
Understanding the Dynamics of a Lipid Monolayer on a Water Surface under a Marangoni Flow

Abstract
:
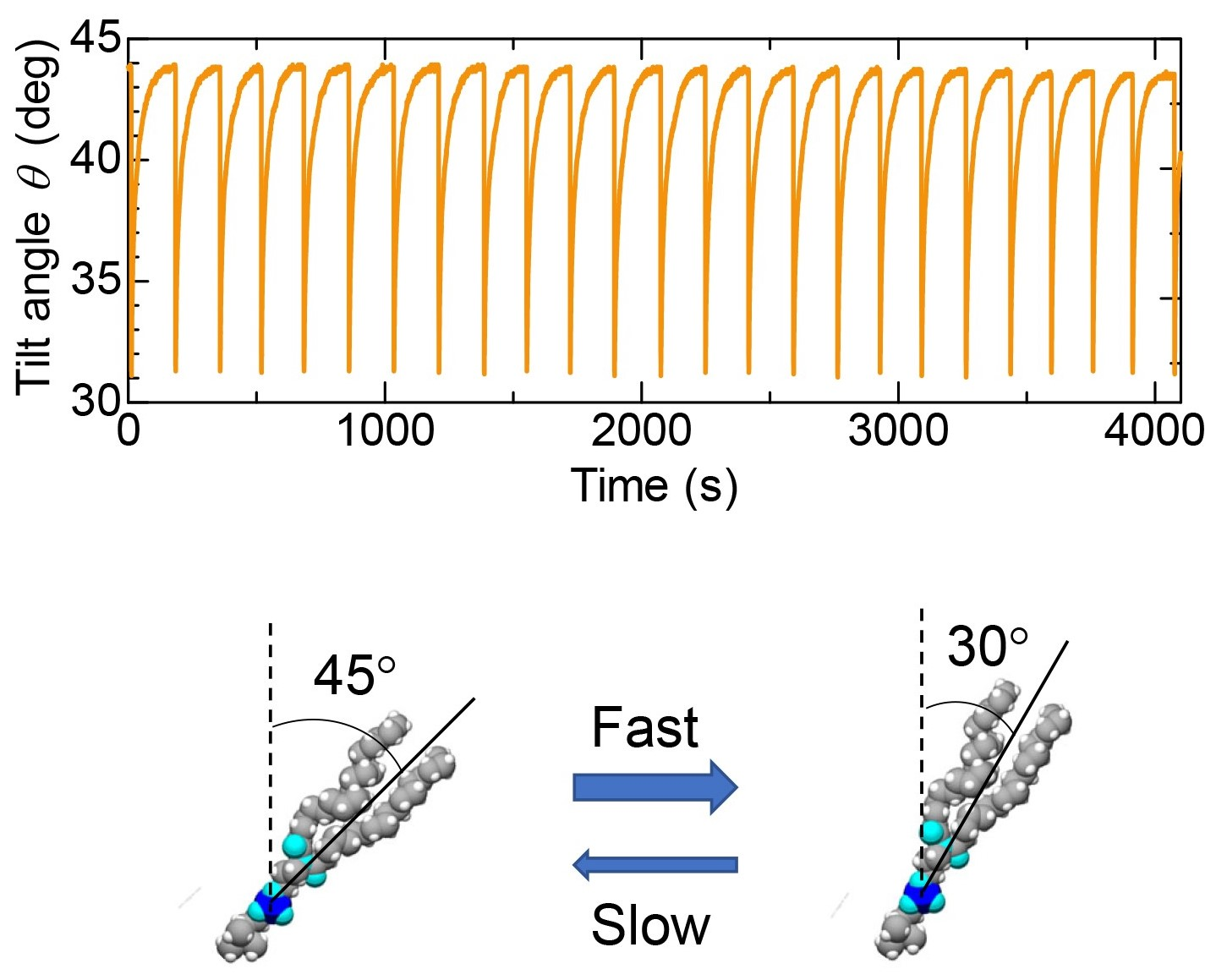{kind=link}
{kind=link}
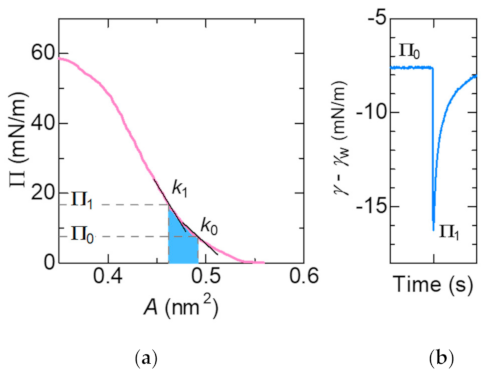{kind=link}
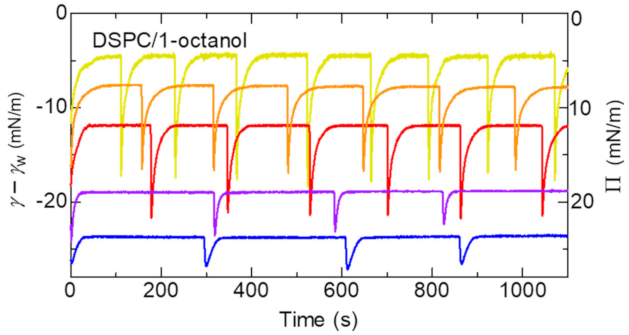{kind=link}
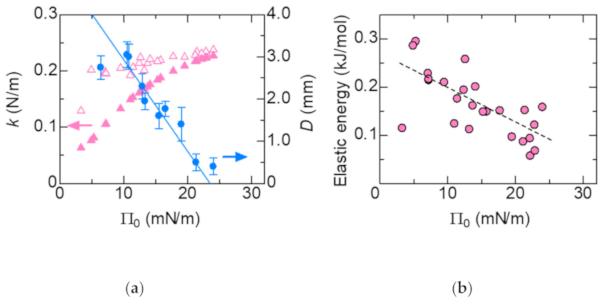{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Experimental Methods
2.1.1. Materials
2.1.2. Experimental Setup
2.1.3. X-ray Reflectometry Measurements for Lipid Monolayers under Static Condition
2.2. Data Analysis
2.2.1. Elasticity of the Lipid Monolayer
2.2.2. X-Ray Reflectometry
3. Results
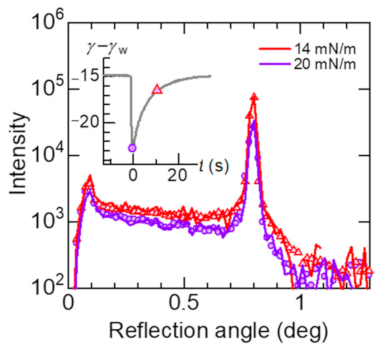
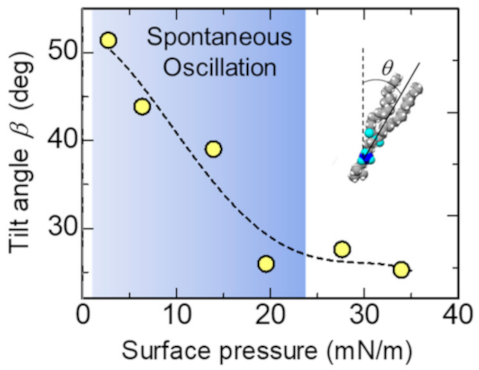
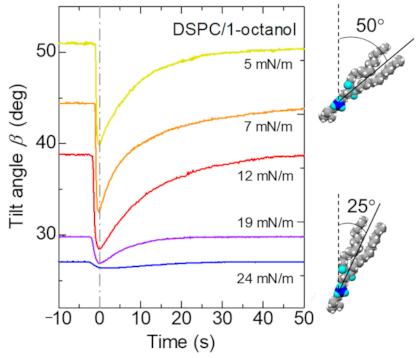
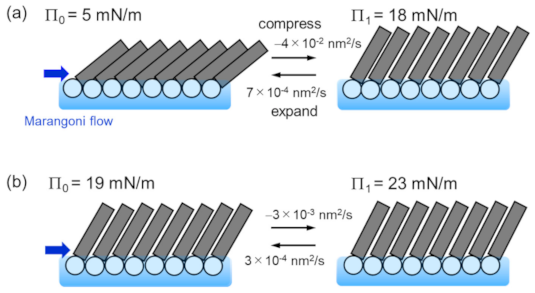
3.1. Elastic Motion of the Lipid Monolayer
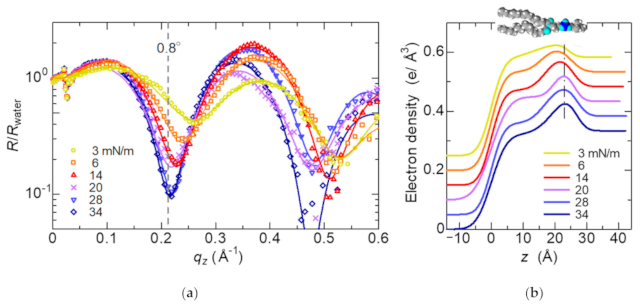
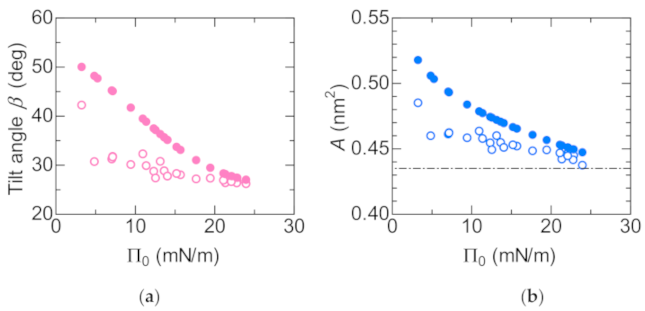
3.2. Structure of the Lipid Monolayer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lingwood, D.; Simons, K. Lipid Rafts as a Membrane-Organizing Principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The Mystery of Membrane Organization: Composition, Regulation and Roles of Lipid Rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, M.S.; Sens, P.; Socci, N.D. Nonequilibrium Raftlike Membrane Domains under Continuous Recycling. Phys. Rev. Lett. 2005, 95, 3–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinosa, G.; López-Montero, I.; Monroya, F.; Langevin, D. Shear Rheology of Lipid Monolayers and Insights on Membrane Fluidity. Proc. Natl. Acad. Sci. USA 2011, 108, 6008–6013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scriven, L.E. The Marangoni Effects. Nature 1960, 187, 186. [Google Scholar] [CrossRef]
- Kovalchuk, V.I.; Kamusewitz, H.; Vollhardt, D.; Kovalchuk, N.M. Auto-Oscillation of Surface Tension. Phys. Rev. E Stat. Phys. PlasmasFluidsRelat. Interdiscip. Top. 1999, 60, 2029–2036. [Google Scholar] [CrossRef] [PubMed]
- Grigorieva, O.v.; Kovalchuk, N.M.; Grigoriev, D.O.; Vollhardt, D. Spontaneous Non-Linear Surface Tension Oscillations in the Presence of a Spread Surfactant Monolayer at the Air/Water Interface. Colloids Surf. A Physicochem. Eng. Asp. 2004, 250, 141–151. [Google Scholar] [CrossRef]
- Kovalchuk, N.M. Spontaneous Oscillations Due to Solutal Marangoni Instability: Air/Water Interface. Cent. Eur. J. Chem. 2012, 10, 1423–1441. [Google Scholar] [CrossRef]
- Kovalchuk, N.M.; Vollhardt, D. Autooscillations of Surface Tension in Water-Alcohol Systems. J. Phys. Chem. B 2000, 104, 7987–7992. [Google Scholar] [CrossRef]
- Kovalchuk, N.M.; Vollhardt, D. A Numerical Study of Surface Tension Auto-Oscillations. Effect of Surfactant Properties. J. Phys. Chem. B 2001, 105, 4709–4714. [Google Scholar] [CrossRef]
- Kovalchuk, N.M.; Vollhardt, D. Theoretical Description of Repeated Surface-Tension Auto-Oscillations. Phys. Rev. E Stat. Phys. PlasmasFluidsRelat. Interdiscip. Top. 2002, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yano, Y.F.; Tada, H.; Arakawa, E.; Voegeli, W.; Ina, T.; Uruga, T.; Matsushita, T. Periodic Elastic Motion in a Self-Assembled Monolayer under Spontaneous Oscillations of Surface Tension: Molecules in a Scrum Push Back a Marangoni Flow. J. Phys. Chem. Lett. 2020, 11, 6330–6336. [Google Scholar] [CrossRef] [PubMed]
- Yano, Y.F.; Uruga, T.; Tanida, H.; Toyokawa, H.; Terada, Y.; Yamada, H. Simultaneous Measurement of X-Ray Specular Reflection and off-Specular Diffuse Scattering from Liquid Surfaces Using a Two-Dimensional Pixel Array Detector: The Liquid-Interface Reflectometer of BL37XU at SPring-8. J. Synchrotron Radiat. 2010, 17, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaines, G.L. Insoluble Monolayers at Liquid-Gas Interfaces; Interscience Publishers: New York, NY, USA, 1966. [Google Scholar]
- Miyoshi, T.; Kato, S. Detailed Analysis of the Surface Area and Elasticity in the Saturated 1,2-Diacylphosphatidylcholine/Cholesterol Binary Monolayer System. Langmuir 2015, 31, 9086–9096. [Google Scholar] [CrossRef] [PubMed]
- Pershan, P.S.; Schlossman, M. Liquid Surfaces and Interfaces: Synchrotron X-Ray Methods; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Skoda, M.W.A. Recent Developments in the Application of X-Ray and Neutron Reflectivity to Soft-Matter Systems. Curr. Opin. Colloid Interface Sci. 2019, 42, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Schalke, M.; Lösche, M. Structural Models of Lipid Surface Monolayers from X-Ray and Neutron Reflectivity Measurements. Adv. Colloid Interface Sci. 2000, 88, 243–274. [Google Scholar] [CrossRef]
- Helm, C.A.; Möhwald, H.; Kjör, K.; Als-Nielsen, J. Phospholipid Monolayer Density Distribution Perpendicular to the Water Surface. A Synchrotron x-Ray Reflectivity Study. Europhys. Lett. 1987, 4, 697–703. [Google Scholar] [CrossRef]
- Nagle, J.F.; Wilkinson, D.A. Lecithin Bilayers. Density Measurement and Molecular Interactions. Biophys. J. 1978, 23, 159–175. [Google Scholar] [CrossRef] [Green Version]
- Marsh, D. Lateral Pressure in Membranes. Biochim. Biophys. Acta-Rev. Biomembr. 1996, 1286, 183–223. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yano, Y.F.; Ina, T.; Uruga, T. Understanding the Dynamics of a Lipid Monolayer on a Water Surface under a Marangoni Flow. Colloids Interfaces 2021, 5, 31. https://doi.org/10.3390/colloids5020031
Yano YF, Ina T, Uruga T. Understanding the Dynamics of a Lipid Monolayer on a Water Surface under a Marangoni Flow. Colloids and Interfaces. 2021; 5(2):31. https://doi.org/10.3390/colloids5020031
Chicago/Turabian StyleYano, Yohko F., Toshiaki Ina, and Tomoya Uruga. 2021. "Understanding the Dynamics of a Lipid Monolayer on a Water Surface under a Marangoni Flow" Colloids and Interfaces 5, no. 2: 31. https://doi.org/10.3390/colloids5020031
APA StyleYano, Y. F., Ina, T., & Uruga, T. (2021). Understanding the Dynamics of a Lipid Monolayer on a Water Surface under a Marangoni Flow. Colloids and Interfaces, 5(2), 31. https://doi.org/10.3390/colloids5020031