
Natural Infection and Vertical Transmission of Zika Virus in Sylvatic Mosquitoes Aedes albopictus and Haemagogus leucocelaenus from Rio de Janeiro, Brazil

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
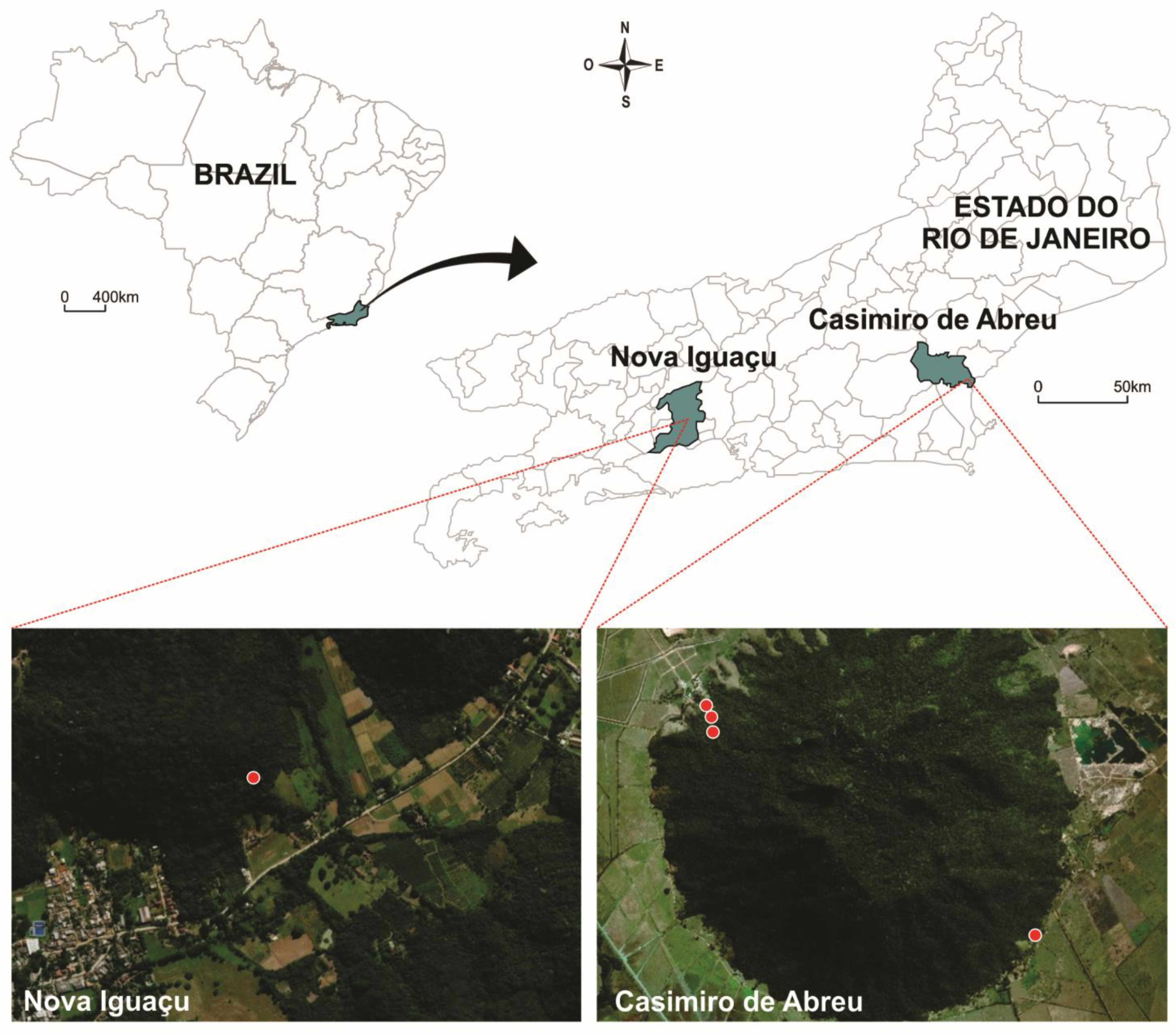
4.2. Study Areas
4.3. Mosquito Sampling
4.4. RNA Extraction
4.5. PCR for Flaviviruses
4.6. Nucleotide Sequencing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Franco, O. História da Febre Amarela no Brasil. Minist. Saúde Dep. Nac. Endem. Rurais. 1969, 1, 1–8. [Google Scholar]
- Soper, F.L.; Penna, H.; Cardoso, E.; Serafim, J., Jr.; Frobisher, M., Jr.; Pinheiro, J. Yellow fever without Aedes aegypti. Study of a rural epidemic in the Valle do Chanaan, Espirito Santo, Brazil, 1932. Am. J. Epidemiol. 1933, 18, 555–587. [Google Scholar] [CrossRef]
- Schuler-Faccini, L.; Ribeiro, E.M.; Feitosa, I.M.L.; Horovitz, D.D.G.; Cavalcanti, D.P.; Pessoa, A.; Doriqui, M.J.R.; Neri, J.I.; de Pina Neto, J.M.; Wanderley, H.Y.C.; et al. Possible Association Between Zika Virus Infection and Microcephaly—Brazil, 2015. Mmwr. Morb. Mortal. Wkly. Rep. 2016, 65, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.O.; Sampaio, S.A.; de Filippis, I.; Fabri, A.; Araujo, E.S.M.; de Sequeira, P.C.; de Mendonça, M.C.L.; de Oliveira, L.; et al. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: A case study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodušek, V.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Epelboin, Y.; Talaga, S.; Epelboin, L.; Dusfour, I. Zika virus: An updated review of competent or naturally infected mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef]
- Brasil, M.d.S. Boletim Epidemiológico 13: Monitoramento dos Casos de Arboviroses Urbanas Transmitidas pelo Aedes (Dengue, Chikungunya e Zika) até a Semana Epidemiológica 12 de 2019; 2019; Volume 50, pp. 1–18. Available online: https://portalarquivos2.saude.gov.br/images/pdf/2019/abril/30/2019-013-Monitoramento-dos-casos-de-arboviroses-urbanas-transmitidas-pelo-Aedes-publicacao.pdf (accessed on 8 February 2021).
- Pauvolid-Corrêa, A.; Campos, Z.; Juliano, R.; Velez, J.; Nogueira, R.M.R.; Komar, N. Serological Evidence of Widespread Circulation of West Nile Virus and Other Flaviviruses in Equines of the Pantanal, Brazil. PLoS Negl. Trop. Dis. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Catenacci, L.S.; Nunes-Neto, J.; Deem, S.L.; Palmer, J.L.; Travassos-da Rosa, E.S.; Tello, J.S. Diversity patterns of hematophagous insects in Atlantic forest fragments and human-modified areas of southern Bahia, Brazil. J. Vector Ecol. 2018, 43, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Moreira-Soto, A.; Carneiro, I.d.O.; Fischer, C.; Feldmann, M.; Kümmerer, B.M.; Silva, N.S.; Santos, U.G.; Souza, B.F.d.C.D.; Liborio, F.d.A.; Valença-Montenegro, M.M.; et al. Limited Evidence for Infection of Urban and Peri-urban Nonhuman Primates with Zika and Chikungunya Viruses in Brazil. mSphere 2018, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Favoretto, S.; Araújo, D.; Oliveira, D.; Duarte, N.; Mesquita, F.; Zanotto, P.; Durigon, E. First detection of Zika virus in neotropical primates in Brazil: A possible new reservoir. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- WHO. Yellow Fever—Brazil. Available online: http://www.who.int/csr/don/11-february-2019-yellow-fever-brazil/en/ (accessed on 8 February 2021).
- De Abreu, F.V.S.; Ribeiro, I.P.; Ferreira-De-Brito, A.; Dos Santos, A.A.C.; De Miranda, R.M.; Bonelly, I.D.S.; Neves, M.S.A.S.; Bersot, M.I.; Dos Santos, T.P.; Gomes, M.Q.; et al. Haemagogus leucocelaenus and Haemagogus janthinomys are the primary vectors in the major yellow fever outbreak in Brazil, 2016–2018. Emerg. Microbes Infect. 2019, 8, 218–231. [Google Scholar] [CrossRef] [Green Version]
- Marcondes, C.; Alencar, J. Revisão de mosquitos Haemagogus Williston (Diptera: Culicidae) do Brasil. Rev. Biomed. 2010, 21, 221–238. [Google Scholar]
- Cardoso, J.d.C.; Corseuil, E.; Barata, J.M.S. Culicinae (Diptera, Culicidae) ocorrentes no Estado do Rio Grande do Sul, Brasil. Rev. Bras. Entomol. 2005, 49, 275–287. [Google Scholar] [CrossRef]
- SES. Informe Epidemiológico 001/2018: Monitoramento das Emergências em Saúde Pública (Febre Amarela) 04 de Janeiro de 2018; pp. 1–6. Available online: http://www.riocomsaude.rj.gov.br/Publico/MostrarArquivo.aspx?C=%2Bp77vqe5mOs%3D (accessed on 8 February 2021).
- Johnson, B.K.; Chanas, A.C.; Shockley, P.; Squires, E.J.; Gardner, P.; Wallace, C.; Simpson, D.I.H.; Bowen, E.T.W.; Platt, G.S.; Way, H.; et al. Arbovirus isolations from, and serological studies on, wild and domestic vertebrates from Kano Plain, kenya. Trans. R. Soc. Trop. Med. Hyg. 1977, 71, 512–517. [Google Scholar] [CrossRef] [Green Version]
- Bueno, M.G.; Martinez, N.; Abdalla, L.; Duarte dos Santos, C.N.; Chame, M. Animals in the Zika Virus Life Cycle: What to Expect from Megadiverse Latin American Countries. PLoS Negl. Trop. Dis. 2016, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzian, A.C.B.; Zini, N.; Sacchetto, L.; Rocha, R.F.; Parra, M.C.P.; Del Sarto, J.L.; Dias, A.C.F.; Coutinho, F.; Rayra, J.; da Silva, R.A.; et al. Evidence of natural Zika virus infection in neotropical non-human primates in Brazil. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciota, A.T.; Bialosuknia, S.M.; Ehrbar, D.J.; Kramer, L.D. Vertical Transmission of Zika Virus by Aedes aegypti and Ae. Albopictus Mosquitoes. Emerg. Infect. Dis. 2017, 23, 880–882. [Google Scholar] [CrossRef]
- Costa-da-Silva, A.L.; Ioshino, R.S.; de Araújo, H.R.C.; Kojin, B.B.; de Andrade Zanotto, P.M.; Oliveira, D.B.L.; Melo, S.R.; Durigon, E.L.; Capurro, M.L. Laboratory strains of Aedes aegypti are competent to Brazilian Zika virus. PLoS ONE 2017, 12, e0171951. [Google Scholar] [CrossRef] [Green Version]
- Baleotti, F.G.; Moreli, M.L.; Figueiredo, L.T.M. Brazilian Flavivirus Phylogeny Based on NS5. Mem. Inst. Oswaldo Cruz 2003, 98, 379–382. [Google Scholar] [CrossRef] [Green Version]
- Pauvolid-Corrêa, A.; Kenney, J.L.; Couto-Lima, D.; Campos, Z.M.S.; Schatzmayr, H.G.; Nogueira, R.M.R.; Brault, A.C.; Komar, N. Ilheus Virus Isolation in the Pantanal, West-Central Brazil. PLoS Negl. Trop. Dis. 2013, 7, e2318. [Google Scholar] [CrossRef] [Green Version]
- Terzian, A.C.; Auguste, A.J.; Vedovello, D.; Ferreira, M.U.; da Silva-Nunes, M.; Sperança, M.A.; Suzuki, R.B.; Juncansen, C.; Araújo, J.P., Jr.; Weaver, S.C.; et al. Isolation and characterization of Mayaro virus from a human in Acre, Brazil. Am. J. Trop. Med. Hyg. 2015, 92, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-de-Brito, A.; Ribeiro, I.P.; de Miranda, R.M.; Fernandes, R.S.; Campos, S.S.; da Silva, K.A.B.; de Castro, M.G.; Bonaldo, M.C.; Brasil, P.; Lourenço-de-Oliveira, R. First detection of natural infection of Aedes aegypti with Zika virus in Brazil and throughout South America. Mem. Inst. Oswaldo Cruz 2016, 111, 655–658. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, C.J.; Miller, B.R.; Gubler, D.J. Vector competence of Aedes albopictus from Houston, Texas, for dengue serotypes 1 to 4, yellow fever and Ross River viruses. J. Am. Mosq. Control. Assoc. 1987, 3, 460–465. [Google Scholar] [PubMed]
- Miller, B.R.; Mitchell, C.J.; Ballinger, M.E. Replication, tissue tropisms and transmission of yellow fever virus in Aedes albopictus. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 252–255. [Google Scholar] [CrossRef]
- Chang, G.J.; Cropp, B.C.; Kinney, R.M.; Trent, D.W.; Gubler, D.J. Nucleotide sequence variation of the envelope protein gene identifies two distinct genotypes of yellow fever virus. J. Virol. 1995, 69, 5773–5780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, L.T.M. The Brazilian flaviviruses. Microbes Infect. 2000, 2, 1643–1649. [Google Scholar] [CrossRef]
- Johnson, B.W.; Chambers, T.V.; Crabtree, M.B.; Filippis, A.M.B.; Vilarinhos, P.T.R.; Resende, M.C.; Macoris, M.d.L.G.; Miller, B.R. Vector competence of Brazilian Aedes aegypti and Ae. albopictus for a Brazilian yellow fever virus isolate. Trans. R. Soc. Trop. Med. Hyg. 2002, 96, 611–613. [Google Scholar] [CrossRef]
- Lourenço de Oliveira, R.; Vazeille, M.; de Filippis, A.M.B.; Failloux, A.B. Large genetic differentiation and low variation in vector competence for dengue and yellow fever viruses of Aedes albopictus from Brazil, the United States, and the Cayman Islands. Am. J. Trop. Med. Hyg. 2003, 69, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Grard, G.; Caron, M.; Mombo, I.M.; Nkoghe, D.; Mboui Ondo, S.; Jiolle, D.; Fontenille, D.; Paupy, C.; Leroy, E.M. Zika Virus in Gabon (Central Africa)—2007: A New Threat from Aedes albopictus? PLoS Negl. Trop. Dis. 2014, 8, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Di Luca, M.; Severini, F.; Toma, L.; Boccolini, D.; Romi, R.; Remoli, M.E.; Sabbatucci, M.; Rizzo, C.; Venturi, G.; Rezza, G.; et al. Experimental studies of susceptibility of Italian Aedes albopictus to Zika virus. Euro Surveill. 2016, 21, 30223. [Google Scholar] [CrossRef]
- Du, S.; Liu, Y.; Liu, J.; Zhao, J.; Champagne, C.; Tong, L.; Zhang, R.; Zhang, F.; Qin, C.F.; Ma, P.; et al. Aedes mosquitoes acquire and transmit Zika virus by breeding in contaminated aquatic environments. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenço-de-Oliveira, R.; Failloux, A.B. Differential Susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika Virus. PLoS Negl. Trop. Dis. 2016, 10, 1–11. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Lai, Z.; Zhou, T.; Jia, Z.; Gu, J.; Wu, K.; Chen, X.G. Temperature Increase Enhances Aedes albopictus Competence to Transmit Dengue Virus. Front. Microbiol. 2017, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alencar, J.; Marcondes, C.B.; Serra-Freire, N.M.; Lorosa, E.S.; Pacheco, J.B.; Guimarães, A.É. Feeding patterns of Haemagogus capricornii and Haemagogus leucocelaenus (Díptera: Culicidae) in two Brazilian states (Rio de Janeiro and Goiás). J. Med. Entomol. 2008, 45, 873–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, B.M.C.; Coelho, F.C.; Armstrong, M.; Saraceni, V.; Lemos, C. Zika: An ongoing threat to women and infants. Cad. Saúde Pública 2018, 34, e00038218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, S.L.; Ponnusamy, L.; Unnasch, T.R.; Hassan, H.K.; Apperson, C.S. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) in relation to availability of human and domestic animals in suburban landscapes of central North Carolina. J. Med. Entomol. 2006, 43, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Faraji, A.; Egizi, A.; Fonseca, D.M.; Unlu, I.; Crepeau, T.; Healy, S.P.; Gaugler, R. Comparative Host Feeding Patterns of the Asian Tiger Mosquito, Aedes albopictus, in Urban and Suburban Northeastern USA and Implications for Disease Transmission. PLoS Negl. Trop. Dis. 2014, 8, e3037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, E.M.; Nkoghe, D.; Ollomo, B.; Nze-Nkogue, C.; Becquart, P.; Grard, G.; Pourrut, X.; Charrel, R.; Moureau, G.; Ndjoyi-Mbiguino, A.; et al. Concurrent chikungunya and dengue virus infections during simultaneous outbreaks, Gabon, 2007. Emerg. Infect. Dis. 2009, 15, 591–593. [Google Scholar] [CrossRef]
- Niebylski, M.L.; Craig, G.B., Jr. Dispersal and survival of Aedes albopictus at a scrap tire yard in Missouri. J. Am. Mosq Control. Assoc. 1994, 10, 339–343. [Google Scholar]
- Silva, N.I.O.; Sacchetto, L.; de Rezende, I.M.; Trindade, G.S.; LaBeaud, A.D.; de Thoisy, B.; Drumond, B.P. Recent sylvatic yellow fever virus transmission in Brazil: The news from an old disease. Virol. J. 2020, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.S.; Bersot, M.I.; Castro, M.G.; Telleria, E.L.; Ferreira-de-Brito, A.; Raphael, L.M.; Bonaldo, M.C.; Lorenço-de-Oliveira, R. Low vector competence in sylvatic mosquitoes limits Zika virus to initiate na enzootic cycle in South America. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Maciel-de-Freitas, R.; Neto, R.B.; Gonçalves, J.M.; Codeço, C.T.; Lourenço-De-Oliveira, R. Movement of dengue vectors between the human modified environment and an urban forest in Rio de Janeiro. J. Med. Entomol. 2006, 43, 1112–1120. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, C.F.; Da Silva, A.V.; Nascimento, V.A.D.; De Souza, V.C.; Monteiro, D.C.D.S.; Terrazas, W.C.M.; Dos Passos, R.A.; Nascimento, S.; Lima, J.B.P.; Naveca, F.G. Evidence of vertical transmission of Zika virus in field-collected eggs of in the Brazilian Amazon. PLoS Negl. Trop. Dis. 2018, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.D.C.; De Almeida, M.A.B.; Dos Santos, E.; Da Fonseca, D.F.; Sallum, M.A.M.; Noll, C.A.; Monteiro, H.A.D.O.; Cruz, A.C.R.; Carvalho, V.L.; Pinto, E.V.; et al. Yellow Fever Virus in Haemagogus leucocelaenus and Aedes serratus Mosquitoes, Southern Brazil, 2008. Emerg. Infect. Dis. 2010, 16, 1918–1924. [Google Scholar] [CrossRef]
- Instituto Nacional de Meteorologia—INMET. Available online: https://portal.inmet.gov.br/ (accessed on 8 February 2021).
- Silver, J.B. Mosquito Ecology. Field Sampling Methods, 3rd ed.; Springer: Dordrecht, The Netherlands, 2008; ISBN 1402066651. [Google Scholar]
- Alencar, J.; De Almeida, H.M.; Marcondes, C.B.; Guimarães, A.É. Effect of multiple immersions on eggs and development of immature forms of Haemagogus janthinomys from south-eastern Brazil (Diptera: Culicidae). Entomol. News 2008, 119, 239–244. [Google Scholar] [CrossRef]
- Alencar, J.; de Mello, C.F.; Gil-Santana, H.R.; Guimarães, A.É.; de Almeida, S.A.; Gleiser, R.M. Vertical oviposition activity of mosquitoes in the Atlantic Forest of Brazil with emphasis on the sylvan vector, Haemagogus leucocelaenus (Diptera: Culicidae). J. Vector Ecol. 2016, 41, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Consoli, R.A.G.B.; Oliveira, R.L.D. Principais Mosquitos de Importância Sanitária no Brasil—Fundação Oswaldo Cruz (Fiocruz): Ciência e Tecnologia em Saúde Para a População Brasileira; FIOCRUZ: Rio de Janeiro, Brazil, 1994; ISBN 85-85676-03-5. [Google Scholar]
- Reinert, J.F. List of abbreviations for currently valid generic-level taxa in family Culicidae (Diptera). J. Eur. Mosq. Control. Assoc. 2009, 27, 68–76. [Google Scholar]
- Patel, P.; Landt, O.; Kaiser, M.; Faye, O.; Koppe, T.; Lass, U.; Sall, A.A.; Niedrig, M. Development of one-step quantitative reverse transcription PCR for the rapid detection of flaviviruses. Virol. J. 2013, 10, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, T.D.; Vasconcellos, E.A.; Gomes, L.H.F.; Moreira, A.S.; Degrave, W.M.; Mendonça-Lima, L.; Alves-Ferreira, M. ChromaPipe: A pipeline for analysis, quality control and management for a DNA sequencing facility. Genet. Mol. Res. 2008, 7, 861–871. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Pool ID * | Mosquito Species | Sex | Total Mosquitoes | Month/Year Collected | Geographic Coordinates | Trap Identification | Pan-Flavivirus PCR Result | Sequence Match |
|---|---|---|---|---|---|---|---|---|
| 43 | Ae. albopictus | ♀ | 32 | Jan/19 | 22°33′01.3″ S 42°00′52.7″ W | TMPRNH -32 * | Positive | Zika virus |
| 45 | Ae. albopictus | ♂ | 2 | Oct/2018 | 22°31′40.1″ S 42°02′58.6″ W | TMF -2 * | Positive | Zika virus |
| 54 | Hg. leucocelaenus | ♀ | 6 | Jan/19 | 22°35′11.98″ S 43°24′34.12″ W | Tinguá * | Positive | Zika virus |
| 62 | Ae. Albopictus | ♀ | 9 | Oct/2018 | 22°31′43.9″ S 42°02′56.8″ W | TMF -3 * | Positive | Yellow fever virus |
| 64 | Hg. leucocelaenus | ♂ | 3 | Sep/2018 | 22°31′49.5″ S 42°02′56.3″ W | TMF -7 * | Positive | Yellow fever virus |
| 65 | Hg. leucocelaenus | ♂ | 5 | Oct/2018 | 22°31′49.5″ S 42°02′56.3″ W | TMF -7 * | Positive | Yellow fever virus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alencar, J.; Ferreira de Mello, C.; Brisola Marcondes, C.; Érico Guimarães, A.; Toma, H.K.; Queiroz Bastos, A.; Olsson Freitas Silva, S.; Lisboa Machado, S. Natural Infection and Vertical Transmission of Zika Virus in Sylvatic Mosquitoes Aedes albopictus and Haemagogus leucocelaenus from Rio de Janeiro, Brazil. Trop. Med. Infect. Dis. 2021, 6, 99. https://doi.org/10.3390/tropicalmed6020099
Alencar J, Ferreira de Mello C, Brisola Marcondes C, Érico Guimarães A, Toma HK, Queiroz Bastos A, Olsson Freitas Silva S, Lisboa Machado S. Natural Infection and Vertical Transmission of Zika Virus in Sylvatic Mosquitoes Aedes albopictus and Haemagogus leucocelaenus from Rio de Janeiro, Brazil. Tropical Medicine and Infectious Disease. 2021; 6(2):99. https://doi.org/10.3390/tropicalmed6020099
Chicago/Turabian StyleAlencar, Jeronimo, Cecilia Ferreira de Mello, Carlos Brisola Marcondes, Anthony Érico Guimarães, Helena Keiko Toma, Amanda Queiroz Bastos, Shayenne Olsson Freitas Silva, and Sergio Lisboa Machado. 2021. "Natural Infection and Vertical Transmission of Zika Virus in Sylvatic Mosquitoes Aedes albopictus and Haemagogus leucocelaenus from Rio de Janeiro, Brazil" Tropical Medicine and Infectious Disease 6, no. 2: 99. https://doi.org/10.3390/tropicalmed6020099
APA StyleAlencar, J., Ferreira de Mello, C., Brisola Marcondes, C., Érico Guimarães, A., Toma, H. K., Queiroz Bastos, A., Olsson Freitas Silva, S., & Lisboa Machado, S. (2021). Natural Infection and Vertical Transmission of Zika Virus in Sylvatic Mosquitoes Aedes albopictus and Haemagogus leucocelaenus from Rio de Janeiro, Brazil. Tropical Medicine and Infectious Disease, 6(2), 99. https://doi.org/10.3390/tropicalmed6020099