Using Fixed-Wing UAV for Detecting and Mapping the Distribution and Abundance of Penguins on the South Shetlands Islands, Antarctica

 ,
,  ,
, 
Abstract
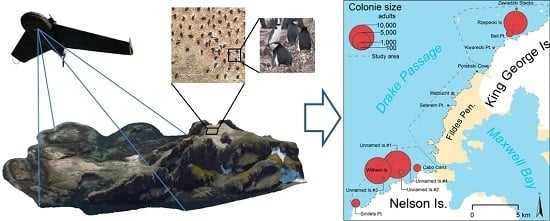
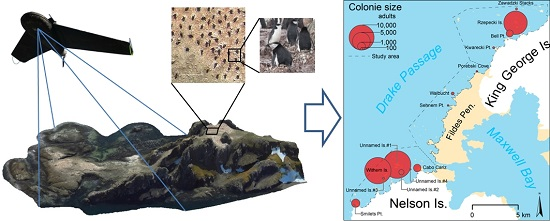
:
1. Introduction
2. Materials and Methods
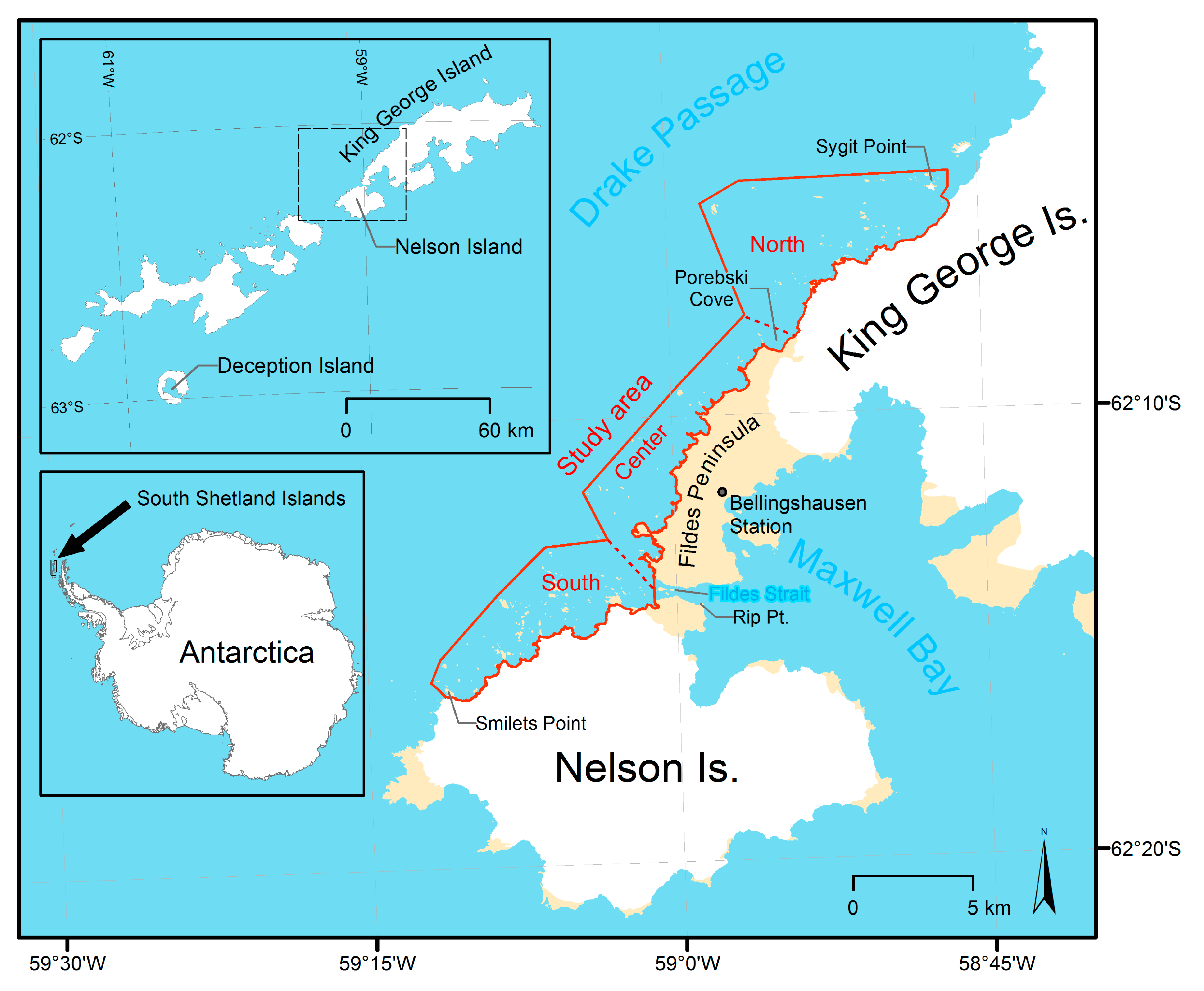
2.1. Study Area
- The southern subarea from Smilets Point to Fildes Strait at the glacierized coast of Nelson Island,
- the central subarea from Fildes Strait to Porebski Cove at the glacier-free coast of Fildes Peninsula, and
- the northern subarea from Porebski Cove to Sygit Point at the glacierized coast of King George Island.
2.2. Ground Survey at Fildes Peninsula
2.3. UAV and Camera
2.4. Geotagging and Orthorectification
2.5. Counts of Individual Adults in the Orthomosaics
2.6. Modeling Breeding Pair Numbers of UAV-Based Counts
2.6.1. Estimating the Number of Nests
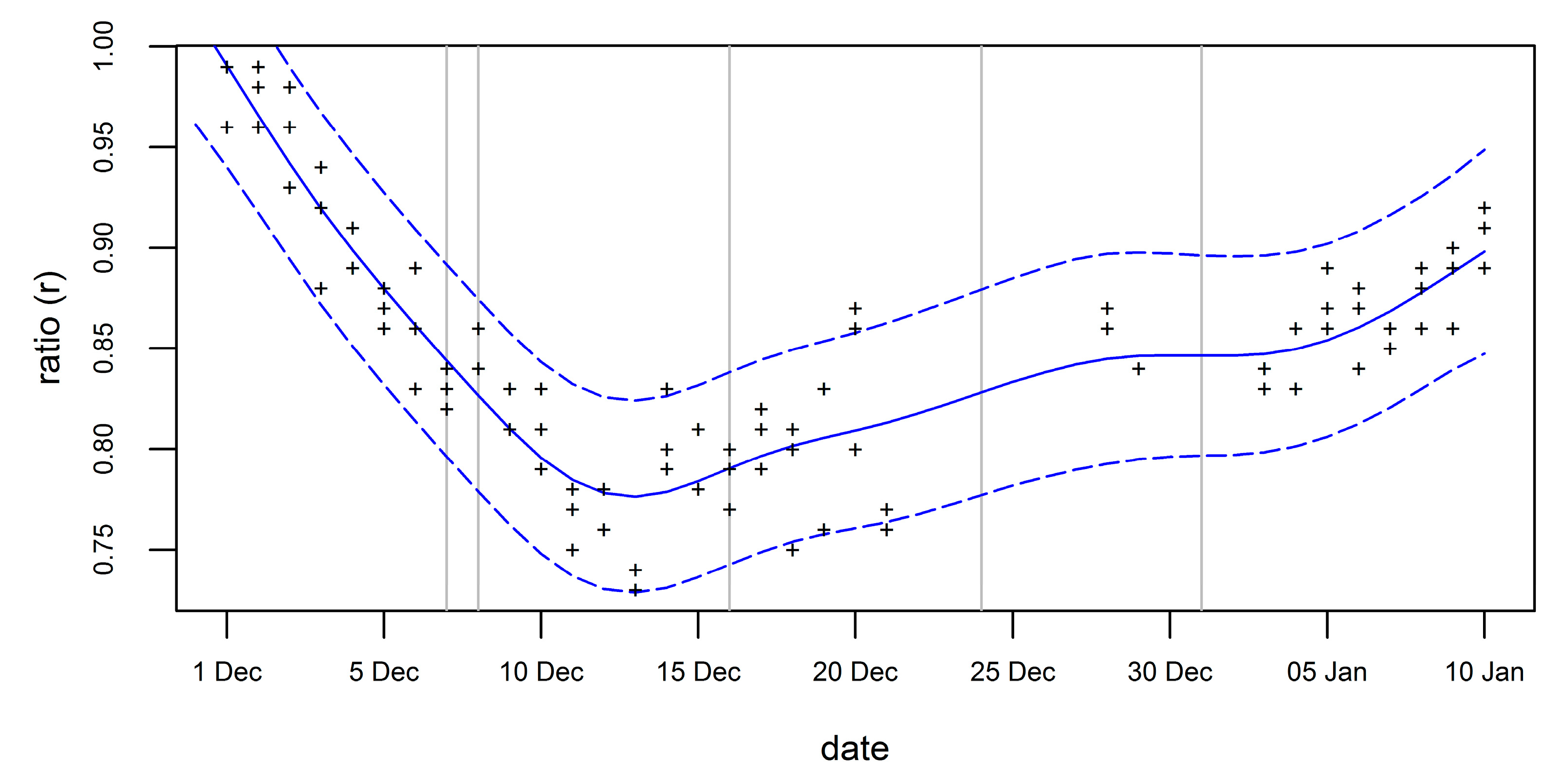
- We used time series of two time-lapse cameras at the Vapour Col colony (62.9912° S, 60.7201° W [50]) on Deception Island (South Shetland Islands; about 100 km southwest of Nelson Island) showing overviews of chinstrap nest groups. For the analysis, three images per day (01 Dec 2016 to 10 Jan 2017 at 11:00, 13:00 and 15:00 UTC-3h) were selected, since all UAV flights took place around these times (see Table 4). The number of adults was counted within a fixed area in every image, and the mean of the counts from both cameras,, was calculated, yielding up to three daily values for 32 days with good visibility for a total of 93 values of . We averaged all three daily values, since we found no systematic differences in the number of adults between these time points.
- We further determined the number of occupied nests, , one week after the peak of egg-laying (CCAMLR [51] Part I, Section 1: Method A3A) in the same area. The adults were counted using time-lapse imagery from 19 Nov 2016, which is one week after the local PoE (12 November 2016 [52]). For the standardization, we calculated the nest-to-individual ratio from the camera-derived and .
- A generalized additive model (GAM [53]) was used to model the (possibly nonlinear) temporal changes in the ratio . This allowed us to remove random variability and close gaps in the camera-derived time series that were caused by unusable or missing imagery (e.g., due to fog or camera malfunctioning). We used the software R (v.3.2.3) and its mgcv package [54]. The 95% prediction intervals of the predicted were used to represent and propagate uncertainties.
- The regional time shift in breeding phenology between camera and UAV sites was determined based on data from Hinke et al. [52], who reported that the PoE of chinstrap penguins in the 2016/17 season on KGI (colony Patelnia, 22 November 2016) took place three days later than on Deception Island (Vapour Col, 19 November 2016).
2.6.2. Accuracy Assessment of the Model
- Ground counts of 233 nests on Unnamed Islands #1 taken on 2 Jan 2018 around noon (about half an hour after the UAV flight)
- UAV-derived counts of adults covering the area of the ground counts
- The number of adults counted in the imagery of two time-lapse cameras of the Vapour Col colony (Deception Island) at the time of the UAV and ground surveys. There was no information on differences in breeding phenology ; a correction was therefore not applied.
- The number of occupied nests visible in time-lapse imagery of the Vapour Col colony taken one week after the local PoE (24 November 2017).
2.7. Change in Abundance and Distribution
2.7.1. Determination of Colony Locations of Previous Surveys
2.7.2. Modeling the Number of Nests of Previous Surveys
- The mean of the range of the number of adults given by Shuford and Spear [16]
- The maximum number of occupied nests n visible in time-lapse imagery of the Vapour Col colony (Deception Island) taken one week after the local PoE (12 November 2016; [52]).
- The number of adults counted in the imagery taken at noon (the exact time of the historic ground counts are unknown) of two time-lapse cameras of the Vapour Col colony at 20 Jan 2016 which is 69 days after the local PoE. We used 69 days because this is the period between the PoE and the counts in season 1984/85 by Shuford and Spear [16]. We estimated the date of the PoE in season 1984/85 using the modified model of Lynch et al. [46] (see Section 2.6).
2.7.3. Estimating the Change Including Uncertainty of Counts
3. Results
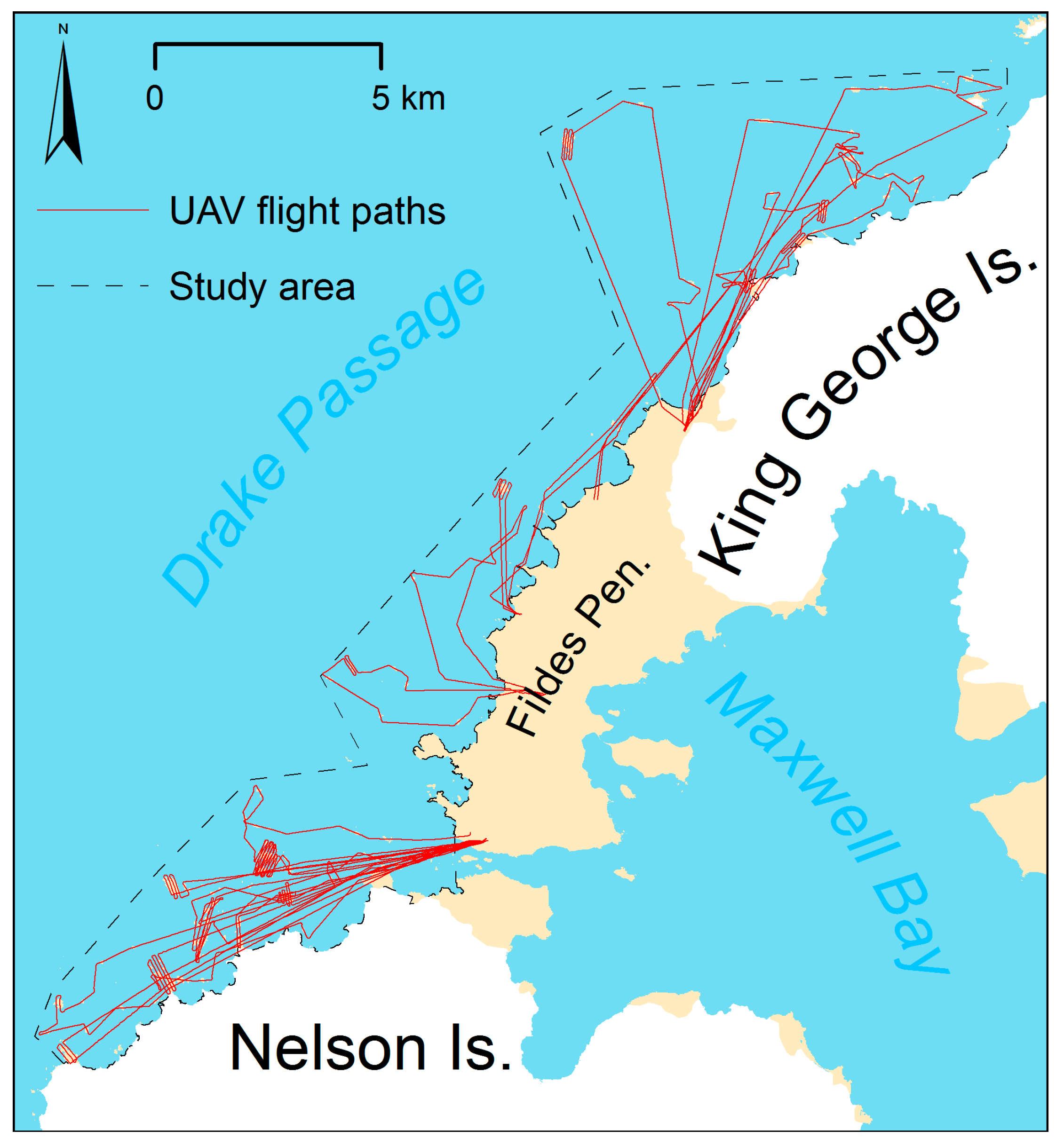
3.1. UAV Survey
3.2. Abundance and Distribution
3.3. Number of Occupied Nests
3.4. Long-Term Changes in Abundance and Distribution
3.4.1. Assignment of Colonies
3.4.2. Long-Term Change
4. Discussion
4.1. Comparison with Earlier Survey Methods
4.2. Comparison with Other UAV Types
4.3. Abundance and Distribution
4.4. Changes in Distribution and Abundance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clarke, A.; Murphy, E.J.; Meredith, M.P.; King, J.C.; Peck, L.S.; Barnes, D.K.; Smith, R.C. Climate change and the marine ecosystem of the western Antarctic Peninsula. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 149–166. [Google Scholar] [CrossRef]
- Croxall, J.P.; Trathan, P.N.; Murphy, E.J. Environmental Change and Antarctic Seabird Populations. Science 2002, 297, 1510. [Google Scholar] [CrossRef] [PubMed]
- Chabot, D.; Bird, D.M. Wildlife research and management methods in the 21st century: Where do unmanned aircraft fit in? J. Unmanned Veh. Syst. 2015, 3, 137–155. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Baylis, S.M.; Mott, R.; Herrod, A.; Clarke, R.H. Precision wildlife monitoring using unmanned aerial vehicles. Sci. Rep. 2016, 6, 22574. [Google Scholar] [CrossRef] [PubMed]
- Rush, G.P.; Clarke, L.E.; Stone, M.; Wood, M.J. Can drones count gulls? Minimal disturbance and semiautomated image processing with an unmanned aerial vehicle for colony-nesting seabirds. Ecol. Evol. 2018, 8, 12322–12334. [Google Scholar] [CrossRef] [PubMed]
- Zmarz, A.; Rodzewicz, M.; Dąbski, M.; Karsznia, I.; Korczak-Abshire, M.; Chwedorzewska, K.J. Application of UAV BVLOS remote sensing data for multi-faceted analysis of Antarctic ecosystem. Remote Sens. Environ. 2018, 217, 375–388. [Google Scholar] [CrossRef]
- Forcada, J.; Trathan, P.N.; Reid, K.; Murphy, E.J.; Croxall, J.P. Contrasting population changes in sympatric penguin species in association with climate warming. Glob. Chang. Biol. 2006, 12, 411–423. [Google Scholar] [CrossRef]
- Trivelpiece, W.Z.; Trivelpiece, S.G.; Geupel, G.R.; Kjelmyr, J.; Volkman, N.J. Adélie and Chinstrap Penguins: Their Potential as Monitors of the Southern Ocean Marine Ecosystem. In Antarctic Ecosystems; Kerry, K.R., Hempel, G., Eds.; Springer: Berlin/Heidelberg, Germany, 1990; pp. 191–202. [Google Scholar]
- Lynch, H.J.; White, R.; Naveen, R.; Black, A.; Meixler, M.S.; Fagan, W.F. In stark contrast to widespread declines along the Scotia Arc, a survey of the South Sandwich Islands finds a robust seabird community. Polar Biol. 2016, 39, 1615–1625. [Google Scholar] [CrossRef]
- Barbosa, A.; Benzal, J.; De León, A.; Moreno, J. Population decline of chinstrap penguins (Pygoscelis antarctica) on Deception Island, South Shetlands, Antarctica. Polar Biol. 2012, 35, 1453–1457. [Google Scholar] [CrossRef]
- Naveen, R.; Lynch, H.J.; Forrest, S.; Mueller, T.; Polito, M. First direct, site-wide penguin survey at Deception Island, Antarctica, suggests significant declines in breeding chinstrap penguins. Polar Biol. 2012, 35, 1879–1888. [Google Scholar] [CrossRef]
- Sander, M.; Balbão, T.C.; Polito, M.J.; Costa, E.S.; Carneiro, A.P.B. Recent decrease in chinstrap penguin (Pygoscelis antarctica) populations at two of Admiralty Bay’s islets on King George Island, South Shetland Islands, Antarctica. Polar Biol. 2007, 30, 659–661. [Google Scholar] [CrossRef]
- Bannasch, R.; Odening, K. Zoologische Untersuchungen im Gebiet der sowjetischen Antarktisstation “Bellingshausen”. Geodätische Und Geophys. Veröffentlichungen 1981, Reihe I 8, 3–20. [Google Scholar]
- Peter, H.-U.; Kaiser, M.; Gebauer, A. Untersuchungen an Vögeln und Robben auf King George Island (South Shetland Islands, Antarktis). Geodätische Und Geophys. Veröffentlichungen 1988, Reihe I 14, 1–127. [Google Scholar]
- Jablonski, B. Distribution and numbers of penguins in the region of King George Island (South Shetland Islands) in the breeding season 1980/1981. Pol. Polar Res 1984, 5, 17–30. [Google Scholar]
- Shuford, W.D.; Spear, L.B. Surveys of breeding Chinstrap Penguins in the South Shetland Islands, Antarctica. Br. Antarct. Surv. Bull. 1988, 81, 19–30. [Google Scholar]
- Braun, C.; Esefeld, J.; Peter, H.-U. Monitoring the Consequences of Local Climate Change on the Natural Resources of the Ice-Free Regions of Maxwell Bay (King George Island, Antarctic); Texte 26/2017; German Environment Agency: Dessau-Roßlau, Germany, 2017; pp. 1–177. [Google Scholar]
- Peter, H.-U.; Braun, C.; Janowski, S.; Nordt, A.; Stelter, M. The Current Environmental Situation and Proposals for the Management of the Fildes Peninsula Region; Texte 02/2013; Federal Environment Agency (Germany): Dessau-Roßlau, Germany, 2013; pp. 1–127. [Google Scholar]
- Peter, H.-U.; Bueßer, C.; Mustafa, O.; Pfeifer, S. Risk Assessment for the Fildes Peninsula and Ardley Island, and Development of Management Plans for Their Designation as Specially Protected or Specially Managed Areas; Texte 20/08; Federal Environment Agency: Dessau-Roßlau, Germany, 2008; pp. 1–344. [Google Scholar]
- Mustafa, O.; Esefeld, J.; Grämer, H.; Maercker, J.; Rümmler, M.-C.; Pfeifer, C. Monitoring Penguin Colonies in the Antarctic Using Remote Sensing Data; Texte 30/2017; German Environment Agency: Dessau-Roßlau, Germany, 2017; pp. 1–162. [Google Scholar]
- Korczak−Abshire, M.; Węgrzyn, M.; Angiel, P.J.; Lisowska, M. Pygoscelid penguins breeding distribution and population trends at Lions Rump rookery, King George Island. Pol. Polar Res. 2013, 34, 87–99. [Google Scholar] [CrossRef]
- Kokubun, N.; Lee, W.Y.; Kim, J.-H.; Takahashi, A. Chinstrap penguin foraging area associated with a seamount in Bransfield Strait, Antarctica. Polar Sci. 2015, 9, 393–400. [Google Scholar] [CrossRef]
- Southwell, C.; Emmerson, L.; McKinlay, J.; Newbery, K.; Takahashi, A.; Kato, A.; Barbraud, C.; DeLord, K.; Weimerskirch, H. Spatially Extensive Standardized Surveys Reveal Widespread, Multi-Decadal Increase in East Antarctic Adélie Penguin Populations. PLoS ONE 2015, 10, e0139877. [Google Scholar] [CrossRef]
- Lynch, H.J.; LaRue, M.A. Erratum: First global census of the Adélie Penguin. Auk 2015, 132, 562. [Google Scholar] [CrossRef]
- Lynch, H.J.; Schwaller, M.R. Mapping the Abundance and Distribution of Adélie Penguins Using Landsat-7: First Steps towards an Integrated Multi-Sensor Pipeline for Tracking Populations at the Continental Scale. PLoS ONE 2014, 9, e113301. [Google Scholar] [CrossRef]
- Schwaller, M.R.; Lynch, H.J.; Tarroux, A.; Prehn, B. A continent-wide search for Antarctic petrel breeding sites with satellite remote sensing. Remote Sens. Environ. 2018, 210, 444–451. [Google Scholar] [CrossRef]
- Schwaller, M.R.; Southwell, C.J.; Emmerson, L.M. Continental-scale mapping of Adélie penguin colonies from Landsat imagery. Remote Sens. Environ. 2013, 139, 353–364. [Google Scholar] [CrossRef]
- Borowicz, A.; McDowall, P.; Youngflesh, C.; Sayre-McCord, T.; Clucas, G.; Herman, R.; Forrest, S.; Rider, M.; Schwaller, M.; Hart, T.; et al. Multi-modal survey of Adélie penguin mega-colonies reveals the Danger Islands as a seabird hotspot. Sci. Rep. 2018, 8, 3926. [Google Scholar] [CrossRef]
- Goebel, M.E.; Perryman, W.L.; Hinke, J.T.; Krause, D.J.; Hann, N.A.; Gardner, S.; LeRoi, D.J. A small unmanned aerial system for estimating abundance and size of Antarctic predators. Polar Biol. 2015, 38, 619–630. [Google Scholar] [CrossRef]
- Perryman, W.; Goebel, M.E.; Ash, N.; LeRoi, D.; Gardner, S. Small unmanned aerial systems for estimating abundance of krill-dependent predators: A feasibility study with preliminary results. AMLR 2014, 2011, 64–72. [Google Scholar]
- Ratcliffe, N.; Guihen, D.; Robst, J.; Crofts, S.; Stanworth, A.; Enderlein, P. A protocol for the aerial survey of penguin colonies using UAVs. J. Unmanned Veh. Syst. 2015, 3, 95–101. [Google Scholar] [CrossRef]
- Goetzendorf-Grabowski, T.; Rodzewicz, M. Design of UAV for photogrammetric mission in Antarctic area. Proc. Inst. Mech. Eng. Part G J. Aerosp. Eng. 2016. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Mott, R.; Baylis, S.M.; Pham, T.T.; Wotherspoon, S.; Kilpatrick, A.D.; Raja Segaran, R.; Reid, I.; Terauds, A.; Koh, L.P. Drones count wildlife more accurately and precisely than humans. Methods Ecol. Evol. 2018, 9, 1160–1167. [Google Scholar] [CrossRef]
- Rümmler, M.-C.; Mustafa, O.; Maercker, J.; Peter, H.-U.; Esefeld, J. Sensitivity of Adélie and Gentoo penguins to various flight activities of a micro UAV. Polar Biol. 2018, 41, 2481–2493. [Google Scholar] [CrossRef]
- Rümmler, M.-C.; Mustafa, O.; Maercker, J.; Peter, H.-U.; Esefeld, J. Measuring the influence of unmanned aerial vehicles on Adélie penguins. Polar Biol. 2016, 39, 1329–1334. [Google Scholar] [CrossRef]
- Verfuss, U.K.; Aniceto, A.S.; Harris, D.V.; Gillespie, D.; Fielding, S.; Jiménez, G.; Johnston, P.; Sinclair, R.R.; Sivertsen, A.; Solbø, S.A.; et al. A review of unmanned vehicles for the detection and monitoring of marine fauna. Mar. Pollut. Bull. 2019, 140, 17–29. [Google Scholar] [CrossRef]
- Barsch, D.; Blümel, W.-D. Untersuchungen zum Periglazial auf der König-Georg-Insel, Südshetlandinseln/Antarktika: Deutsche physiogeographische Forschungen in der Antarktis; Bericht über die Kampagne 1983/84. Ber. Zur Polarforsch. (Rep. Polar Res.) 1985, 24, 76. [Google Scholar]
- Trujillo, A.P.; Thurman, H.V. Essentials of Oceanography, 11th ed.; Pearson: Boston, MA, USA, 2014; ISBN 978-0-321-81405-0. [Google Scholar]
- Johnston, D.W. Unoccupied Aircraft Systems in Marine Science and Conservation. Annu. Rev. Mar. Sci. 2019, 11, 439–463. [Google Scholar] [CrossRef] [PubMed]
- Dellaert, F.; Seitz, S.M.; Thorpe, C.E.; Thrun, S. Structure from motion without correspondence. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Hilton Head Island, SC, USA, 15 June 2000; Volume 2, pp. 557–564. [Google Scholar]
- Fretwell, P.T.; Phillips, R.A.; Brooke, M.d.L.; Fleming, A.H.; McArthur, A. Using the unique spectral signature of guano to identify unknown seabird colonies. Remote Sens. Environ. 2015, 156, 448–456. [Google Scholar] [CrossRef]
- Rees, W.G.; Brown, J.A.; Fretwell, P.T.; Trathan, P.N. What colour is penguin guano? Antarct. Sci. 2017, 29, 417–425. [Google Scholar] [CrossRef]
- Schwaller, M.R.; Olson, C.E.; Ma, Z.; Zhu, Z.; Dahmer, P. A remote sensing analysis of Adélie penguin rookeries. Remote Sens. Environ. 1989, 28, 199–206. [Google Scholar] [CrossRef]
- Southwell, C.; McKinlay, J.; Low, M.; Wilson, D.; Newbery, K.; Lieser, J.L.; Emmerson, L. New methods and technologies for regional-scale abundance estimation of land-breeding marine animals: Application to Adélie penguin populations in East Antarctica. Polar Biol. 2013, 36, 843–856. [Google Scholar] [CrossRef]
- Ainley, D.G. The Adélie Penguin: Bellwether of Climate Change; Ebrary Inc.: New York, NY, USA, 2002; ISBN 0-231-12306-X. [Google Scholar]
- Lynch, H.J.; Fagan, W.F.; Naveen, R.; Trivelpiece, S.G.; Trivelpiece, W.Z. Timing of clutch initiation in Pygoscelis penguins on the Antarctic Peninsula: Towards an improved understanding of off-peak census correction factors. Ccamlr Sci. 2009, 16, 149–165. [Google Scholar]
- McKinlay, J.; Southwell, C.; Trebilco, R. Integrating Count Effort by Seasonally Correcting Animal Population Estimates (ICESCAPE): A method for estimating abundance and its uncertainty from count data using Adélie penguins as a case study. Ccamlr Sci. 2010, 17, 213–227. [Google Scholar]
- Southwell, C.; Emmerson, L. Remotely-operating camera network expands Antarctic seabird observations of key breeding parameters for ecosystem monitoring and management. J. Nat. Conserv. 2015, 23, 1–8. [Google Scholar] [CrossRef]
- Müller-Schwarze, D. The Behavior of Penguins: Adapted to Ice and Tropics; SUNY Series in Animal Behavior; State University of New York Press: Albany, NY, USA, 1984; ISBN 978-0-87395-866-0. [Google Scholar]
- Barbosa, A.; Moreno, J.; Potti, J.; Merino, S. Breeding group size, nest position and breeding success in the chinstrap penguin. Polar Biol. 1997, 18, 410–414. [Google Scholar] [CrossRef]
- CCAMLR CCAMLR Ecosystem Monitoring Program: Standard Methods; CCAMLR: Hobart, Australia, 2014.
- Hinke, J.T.; Barbosa, A.; Emmerson, L.M.; Hart, T.; Juáres, M.A.; Korczak-Abshire, M.; Milinevsky, G.; Santos, M.; Trathan, P.N.; Watters, G.M.; et al. Estimating nest-level phenology and reproductive success of colonial seabirds using time-lapse cameras. Methods Ecol. Evol. 2018, 9, 1853–1863. [Google Scholar] [CrossRef]
- Hastie, T.J.; Tibshirani, R.J. Generalized Additive Models, Volume 43 of Monographs on Statistics and Applied Probability; Chapman & Hall: London, UK, 1990. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC: Boca Raton, FL, USA, 2006. [Google Scholar]
- Black, C.E. A comprehensive review of the phenology of Pygoscelis penguins. Polar Biol. 2016, 39, 405–432. [Google Scholar] [CrossRef]
- Erfurt, J.; Grimm, H. Expeditionsbericht der 2. DDR-Antarktisexpedition, Überwinterungsteilnehmer an der 34. Sowjetischen Antarktisexpedition, Station “Bellingshausen” 1988-1990; Unpublished Expedition report; Akademie der Wissenschaften: Potsdam, Germany, 1990. [Google Scholar]
- Mönke, R.; Bick, A. Fachlicher Bericht über die Teilnahme der DDR-Biologengruppe an der 31. Sowjetischen Antarktisexpedition (SAE), Station Bellingshausen, King George Island (Südshetland Inseln/Antarktis); Unpublished Expedition Report; Akademie der Wissenschaften: Potsdam, Germany, 1988. [Google Scholar]
- Woehler, E.J. The Distribution and Abundance of Antarctic and Subantarctic Penguins. Polar Rec. 1993, 30, 66. [Google Scholar]
- Mesas-Carrascosa, F.-J.; Notario García, M.D.; Meroño de Larriva, J.E.; García-Ferrer, A. An Analysis of the Influence of Flight Parameters in the Generation of Unmanned Aerial Vehicle (UAV) Orthomosaicks to Survey Archaeological Areas. Sensor 2016, 16, 1838. [Google Scholar] [CrossRef]
- Croxall, J.P.; Kirkwood, E.D. The Distribution of Penguins on the Antarctic Peninsula and Islands of the Scotia Sea; British Antarctic Survey: Cambridge, UK, 1979. [Google Scholar]
- Zippel, D. Teilbericht über die Wissenschaftlichen Ergebnisse Während der 30. SAE im Untersuchungsgebiet King George Island; Unpublished Expedition Report; Akademie der Wissenschaften: Potsdam, Germany, 1987. [Google Scholar]
- Southwell, C.; Emmerson, L.; Newbery, K.; McKinlay, J.; Kerry, K.; Woehler, E.; Ensor, P. Re-Constructing Historical Adélie Penguin Abundance Estimates by Retrospectively Accounting for Detection Bias. PLoS ONE 2015, 10, e0123540. [Google Scholar] [CrossRef]
- Korczak-Abshire, M.; Zmarz, A.; Rodzewicz, M.; Kycko, M.; Karsznia, I.; Chwedorzewska, K.J. Study of fauna population changes on Penguin Island and Turret Point Oasis (King George Island, Antarctica) using an unmanned aerial vehicle. Polar Biol. 2019, 42, 217–224. [Google Scholar] [CrossRef]
- Grenzdörffer, G.J. UAS-based automatic bird count of a common gull colony. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2013, 1, W2. [Google Scholar] [CrossRef]
- Harris, C.M. Aircraft operations near concentrations of birds in Antarctica: The development of practical guidelines. Biol. Conserv. 2005, 125, 309–322. [Google Scholar] [CrossRef]
- Sander, M.; Carneiro, A.P.; Balbao, T.C.; Bays, S.R.; Costa, E.S.; Mascarello, N.E.; Oliva, T.D.; dos Santos, C.R. Status and Trends of Antarctic Seabirds at Admiralty Bay, King George Island. Polarforschung 2006, 75, 145–150. [Google Scholar]
- Petry, M.V.; Valls, F.C.L.; Petersen, E.d.S.; Krüger, L.; Piuco, R.d.C.; dos Santos, C.R. Breeding sites and population of seabirds on Admiralty Bay, King George Island, Antarctica. Polar Biol. 2016, 39, 1343–1349. [Google Scholar] [CrossRef]
- Silva, M.P.; Favero, M.; Casaux, R.; Baroni, A. The status of breeding birds at Harmony Point, Nelson Island, Antarctica in summer 1995/96. Mar. Ornithol. 1998, 26, 75–78. [Google Scholar]
- Gil-Delgado, J.A.; González-Solís, J.; Barbosa, A. Populations of breeding birds in Byers Peninsula, Livingston Island, South Shetland Islands. Antarct. Sci. 2013, 25, 303–306. [Google Scholar] [CrossRef]
- Lynch, H.J.; Naveen, R.; Fagan, W.F. Censuses of penguin, blue-eyed shag Phalacrocorax atriceps and southern giant petrel Macronectes giganteus populations on the Antarctic Peninsula, 2001–2007. Mar. Ornithol. 2008, 36, 83–97. [Google Scholar]
- Ainley, D.G.; Blight, L.K. Ecological repercussions of historical fish extraction from the Southern Ocean. Fish Fish. 2009, 10, 13–38. [Google Scholar] [CrossRef]
- ATCM Antarctic Specially Protected Area No 171 (Narębski Point, Barton Peninsula, King George Island): Revised Management Plan. In Proceedings of the Annex II, Brasilia, Brazil, 28 April–7 May 2014.
- Secretariat, S.C.A.R. Composite Gazetteer of Antarctica, Scientific Committee on Antarctic Research. GCMD Metadata. Available online: http://gcmd.nasa.gov/records/SCAR_Gazetteer.html (accessed on 25 June 2018).
- Cervellati, R.; Ramorino, C.; Sievers, J.; Thomson, J.; Clarke, D. A composite gazetteer of Antarctica. Polar Rec. 2000, 36, 278. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | |
|---|---|
| Size (in flight mode) | 1.0 m wingspan, 0.64 m length |
| Take-off weight of the modified UAV | 1 kg |
| Motor/Propellers | 160 W brushless motor with 8 × 4.5″ folding propellers |
| Batteries | 2 × 3S 2200 mAh lithium polymer |
| Autopilot | Pixhawk 1 |
| GNSS | uBlox Neo M8N module (GPS & GLONASS were used) |
| Telemetry | SiK Telemetry Radio, max. 100 mW output power, 433 MHz band with an omnidirectional antenna |
| Electronic speed control (ESC) | max. 25 A |
| Additional sensors | Pixhawk digital differential airspeed sensor (4525DO) MAPIR Survey-2 RGB digital camera |
| Category | Description |
|---|---|
| Breeding group | Occupied nests with a distance < 2 m |
| Subcolony | Breeding groups with a distance < 50 m |
| Colony | Subcolonies with a distance < 500 m |
| Breeding site | Topographically distinct (ice-free) areas with recent and/or historic colonies |
| Base of Counts and Accuracy | Inferred Uncertainty |
|---|---|
| A1, N1, C1 (±0 to 5%) | 0.05 |
| A2, N2, C2 (±5 to 10%) | 0.1 |
| A3, N3, C3 (±10 to 25%) | 0.25 |
| A4, N4, C4 (±25 to 50%) | 0.5 |
| A5, N5, C5 (nearest order of magnitude) | - |
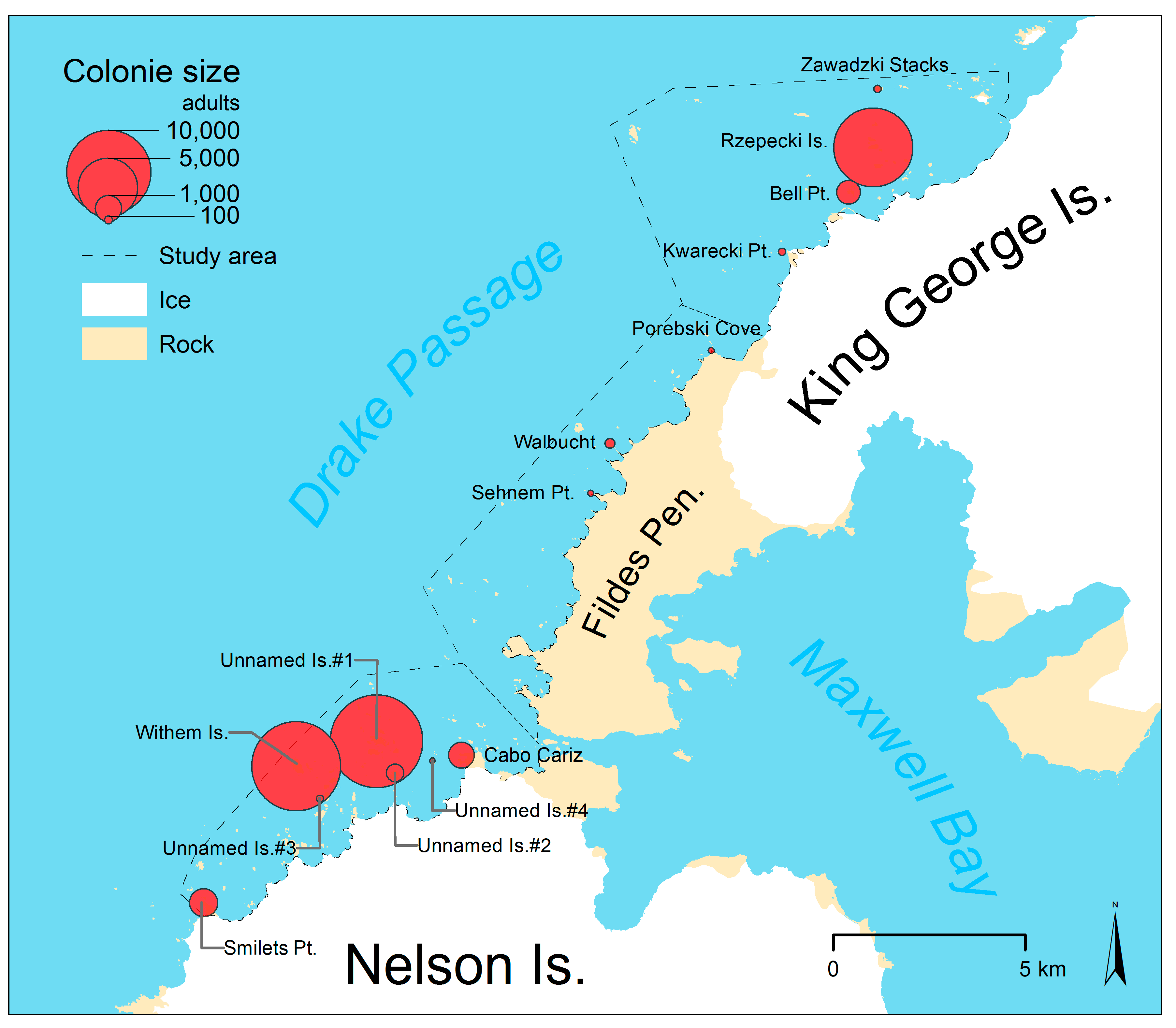
| Sub-Area | Site | Number of Adults | Modeled Number of Nests ± ME | Survey Date/ Time (UTC−3h) | Lat. | Lon. | ||
|---|---|---|---|---|---|---|---|---|
| Sub-Colony | Colony | Sub-Colony | Colony | |||||
| North | Zawadzki Stacks | − | 83 | − | 68 ± 4 | 28 Dec 2016/14:34 | −62.0815 | −58.8388 |
| Rzepecki Islands | 2486 | 8703 | 2046 ± 126 | 7162 ± 440 | 28 Dec 2016/13:28 | −62.0995 | −58.8394 | |
| 3327 | 2738 ± 168 | −62.0951 | −58.8417 | |||||
| 2890 | 2378 ± 146 | −62.0934 | −58.8416 | |||||
| Bell Point | 807 | 869 | 664 ± 41 | 715 ± 44 | 28 Dec 2016/11:49 | −62.1054 | −58.8546 | |
| 5 | 4 ± 0 | −62.1071 | −58.8592 | |||||
| 57 | 47 ± 3 | −62.106 | −58.8594 | |||||
| Kwarecki Point | 85 | 136 | 70 ± 4 | 112 ± 6 | 28 Dec 2016/11:59 | −62.1188 | −58.8887 | |
| 46 | 38 ± 2 | −62.119 | −58.891 | |||||
| 5 | 4 ± 0 | −62.1187 | −58.8861 | |||||
| Center | Porebski Cove | − | 61 | − | 50 ± 3 | 28 Dec 2016/12:19 | −62.1415 | −58.9254 |
| Walbucht | − | 155 | − | 131 ± 7 | 12 Dec 2016/12:54 | −62.1625 | −58.9773 | |
| Sehnem Point | − | − | − | 48 1 | 15 Dec 2016 | −62.174 | −58.9877 | |
| South | Unnamed Islands #1 | 1460 | 11,959 | 1145 ± 70 | 9376 ± 569 | 20 Dec 2016/11:07 | −62.2330 | −59.0889 |
| 26 | 20 ± 1 | −62.2307 | −59.0900 | |||||
| 5625 | 4410 ± 268 | −62.2304 | −59.0985 | |||||
| 145 | 114 ± 7 | −62.2311 | −59.0928 | |||||
| 136 | 107 ± 6 | −62.2308 | −59.1024 | |||||
| 1370 | 1074 ± 65 | −62.2280 | −59.0984 | |||||
| 3197 | 2506 ± 152 | −62.2329 | −59.1010 | |||||
| Unnamed Island #4 | − | 41 2 | − | 35 ± 2 | 11 Dec 2016/14:11 | −62.2354 | −59.0709 | |
| Cabo Cariz | − | 935 | − | 791 ± 48 | 04 Jan 2017/15:04 | −62.2343 | −59.0563 | |
| Unnamed Island #3 | − | 63 2 | − | 53 ± 3 | 04 Jan 2017/12:42 | −62.2433 | −59.1279 | |
| Unnamed Island #2 | 408 | 444 | 352 ± 19 | 383 ± 21 | 11 Dec 2016/14:09 | −62.2375 | −59.0902 | |
| 7 | 6 ± 0 | −62.2385 | −59.0914 | |||||
| 29 | 25 ± 1 | −62.2391 | −59.0881 | |||||
| Withem Island | 10,921 | 11,007 | 9409 ± 520 | 9483 ± 524 | 11 Dec 2016/12:24 | −62.2356 | −59.1392 | |
| 86 | 74 ± 4 | −62.2355 | −59.1335 | |||||
| Smilets Point | 570 | 1148 | 491 ± 27 | 989 ± 55 | 11 Dec 2016/15:06 | −62.2667 | −59.1878 | |
| 578 | 498 ± 28 | −62.2685 | −59.1863 | |||||
| Reference | Description in Reference (Translated to English) | Assigned Position |
|---|---|---|
| Jablonski [15] | “Islands between Bell Island and Kwarecki Pt.” - Map #41 | Kwarecki Point |
| Shuford and Spear [16] | “15,000-20,000 adults” at No. 36 | Unnamed Islands #1 |
| Peter et al. [14] | “Figure 8” | Unnamed Island #2 |
| Peter et al. [14] | “off Withem Island”/“Figure 8” | Withem Island |
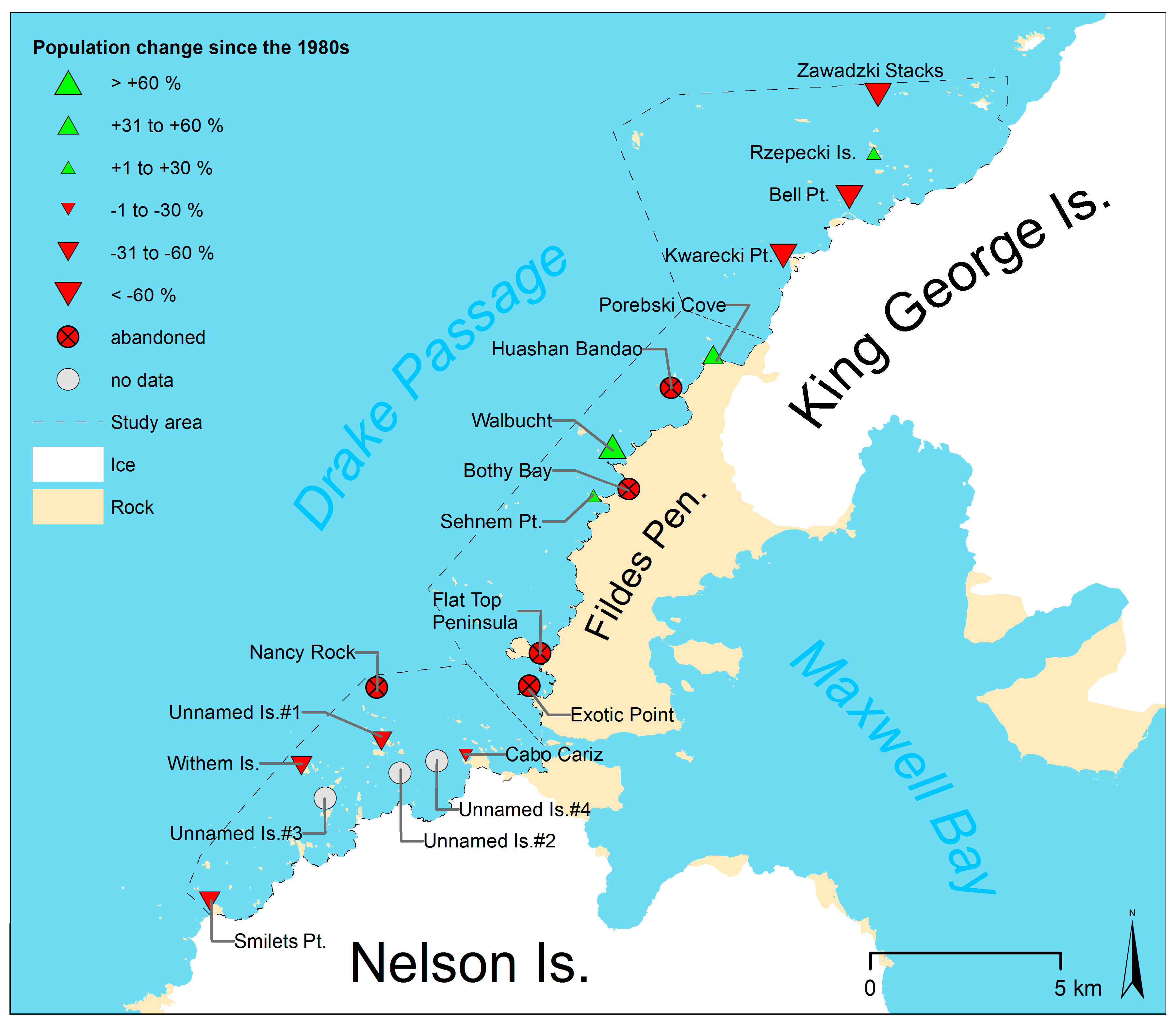
| Sub-Area | Site | Colony Size [Nests] | Base of Counts and Accuracy | Change [%] (Uncertainty Interval) | Season | Reference |
|---|---|---|---|---|---|---|
| North | Zawadzki Stacks | 450 | N2 or C2 | −85 (−87, −82) | 1980/81 | Jablonski [15] |
| 68 | A1 | 2016/17 | This study | |||
| Rzepecki Islands | 6430 | N2 or C2 | 12 (−5, 31) | 1980/81 | Jablonski [15] | |
| 7162 | A1 | 2016/17 | This study | |||
| Bell Point | 3274 | N1−2 or C2 | −78 (−81, −74) | 1980/81 | Jablonski [15] | |
| 715 | A1 | 2016/17 | This study | |||
| Kwarecki Point | 800 | N2 or C2 | −86 (−88, −84) | 1980/81 | Jablonski [15] | |
| 112 | A1 | 2016/17 | This study | |||
| Center | Porebski Cove | 32 | N1 | 56 (40, 74) | 1980/81 | Jablonski [15] |
| 50 | A1 | 2016/17 | This study | |||
| Huashan Bandao | 20−30 | A3−4 | abandoned | 1980/81 | Jablonski [15] | |
| 0 | A1 | 2016/17 | This study | |||
| Walbucht | 39 | N1 | 235 (203, 272) | 1984/85 | Peter et al. [14] | |
| 131 | A1 | 2016/17 | This study | |||
| Bothy Bay | 3 | N1 | abandoned | 1984/85 | Peter et al. [14] | |
| 0 | A1 | 2016/17 | This study | |||
| Sehnem Point | 36 | N1 | 33 (21, 47) | 1984/85 | Peter et al. [14] | |
| 48 | N1 | 2016/17 | This study | |||
| Flat Top Peninsula | 18 | N1 | abandoned | 1980/81 | Jablonski [15] | |
| 0 | A1 | 2016/17 | This study | |||
| Exotic Point | 25 | N1 | abandoned | 1984/85 | Peter et al. [14] | |
| 0 | A1 | 2016/17 | This study | |||
| South | Nancy Rock | “covered with penguins” | abandoned | 1986/87 | Shuford and Spear [16] | |
| 0 | A1 | 2016/17 | This study | |||
| Unnamed Islands #1 | 21,444 | A3 | −56 (−64, −45) | 1986/87 | Shuford and Spear [16] | |
| 9376 | A1 | 2016/17 | This study | |||
| Cabo Cariz | 1000 | N4 | −21 (−50, 68) | 1984/85 | Peter et al. [14] | |
| 791 | A1 | 2016/17 | This study | |||
| Unnamed Island #2 | 100−999 | N5 | not defined | 1984/85 | Peter et al. [14] | |
| 383 | A1 | 2016/17 | This study | |||
| Withem Islands | 20,000 | N4 | −53 (−70, 0) | 1984/85 | Peter et al. [14] | |
| 9483 | A1 | 2016/17 | This study | |||
| Smilets Point | 2142 | A3 | −54 (−62, −43) | 1986/87 | Shuford and Spear [16] | |
| 989 | A1 | 2016/17 | This study | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfeifer, C.; Barbosa, A.; Mustafa, O.; Peter, H.-U.; Rümmler, M.-C.; Brenning, A. Using Fixed-Wing UAV for Detecting and Mapping the Distribution and Abundance of Penguins on the South Shetlands Islands, Antarctica. Drones 2019, 3, 39. https://doi.org/10.3390/drones3020039
Pfeifer C, Barbosa A, Mustafa O, Peter H-U, Rümmler M-C, Brenning A. Using Fixed-Wing UAV for Detecting and Mapping the Distribution and Abundance of Penguins on the South Shetlands Islands, Antarctica. Drones. 2019; 3(2):39. https://doi.org/10.3390/drones3020039
Chicago/Turabian StylePfeifer, Christian, Andres Barbosa, Osama Mustafa, Hans-Ulrich Peter, Marie-Charlott Rümmler, and Alexander Brenning. 2019. "Using Fixed-Wing UAV for Detecting and Mapping the Distribution and Abundance of Penguins on the South Shetlands Islands, Antarctica" Drones 3, no. 2: 39. https://doi.org/10.3390/drones3020039
APA StylePfeifer, C., Barbosa, A., Mustafa, O., Peter, H. -U., Rümmler, M. -C., & Brenning, A. (2019). Using Fixed-Wing UAV for Detecting and Mapping the Distribution and Abundance of Penguins on the South Shetlands Islands, Antarctica. Drones, 3(2), 39. https://doi.org/10.3390/drones3020039