhao, Z. (2024). The Effect of Artificial Substrate and Carbon Source Addition on Bacterial Diversity and Community Composition in Water in a Pond Polyculture System. Fishes, 9(3), 80. https://doi.org/10.3390/fishes9030080