
A Narrative Review of Auditory Categorisation and Its Potential Role in Tinnitus Perception


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
3. Results and Discussion
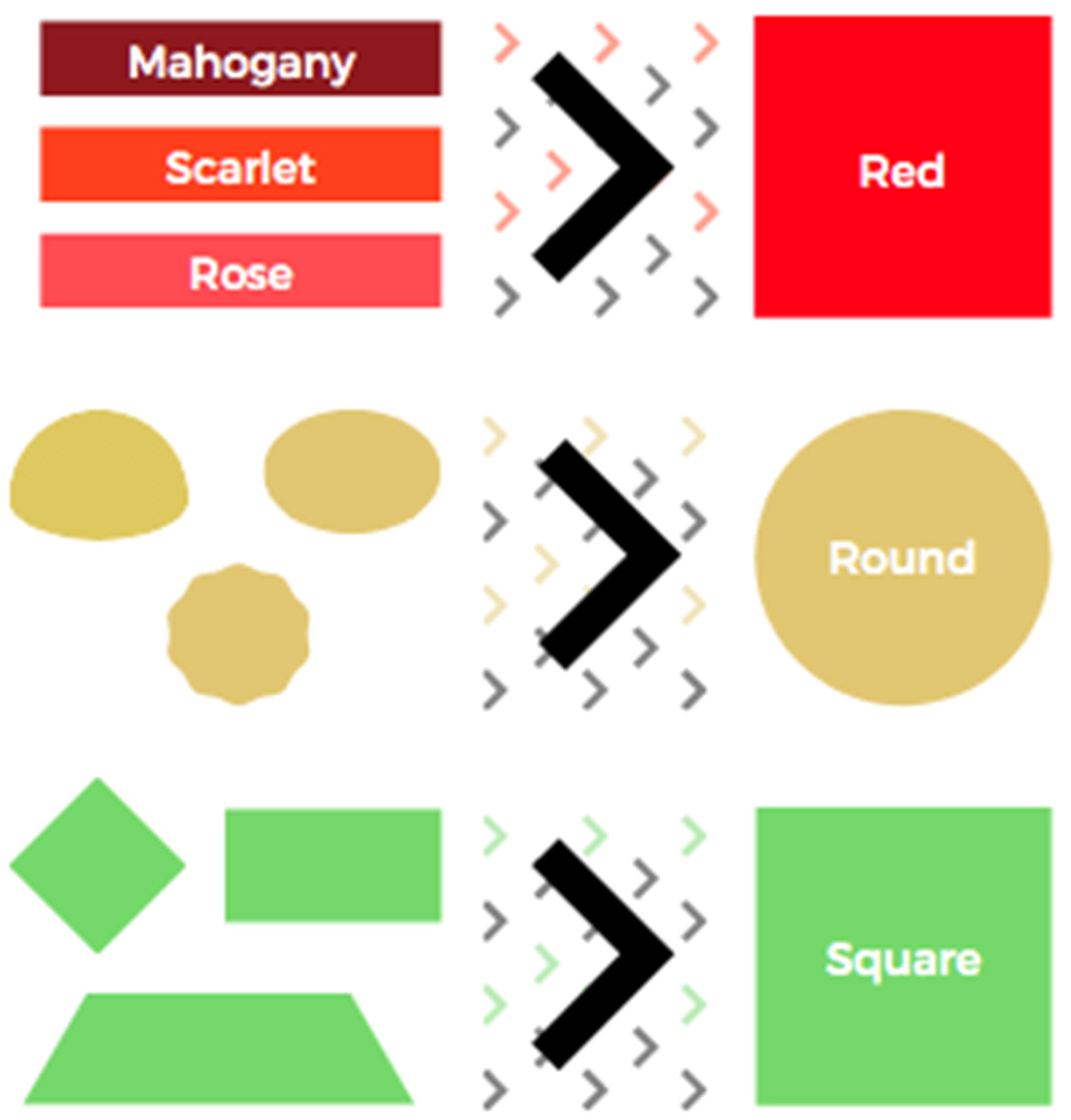
3.1. Categorisation: The What, When, and Where
3.1.1. Categorisation and Its Origin
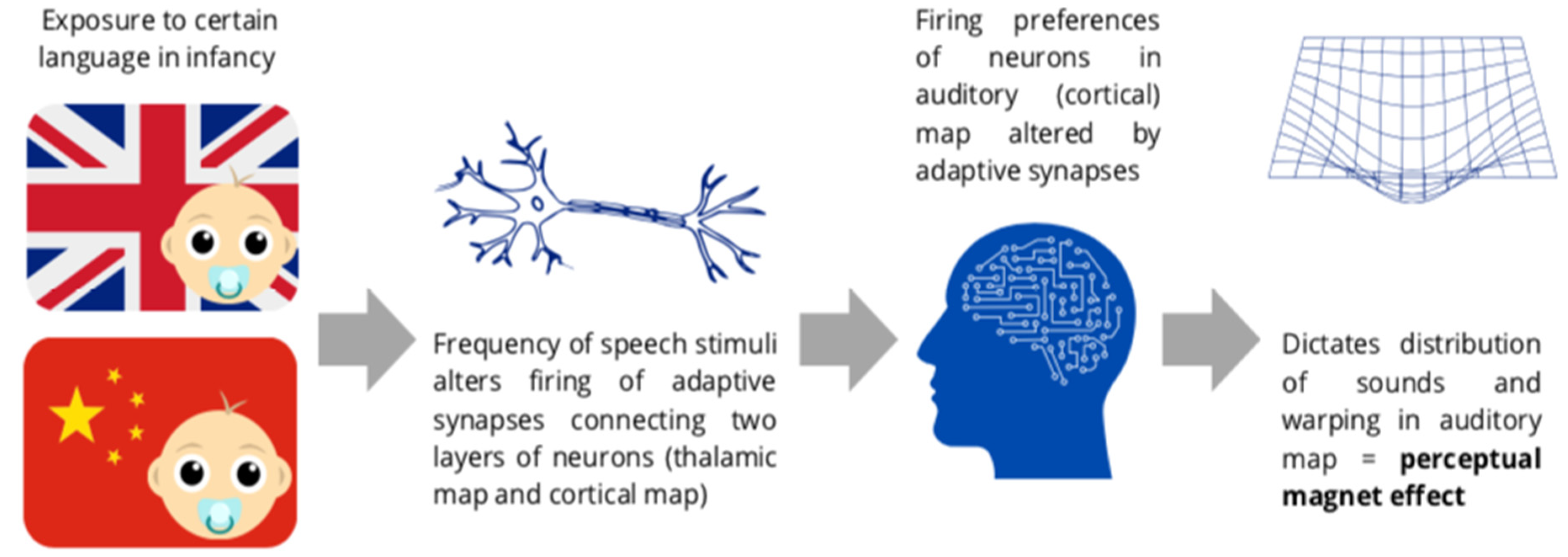
3.1.2. Warping of the Auditory Perceptual Space
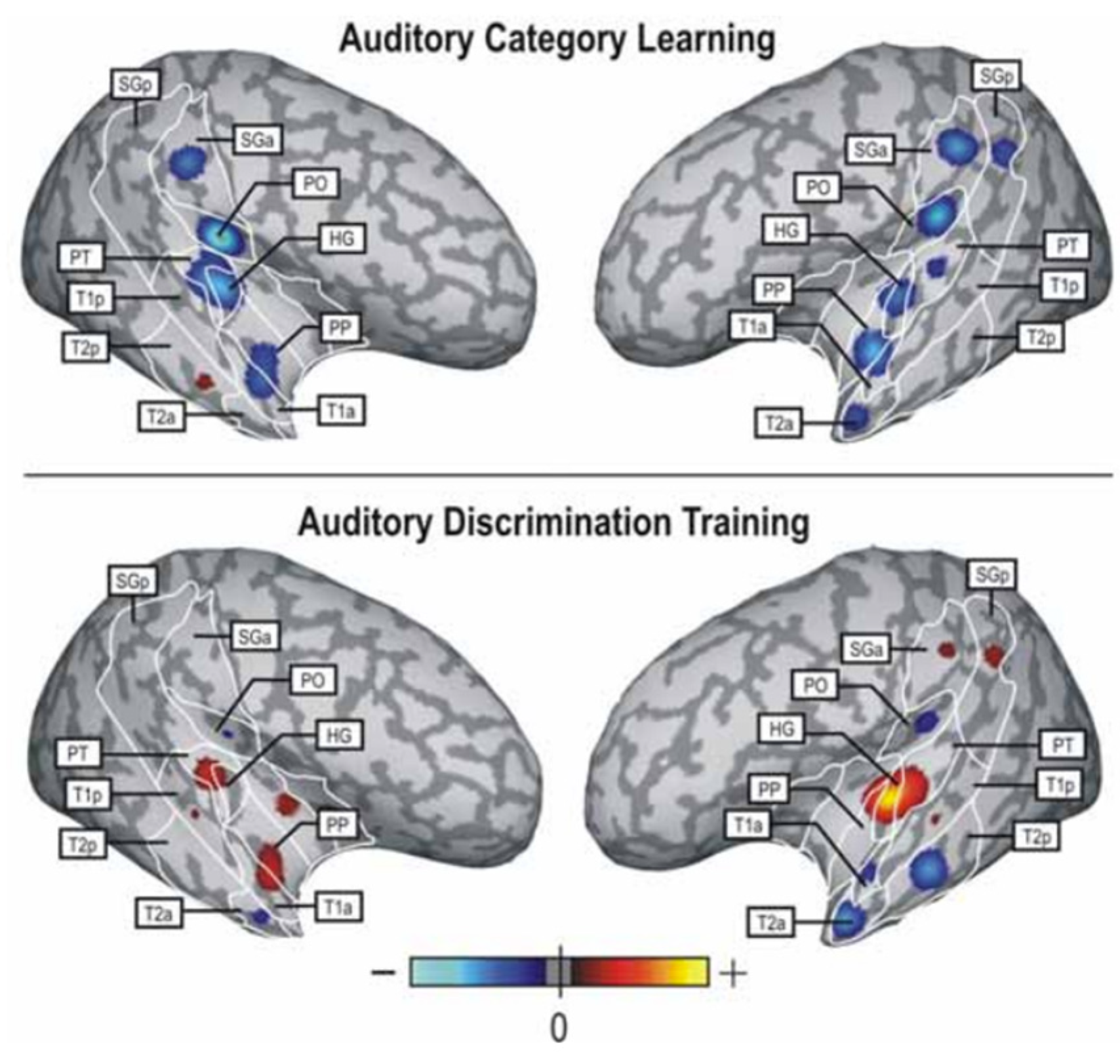
3.1.3. Neural Changes Resulting from Categorisation Training
3.1.4. Passive Categorisation: A Consequence of Evolution?
3.2. Categorisation as an Innate Feature
3.2.1. Cross Cultural Studies
3.2.2. Infancy Studies
3.2.3. Animal Studies
3.2.4. Passive Categorisation in Vision
3.3. Categorisation as an Learned Feature
3.4. Controversy Surrounding Categorisation
3.5. Categorisation for Tinnitus
4. Conclusions
Funding
Conflicts of Interest
References
- Guenther, F.H.; Nieto-Castañón, A.; Ghosh, S.S.; Tourville, J.A. Representation of Sound Categories in Auditory Cortical Maps. J. Speech Lang. Hear. Res. 2004, 47, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, P.K. Human adults and human infants show a “perceptual magnet effect” for the prototypes of speech categories, monkeys do not. Percept. Psychophys. 1991, 50, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Guenther, F.H.; Bohland, J. Learning sound categories: A neural model and supporting experiments. Acoust. Sci. Technol. 2002, 23, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Guenther, F.H.; Gjaja, M.N. The perceptual magnet effect as an emergent property of neural map formation. J. Acoust. Soc. Am. 1996, 100, 1111–1121. [Google Scholar] [CrossRef] [Green Version]
- Harnad, S. Psychophysical and cognitive aspects of categorical perception: A critical overview. In Categorical Perception: The Groundwork of Cognition; Cambridge University Press: New York, NY, USA, 1987; pp. 1–52. [Google Scholar]
- Beale, J.M.; Keil, F.C. Categorical Perception as an Acquired Phenomenon: What are the Implications? In Neural Computation and Psychology; Springer: London, UK, 1995. [Google Scholar]
- Burns, E.M.; Ward, W.D. Categorical perception—Phenomenon or epiphenomenon: Evidence from experiments in the perception of melodic musical intervals. J. Acoust. Soc. Am. 1978, 63, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, R.L. Influences of categorization on perceptual discrimination. J. Exp. Psychol. Gen. 1994, 123, 178. [Google Scholar] [CrossRef] [PubMed]
- Guenther, F.H.; Husain, F.T.; Cohen, M.A.; Shinn-Cunningham, B.G. Effects of categorization and discrimination training on auditory perceptual space. J. Acoust. Soc. Am. 1999, 106, 2900–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, H. The motor theory of speech perception: A critical review. Psychol. Rev. 1965, 72, 275–309. [Google Scholar] [CrossRef]
- Liberman, A.M. Some results of research on speech perception. J. Acoust. Soc. Am. 1957, 29, 117–123. [Google Scholar] [CrossRef]
- Eggermont, J.J.; Tass, P. Maladaptive Neural Synchrony in Tinnitus: Origin and Restoration. Front. Neurol. 2015, 6, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norena, A.J.; Eggermont, J.J. Changes in spontaneous neural activity immediately after an acoustic trauma: Implications for neural correlates of tinnitus. Hear. Res. 2003, 183, 137–153. [Google Scholar] [CrossRef]
- Roberts, L.E. Neural plasticity and its initiating conditions in tinnitus. HNO 2017, 66, 172–178. [Google Scholar] [CrossRef]
- Green, B.N.; Johnson, C.D.; Adams, A. Writing narrative literature reviews for peer-reviewed journals: Secrets of the trade. J. Chiropr. Med. 2006, 5, 101–117. [Google Scholar] [CrossRef] [Green Version]
- Liberman, A.M.; Harris, K.S.; Hoffman, H.S.; Griffith, B.C. The discrimination of speech sounds within and across phoneme boundaries. J. Exp. Psychol. 1957, 54, 358–368. [Google Scholar] [CrossRef] [Green Version]
- Lively, S.E.; Pisoni, D.B. On prototypes and phonetic categories: A critical assessment of the perceptual magnet effect in speech perception. J. Exp. Psychol. Hum. Percept. Perform. 1997, 23, 1665. [Google Scholar] [CrossRef]
- Samuel, A.G. Phonetic prototypes. Percept. Psychophys. 1982, 31, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iverson, P.; Kuhl, P.K. Mapping the perceptual magnet effect for speech using signal detection theory and multidimensional scaling. J. Acoust. Soc. Am. 1995, 97, 553–562. [Google Scholar] [CrossRef]
- Aaltonen, O.; Eerola, O.; Hellström, Å.; Uusipaikka, E.; Lang, A.H. Perceptual magnet effect in the light of behavioral and psychophysiological data. J. Acoust. Soc. Am. 1997, 101, 1090–1105. [Google Scholar] [CrossRef]
- Bornstein, M.H. Perceptual categories in vision and audition. In Categorical Perception: The Groundwork of Cognition; Cambridge University Press: New York, NY, USA, 1987; pp. 287–300. [Google Scholar]
- Harnad, S. Categorical perception. In Encyclopedia of Cognitive Science; Nadel, L., Ed.; Nature Publishing Group: Berlin, Germany, 2003; pp. 64–67. [Google Scholar]
- Whorf, B.L. Language, Thought, and Reality: Selected Writings; Technology Press of Massachusetts Institute of Technology: Cambridge, MA, USA, 1956. [Google Scholar]
- Abramson, A.S.; Lisker, L. Discriminability along the voicing continuum: Cross-language tests. In Proceedings of the Sixth International Congress of Phonetic Sciences, Prague, Czech Republic, 7–13 September 1967; Volume 196, pp. 569–573. [Google Scholar]
- Berlin, B.; Kay, P. Basic Color Terms: Their Universality and Evolution; University of California Press: Berkeley, CA, USA, 1969. [Google Scholar]
- Boynton, R.M.; Gordon, J. Bezold–Brücke hue shift measured by color-naming technique. JOSA 1965, 55, 78–86. [Google Scholar] [CrossRef]
- Eimas, P.D.; Siqueland, E.R.; Jusczyk, P.; Vigorito, J. Speech Perception in Infants. Science 1971, 171, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K. Speech perception in early infancy: Perceptual constancy for spectrally dissimilar vowel categories. J. Acoust. Soc. Am. 1979, 66, 1668–1679. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K.; Miller, J.D. Speech perception by the chinchilla: Identification functions for synthetic VOT stimuli. J. Acoust. Soc. Am. 1978, 63, 905–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyawaki, K.; Strange, W.; Verbrugge, R.R.; Liberman, A.M.; Jenkins, J.J.; Fujimura, O. An effect of linguistic experience: The discrimination of (r) and (l) by native speakers of Japanese and English. Percept. Psychophys. 1975, 18, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Sandell, J.H.; Gross, C.G.; Bornstein, M.H. Color categories in macaques. J. Comp. Physiol. Psychol. 1979, 93, 626. [Google Scholar] [CrossRef] [PubMed]
- von Frisch, K. Bees: Their Vision, Chemical Senses, and Language; Cornell University Press: Ithaca, NY, USA, 1964. [Google Scholar]
- Werker, J.F.; Gilbert, J.H.; Humphrey, K.; Tees, R.C. Developmental aspects of cross-language speech perception. Child Dev. 1981, 52, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Werker, J.F.; Tees, R.C. Cross-language speech perception: Evidence for perceptual reorganization during the first year of life. Infant Behav. Dev. 1984, 7, 49–63. [Google Scholar] [CrossRef]
- Wright, A.A. The influence of ultraviolet radiation on the pigeon’s color discrimination. J. Exp. Anal. Behav. 1972, 17, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.A.; Cumming, W.W. Color-naming functions for the pigeon. J. Exp. Anal. Behav. 1971, 15, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Gandour, J.T.; Francis, A.L. Effects of language experience and stimulus complexity on the categorical perception of pitch direction. J. Acoust. Soc. Am. 2006, 120, 1063–1074. [Google Scholar] [CrossRef] [Green Version]
- Stevens, K.; Libermann, A.; Studdert-Kennedy, M.; Öhman, S. Crosslanguage Study of Vowel Perception. Lang. Speech 1969, 12, 1–23. [Google Scholar] [CrossRef]
- Lasky, R.E.; Syrdal-Lasky, A.; Klein, R.E. VOT Discrimination by four to six and a half month old infants from Spanish environments. J. Exp. Child Psychol. 1975, 20, 215–225. [Google Scholar] [CrossRef]
- Streeter, L.A. Language perception of 2-month-old infants shows effects of both innate mechanisms and experience. Nature 1976, 259, 39–41. [Google Scholar] [CrossRef] [PubMed]
- Trehub, S.E.; Rabinovitch, M.S. Auditory-linguistic sensitivity in early infancy. Dev. Psychol. 1972, 6, 74–77. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, P.K. Perceptual constancy for speech-sound categories in early infancy. Child Phonol. 1980, 2, 41–66. [Google Scholar]
- Kuhl, P.K. Perception of auditory equivalence classes for speech in early infancy. Infant Behav. Dev. 1983, 6, 263–285. [Google Scholar] [CrossRef]
- Jusczyk, P.W.; Rosner, B.S.; Cutting, J.E.; Foard, C.F.; Smith, L.B. Categorical perception of nonspeech sounds by 2-month-old infants. Percept. Psychophys. 1977, 21, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Sinnott, J.M.; Beecher, M.D.; Moody, D.B.; Stebbins, W.C. Speech sound discrimination by monkeys and humans. J. Acoust. Soc. Am. 1976, 60, 687–695. [Google Scholar] [CrossRef]
- Kuhl, P.K.; Padden, D.M. Enhanced discriminability at the phonetic boundaries for the voicing feature in macaques. Percept. Psychophys. 1982, 32, 542–550. [Google Scholar] [CrossRef] [Green Version]
- May, B.; Moody, D.B.; Stebbins, W.C. Categorical perception of conspecific communication sounds by Japanese macaques, Macaca fuscata. J. Acoust. Soc. Am. 1989, 85, 837–847. [Google Scholar] [CrossRef]
- Waters, R.; Wilson, W.A. Speech perception by rhesus monkeys: The voicing distinction in synthesized labial and velar stop consonants. Percept. Psychophys. 1976, 19, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Recanzone, G.H.; Schreiner, C.; Merzenich, M.M. Plasticity in the frequency representation of primary auditory cortex following discrimination training in adult owl monkeys. J. Neurosci. 1993, 13, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Ehret, G.; Haack, B. Categorical perception of mouse pup ultrasound by lactating females. Naturwissenschaften 1981, 68, 208–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kluender, K.R.; Diehl, R.L.; Killeen, P.R. Japanese Quail Can Learn Phonetic Categories. Science 1987, 237, 1195–1197. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.A.; Marler, P. Categorical Perception of a Natural Stimulus Continuum: Birdsong. Science 1989, 244, 976–978. [Google Scholar] [CrossRef] [PubMed]
- Hulse, S.H.; Cynx, J. Relative pitch perception is constrained by absolute pitch in songbirds (Mimus, Molothrus, and Sturnus). J. Comp. Psychol. 1985, 99, 176. [Google Scholar] [CrossRef]
- Dooling, R.J.; Brown, S.D. Speech perception by budgerigars (Melopsittacus undulatus): Spoken vowels. Percept. Psychophys. 1990, 47, 568–574. [Google Scholar] [CrossRef]
- Eriksson, J.L.; Villa, A.E. Learning of auditory equivalence classes for vowels by rats. Behav. Process. 2006, 73, 348–359. [Google Scholar] [CrossRef]
- Ohl, F.W.; Scheich, H.; Freeman, W.J. Change in pattern of ongoing cortical activity with auditory category learning. Nature 2001, 412, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, W.; Wagner, T.; Ohl, F.; Scheich, H. Categorical discrimination of direction in frequency-modulated tones by Mongolian gerbils. Behav. Brain Res. 1998, 91, 29–39. [Google Scholar] [CrossRef]
- Bornstein, M.H.; Kessen, W.; Weiskopf, S. Color vision and hue categorization in young human infants. J. Exp. Psychol. Hum. Percept. Perform. 1976, 2, 115. [Google Scholar] [CrossRef] [PubMed]
- Gaißert, N.; Waterkamp, S.; Fleming, R.W.; Bülthoff, I. Haptic Categorical Perception of Shape. PLoS ONE 2012, 7, e43062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingston, K.R.; Andrews, J.K.; Harnad, S. Categorical perception effects induced by category learning. J. Exp. Psychol. Learn. Mem. Cogn. 1998, 24, 732. [Google Scholar] [CrossRef] [PubMed]
- Angeli, A.; Davidoff, J.; Valentine, T. Face familiarity, distinctiveness, and categorical perception. Q. J. Exp. Psychol. 2008, 61, 690–707. [Google Scholar] [CrossRef] [Green Version]
- Kikutani, M.; Roberson, D.; Hanley, J.R. What’s in the name? Categorical perception for unfamiliar faces can occur through labeling. Psychon. Bull. Rev. 2008, 15, 787–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenage, S.V. Which twin are you? A demonstration of induced categorical perception of identical twin faces. Br. J. Psychol. 1998, 89, 39–57. [Google Scholar] [CrossRef]
- Viviani, P.; Binda, P.; Borsato, T. Categorical perception of newly learned faces. Vis. Cogn. 2007, 15, 420–467. [Google Scholar] [CrossRef]
- Calder, A.J.; Young, A.W.; Perrett, D.I.; Etcoff, N.L.; Rowland, D. Categorical perception of morphed facial expressions. Vis. Cogn. 1996, 3, 81–118. [Google Scholar] [CrossRef]
- Etcoff, N.L.; Magee, J.J. Categorical perception of facial expressions. Cognition 1992, 44, 227–240. [Google Scholar] [CrossRef]
- Bialystok, E.; Olson, D.R. Spatial categories: The perception and conceptualization of spatial relations. In Categorical Perception: The Groundwork of Cognition; Harnad, S., Ed.; Cambridge University Press: New York, NY, USA, 1987; pp. 511–531. [Google Scholar]
- Eimas, P.D. Infants, speech, and language: A look at some connections. Cognition 1981, 10, 79–84. [Google Scholar] [CrossRef]
- Kuhl, P.K.; Conboy, B.T.; Padden, D.; Nelson, T.; Pruitt, J. Early speech perception and later language development: Implications for the “critical period”. Lang. Learn. Dev. 2005, 1, 237–264. [Google Scholar]
- Kuhl, P.K.; Williams, K.A.; Lacerda, F.; Stevens, K.N.; Lindblom, B. Linguistic experience alters phonetic perception in infants by 6 months of age. Science 1992, 255, 606–608. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, P.K. Innate predispositions and the effects of experience in speech perception: The native language magnet theory. In Developmental Neurocognition: Speech and Face Processing in the First Year of Life; Springer: Dordrecht, The Netherlands, 1993; pp. 259–274. [Google Scholar]
- Quinn, P.C.; Eimas, P.D. On categorization in early infancy. Merrill-Palmer Q. 1986, 32, 331–363. [Google Scholar]
- Lynch, M.P.; Eilers, R.E.; Oller, D.K.; Urbano, R.C. Innateness, experience, and music perception. Psychol. Sci. 1990, 1, 272–276. [Google Scholar] [CrossRef]
- Davidoff, J.; Davies, I.; Roberson, D. Colour categories in a stone-age tribe. Nature 1999, 398, 203–204. [Google Scholar] [CrossRef] [PubMed]
- Massaro, D.W. Categorical perception: Important phenomenon or lasting myth? In Proceedings of the 5th International Congress of Spoken Language Processing, Sydney, Australia, 30 November–4 December 1998. [Google Scholar]
- Searchfield, G.D. Sense and sensibility: A review of the behavioral neuroscience of tinnitus sound therapy and a new typology. In The Behavioral Neuroscience of Tinnitus; Springer: Cham, Switzerland, 2020; pp. 213–247. [Google Scholar]
- Hoare, D.J.; Stacey, P.C.; Hall, D.A. The Efficacy of Auditory Perceptual Training for Tinnitus: A Systematic Review. Ann. Behav. Med. 2010, 40, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jepsen, K.; Sanders, M.; Searchfield, G.; Kobayashi, K. Perceptual training of tinnitus. In Proceedings of the ‘Tinnitus Discovery’: Asia-Pacific Tinnitus Symposium, Auckland, New Zealand, 11–12 September 2009; New Zealand Medical Association: Wellington, New Zealand, 2010; Volume 123, pp. 141–153. [Google Scholar]
- Hemanth, N.; Ghosh, P.G.V. Effect of Frequency Discrimination Training on Tinnitus in Individuals with Flat Sensorineural Hearing loss. Indian J. Otolaryngol. Head Neck Surg. 2021, 74, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Hoare, D.J.; Kowalkowski, V.L.; Hall, D.A. Effects of Frequency Discrimination Training on Tinnitus: Results from Two Randomised Controlled Trials. J. Assoc. Res. Otolaryngol. 2012, 13, 543–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menning, H.; Roberts, L.E.; Pantev, C. Plastic changes in the auditory cortex induced by intensive frequency discrimination training. Neuroreport 2000, 11, 817–822. [Google Scholar] [CrossRef]
- Norena, A.J.; Gourévitch, B.; Aizawa, N.; Eggermont, J.J. Spectrally enhanced acoustic environment disrupts frequency representation in cat auditory cortex. Nat. Neurosci. 2006, 9, 932–939. [Google Scholar] [CrossRef]
- Brown, M.; Irvine, D.R.; Park, V.N. Perceptual learning on an auditory frequency discrimination task by cats: Association with changes in primary auditory cortex. Cereb. Cortex 2004, 14, 952–965. [Google Scholar] [CrossRef]
- Durai, M.; Doborjeh, Z.; Sanders, P.; Vajsakovic, D.; Wendt, A.; Searchfield, G. Behavioral Outcomes and Neural Network Modeling of a Novel, Putative, Recategorization Sound Therapy. Brain Sci. 2021, 11, 554. [Google Scholar] [CrossRef] [PubMed]
- Searchfield, G.D.; Kobayashi, K.; Wise, K.J. Auckland Uniservices Ltd., Assignee. Interactive Gaming System. U.S. Patent 9,808,715, 7 November 2017. [Google Scholar]
- Neal, L. Implications of computer games for system design. In Proceedings of the IFIP TC13 Third International Conference on Human-Computer Interaction, Cambridge, UK, 27–31 August 1990. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vajsakovic, D.; Maslin, M.R.D.; Searchfield, G.D. A Narrative Review of Auditory Categorisation and Its Potential Role in Tinnitus Perception. J. Otorhinolaryngol. Hear. Balance Med. 2022, 3, 6. https://doi.org/10.3390/ohbm3030006
Vajsakovic D, Maslin MRD, Searchfield GD. A Narrative Review of Auditory Categorisation and Its Potential Role in Tinnitus Perception. Journal of Otorhinolaryngology, Hearing and Balance Medicine. 2022; 3(3):6. https://doi.org/10.3390/ohbm3030006
Chicago/Turabian StyleVajsakovic, Dunja, Michael R. D. Maslin, and Grant D. Searchfield. 2022. "A Narrative Review of Auditory Categorisation and Its Potential Role in Tinnitus Perception" Journal of Otorhinolaryngology, Hearing and Balance Medicine 3, no. 3: 6. https://doi.org/10.3390/ohbm3030006
APA StyleVajsakovic, D., Maslin, M. R. D., & Searchfield, G. D. (2022). A Narrative Review of Auditory Categorisation and Its Potential Role in Tinnitus Perception. Journal of Otorhinolaryngology, Hearing and Balance Medicine, 3(3), 6. https://doi.org/10.3390/ohbm3030006