Distribution of Phlebotomine Sandflies in the Cave Area of Satun Province, Thailand

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Environmental Parameter Assessment
2.2. Sandfly Abundance
2.3. Potential Breeding Site Exploration
2.4. Morphological Identification
2.5. Leishmania spp. Detection
2.6. Data Analysis
3. Results
3.1. Sandfly Distribution and Abundance
3.2. Potential Breeding Sites and Environmental Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Apiwathnasorn, C.; Samung, Y.; Prummongkol, S.; Phayakaphon, A.; Panasopolkul, C. Cavernicolous species of phlebotomine sand flies from Kanchanaburi Province, with an updated species list for Thailand. Southeast Asian J. Trop. Med. Public Health 2011, 42, 42. [Google Scholar]
- Kanjanopas, K.; Siripattanapipong, S.; Ninsaeng, U.; Hitakarun, A.; Jitkaew, S.; Kaewtaphaya, P.; Tan-Ariya, P.; Mungthin, M.; Charoenwong, C.; Leelayoova, S. Sergentomyia (Neophlebotomus) gemmea, a potential vector of Leishmania siamensis in southern Thailand. BMC Infect. Dis. 2013, 13, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lainson, R.; Ward, R.D.; Shaw, J.J. Leishmania in phlebotomid sandflies: VI. Importance of hindgut development in distinguishing between parasites of the Leishmania mexicana and L. braziliensis complexes. Proc. R. Soc. Lond. Ser. B Boil. Sci. 1977, 199, 309–320. [Google Scholar] [CrossRef]
- Leelayoova, S.; Siripattanapipong, S.; Manomat, J.; Piyaraj, P.; Tan-Ariya, P.; Bualert, L.; Mungthin, M. Leishmaniasis in Thailand: A Review of Causative Agents and Situations. Am. J. Trop. Med. Hyg. 2017, 96, 534–542. [Google Scholar] [CrossRef] [Green Version]
- Viriyavejakul, P.; Viravan, C.; Riganti, M.; Punpoowong, B. Imported cutaneous leishmaniasis in Thailand. Southeast Asian J. Trop. Med. Public Health 1997, 28, 558–562. [Google Scholar]
- Jaturas, N.; Vitta, A.; Samung, Y.; Apiwathnasorn, C.; Polseela, R. Species composition and nocturnal activity of phlebotomine sand flies (Diptera: Psychodidae) inhabiting a limestone cave in Thailand. J. Vector Ecol. 2018, 43, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Pawestri, A.R.; Thammapalo, S.; Tuekprakhon, A.; Apiwathnasorn, C.; Kumlert, R. Updated directory of sandfly (Subfamily Phlebotominae) species in Thailand. (In preparation)
- Chusri, S.; Thammapalo, S.; Silpapojakul, K.; Siriyasatien, P. Animal reservoirs and potential vectors of Leishmania siamensis in southern Thailand. Southeast Asian J. Trop. Med. Public Health 2014, 45, 13. [Google Scholar]
- Polseela, R.; Vitta, A.; Apiwathnasorn, C. Distribution of phlebotomine sand flies (Diptera:Psychodidae) in Limestone Caves, Khao Pathawi, Uthai Thani Province, Thailand. Southeast Asian J. Trop. Med. Public Health 2015, 46, 425–433. [Google Scholar]
- Volf, P.; Volfova, V. Establishment and maintenance of sand fly colonies. J. Vector Ecol. 2011, 36, S1–S9. [Google Scholar] [CrossRef]
- Alencar, R.B.; De Queiroz, R.G.; Barrett, T.V. Breeding sites of phlebotomine sand flies (Diptera: Psychodidae) and efficiency of extraction techniques for immature stages in terra-firme forest in Amazonas State, Brazil. Acta Trop. 2011, 118, 204–208. [Google Scholar] [CrossRef]
- Müller, P.; Kravchenko, V.D.; Rybalov, L.; Schlein, Y. Characteristics of resting and breeding habitats of adult sand flies in the Judean Desert. J. Vector Ecol. 2011, 36, S195–S205. [Google Scholar] [CrossRef] [PubMed]
- Choocherd, S.; Phasuk, J. Nocturnal Activity of Phlebotomine Sand Flies in Satun Province, Thailand. Southeast Asian J. Trop. Med. Public Health 2018, 49, 383–390. [Google Scholar]
- Panthawong, A.; Chareonviriyaphap, T.; Phasuk, J. Species Diversity and Seasonality of Phlebotomine Sand Flies (Diptera: Psychodidae) in Satun Province, Thailand. Southeast Asian J. Trop. Med. Public Health 2015, 46, 857. [Google Scholar] [PubMed]
- Tourism Authority of Thailand Satun. Available online: https://www.tourismthailand.org/Destinations/Provinces/Satun/352 (accessed on 29 September 2020).
- Open Street Map Project Climate-Data.org, Thailand, Satun Province. Available online: https://en.climate-data.org/asia/thailand/satun-province/satun-30340/ (accessed on 17 September 2020).
- Division of Vector Borne Diseases; DDC (MOPH); Sandfly, T. Leishmaniasis; Division of Vector Borne Diseases; Department of Disease Control, Ministry of Public Health: Nonthaburi, Thailand, 2003; ISBN 9742972206.
- Lewis, D.J. Phlebotomine sandflies (Diptera: Psychodidae) from the Oriental Region. Syst. Èntomol. 1987, 12, 163–180. [Google Scholar] [CrossRef]
- Galati, E.A.B.; Galvis-Ovallos, F.; Lawyer, P.; Léger, N.; Depaquit, J. An illustrated guide for characters and terminology used in descriptions of Phlebotominae (Diptera, Psychodidae). Parasite 2017, 24, 26. [Google Scholar] [CrossRef] [Green Version]
- Rispail, P.; Léger, N. Numerical taxonomy of Old World Phlebotominae (Diptera: Psychodidae). 2. Restatement of classification upon subgeneric morphological characters. Memórias Do Inst. Oswaldo Cruz 1999, 93, 787–793. [Google Scholar] [CrossRef] [Green Version]
- Almazán, M.C.; Copa, G.N.; Lauthier, J.J.; Gil, J.F.; Quiroga, I.L.; Hoyos, C.L.; Fernández, M.E.D.; Nasser, J.R.; Korenaga, M.; Marco, J.D.; et al. Sand fly typing: A simple and morphologically-supported method based on polymorphism of 18S rRNA gene in a Leishmaniasis endemic area of Argentina. Acta Trop. 2020, 211, 105609. [Google Scholar] [CrossRef]
- Oswaldo Cruz. LeishEpiNetSA Manual Molecular Procedures, Training Course Molecular Epidemiology Leishmaniasis; Instituto Oswaldo Cruz: Rio de Janeiro, Brazil, 2009. [Google Scholar]
- Mirahmadi, H.; Khorashad, A.S.; Sohrabnahad, A.; Heydarian, P.; Bizhani, N. Species Identification and Molecular Typing of Leishmania Spp. Using Targeting HSP70 Gene in Suspected Patients of Cutaneous Leishmaniasis from Sistan and Baluchestan Province, Southeast Iran. Iran. J. Parasitol. 2016, 11, 489–498. [Google Scholar]
- Requena, J.M.; Chicharro, C.; García, L.; Parrado, R.; Puerta, C.J.; Cañavate, C. Sequence analysis of the 3′-untranslated region of HSP70 (type I) genes in the genus Leishmania: Its usefulness as a molecular marker for species identification. Parasites Vectors 2012, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Feliciangeli, M.D. Natural breeding places of phlebotomine sandflies. Med. Veter. Èntomol. 2004, 18, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Hanafi, H.A.; Fryauff, D.J.; Modi, G.B.; Ibrahim, M.O.; Main, A.J. Bionomics of phlebotomine sandflies at a peacekeeping duty site in the north of Sinai, Egypt. Acta Trop. 2007, 101, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Veter. Èntomol. 2012, 27, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Phasuk, J.; Inpankaew, T.; Choocherd, S.; Samung, Y. Identification of Mammalian Blood Meals from Sergentomyia (Parrotomyia) barraudi (Diptera: Psychodidae) in Songkhla Province, Southern Thailand. Southeast Asian J. Trop. Med. Public Health 2019, 50, 991–995. [Google Scholar]
- Polseela, R.; Vitta, A.; Nateeworanart, S.; Apiwathnasorn, C. Distribution of cave-dwelling phlebotomine sand flies and their nocturnal and diurnal activity in Phitsanulok Province, Thailand. Southeast Asian J. Trop. Med. Public Health 2011, 42, 42. [Google Scholar]
- Polseela, R.; Apiwathnasorn, C.; Samung, Y. Seasonal distribution of phlebotomine sand flies (Diptera: Psychodidae) in Tham Phra Phothisat temple, Saraburi province, Thailand. Trop. Biomed. 2011, 28, 366–375. [Google Scholar]
- Votýpka, J.; Pavlasova, M.; Volfova, V.; Volf, P.; And, V.V. Rotation of male genitalia in various species of phlebotomine sandfly. Med. Veter. Èntomol. 2015, 29, 355–360. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. The biology and control of Phlebotomine sand flies. Clin. Dermatol. 1999, 17, 279–289. [Google Scholar] [CrossRef]
- Ready, P.D. Biology of Phlebotomine Sand Flies as Vectors of Disease Agents. Annu. Rev. Èntomol. 2013, 58, 227–250. [Google Scholar] [CrossRef]
- Guzmán, H.; Tesh, R.B. Effects of temperature and diet on the growth and longevity of phlebotomine sand flies. Biomedica 2000, 20, 190–199. [Google Scholar] [CrossRef] [Green Version]
- Mellor, P.; Leake, C. Climatic and geographic influences on arboviral infections and vectors. Rev. Sci. Tech. 2000, 19, 41–54. [Google Scholar] [CrossRef]
- Oshaghi, M.A.; Ravasan, N.M.; Javadian, E.; Rassi, Y.; Sadraei, J.; A Enayati, A.; Hassan, V.; Zare, Z.; Emami, S.N. Application of predictive degree day model for field development of sandfly vectors of visceral leishmaniasis in northwest of Iran. J. Vector Borne Dis. 2009, 46, 247–255. [Google Scholar] [PubMed]
- Wijerathna, T.; Gunathilaka, N. Diurnal adult resting sites and breeding habitats of phlebotomine sand flies in cutaneous leishmaniasis endemic areas of Kurunegala District, Sri Lanka. Parasites Vectors 2020, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Vivero, R.J.; Torres-Gutierrez, C.; E Bejarano, E.; Peña, H.C.; Estrada, L.G.; Florez, F.; Ortega, E.; Aparicio, Y.; E Muskus, C. Study on natural breeding sites of sand flies (Diptera: Phlebotominae) in areas of Leishmania transmission in Colombia. Parasites Vectors 2015, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
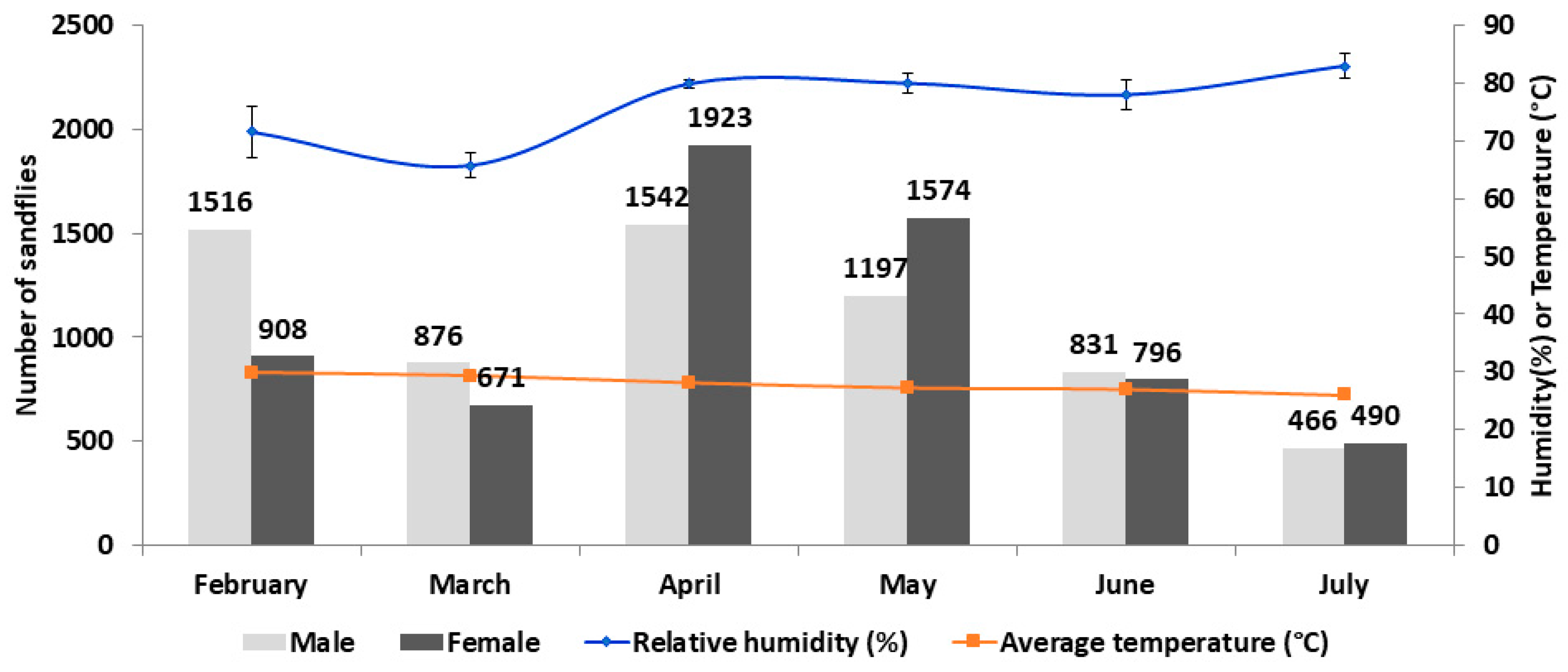
| Month | Relative Humidity (%) (Mean ± SD) | Average Temperature (°C) (Mean ± SD) | Total Number (Male:Female) | Blood Engorged Female | Trap Success 1 | Female Abundance 2 |
|---|---|---|---|---|---|---|
| February | 72 ± 4.51 | 30 ± 0.77 | 2424 (1.7:1) | 12 (1.3%) | 242 | 91 |
| March | 66 ± 2.16 | 29 ± 1.05 | 1547 (1.3:1) | 45 (6.7%) | 155 | 67 |
| April | 80 ± 0.70 | 28 ± 0.64 | 3465 (0.8:1) | 118 (6.2%) | 347 | 192 |
| May | 80 ± 1.61 | 27 ± 0.58 | 2771 (0.8:1) | 128 (8.2%) | 277 | 157 |
| June | 78 ± 2.48 | 27 ± 0.55 | 1627 (1:1) | 49 (6.1%) | 163 | 80 |
| July | 83 ± 2.09 | 26 ± 0.63 | 956 (1:1) | 17 (3.5%) | 96 | 49 |
| Total | 12790 (1:1) | 369 (5.8%) |
| Species | February | March | April | May | June | July | Relative Abundance (%) |
|---|---|---|---|---|---|---|---|
| S. anodontis | 28 | 20 | 19 | 19 | 36 | 38 | 160 (26.8%) |
| S. barraudi | 25 | 3 | 2 | 0 | 1 | 7 | 38 (6.4%) |
| S. sylvatica | 0 | 0 | 0 | 4 | 0 | 1 | 5 (0.8%) |
| Sergentomyia spp. 1 | 38 | 57 | 62 | 59 | 40 | 34 | 290 (48.5%) |
| P. asperulus | 6 | 6 | 5 | 2 | 10 | 6 | 35 (5.9%) |
| P. stantoni | 1 | 4 | 3 | 3 | 0 | 0 | 11 (1.8%) |
| P. betisi | 1 | 0 | 0 | 0 | 0 | 0 | 1 (0.2%) |
| Phlebotomus spp. 2 | 12 | 6 | 7 | 10 | 11 | 12 | 58 (9.7%) |
| 111 | 96 | 98 | 97 | 98 | 98 | 598 |
| Month | Total Number | Numbers of Samples Selected | Unrotated or Partially Rotated Genitalia | Fully-Rotated Genitalia |
|---|---|---|---|---|
| February | 1516 | 53 | 43 (81%) | 10 (19%) |
| March | 876 | 71 | 52 (73%) | 19 (27%) |
| April | 1542 | 95 | 80 (84%) | 15 (16%) |
| May | 1197 | 113 | 80 (71%) | 33 (29%) |
| June | 831 | 111 | 96 (86%) | 15 (14%) |
| July | 466 | 102 | 78 (76%) | 24 (24%) |
| Total | 6828 | 545 | 429 (78.7%) | 116 (21.3%) |
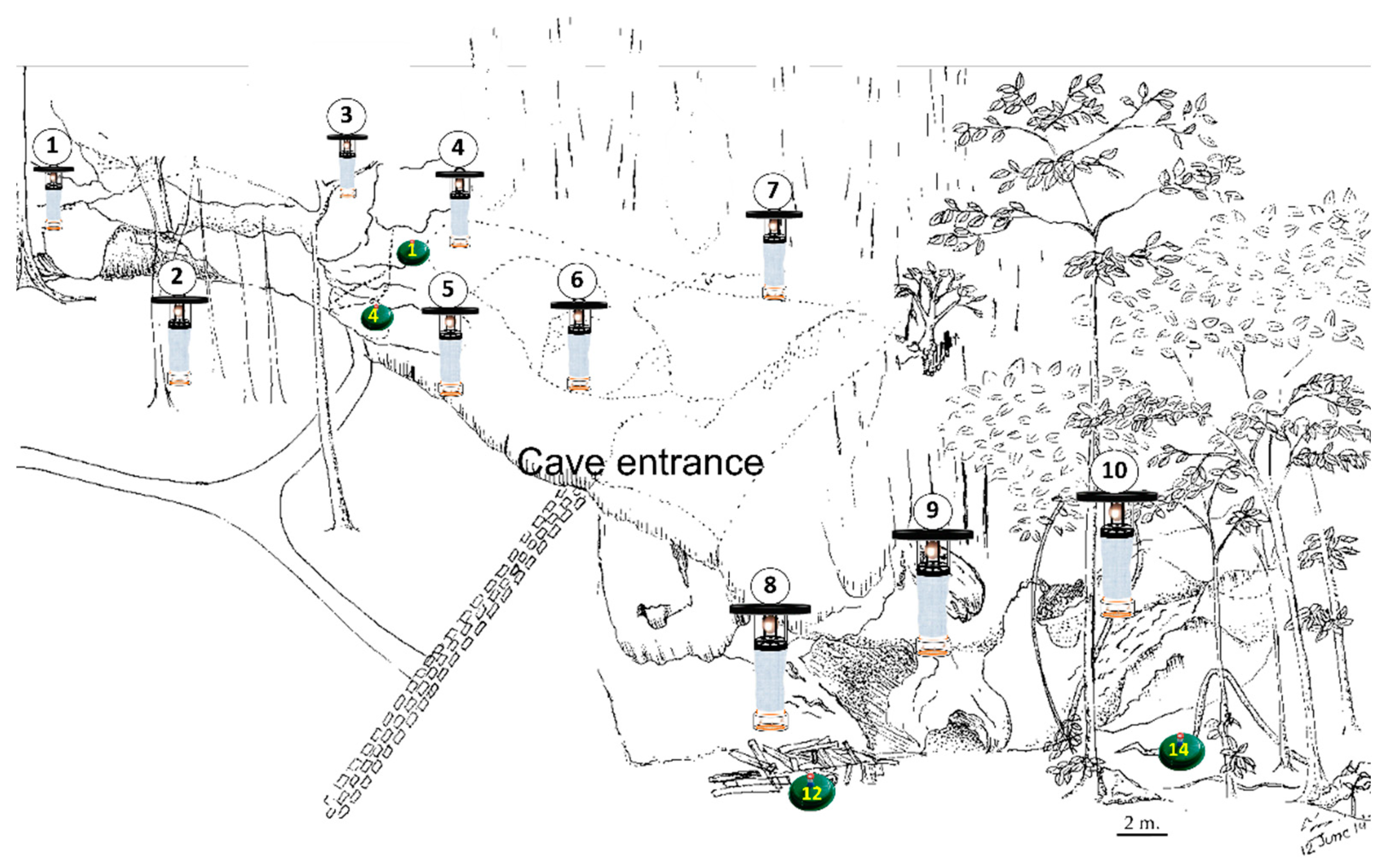
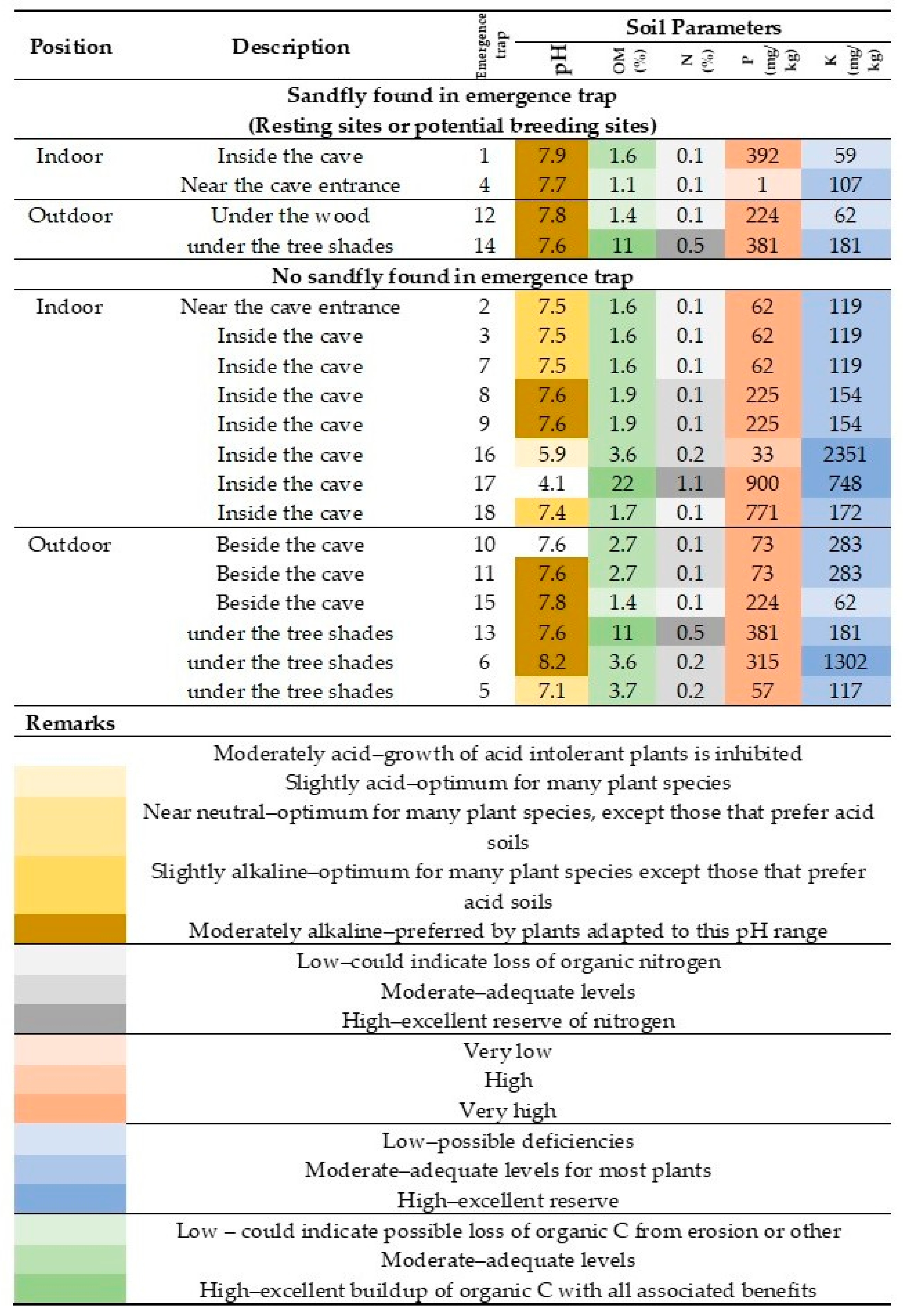
| Trap | Location of Resting Sites or Potential Breeding Sites | Month | Presence of Sandfly |
|---|---|---|---|
| Number | |||
| 01 | Inside the cave (aphotic area) | June | 1 female S. anodontis |
| 04 | Near the cave entrance (photic area) | May | 1 female Sergentomyia sp. |
| 12 | Outside the cave, under the wood piles | February | 1 male partially rotated genitalia |
| 14 | Outside the cave, under the tree shades | March | 1 male partially rotated genitalia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thammapalo, S.; Pawestri, A.R.; Kolaeh, K.; Boondej, P.; Benarlee, R.; Apiwathnasorn, C.; Kumlert, R. Distribution of Phlebotomine Sandflies in the Cave Area of Satun Province, Thailand. Trop. Med. Infect. Dis. 2020, 5, 174. https://doi.org/10.3390/tropicalmed5040174
Thammapalo S, Pawestri AR, Kolaeh K, Boondej P, Benarlee R, Apiwathnasorn C, Kumlert R. Distribution of Phlebotomine Sandflies in the Cave Area of Satun Province, Thailand. Tropical Medicine and Infectious Disease. 2020; 5(4):174. https://doi.org/10.3390/tropicalmed5040174
Chicago/Turabian StyleThammapalo, Suwich, Aulia Rahmi Pawestri, Kamal Kolaeh, Patcharida Boondej, Rittiporn Benarlee, Chamnarn Apiwathnasorn, and Rawadee Kumlert. 2020. "Distribution of Phlebotomine Sandflies in the Cave Area of Satun Province, Thailand" Tropical Medicine and Infectious Disease 5, no. 4: 174. https://doi.org/10.3390/tropicalmed5040174
APA StyleThammapalo, S., Pawestri, A. R., Kolaeh, K., Boondej, P., Benarlee, R., Apiwathnasorn, C., & Kumlert, R. (2020). Distribution of Phlebotomine Sandflies in the Cave Area of Satun Province, Thailand. Tropical Medicine and Infectious Disease, 5(4), 174. https://doi.org/10.3390/tropicalmed5040174