
The Latest Occurrence of Stephanorhinus hundsheimensis (Rhinocerotidae) in Europe: The Skeletons from the Cova del Rinoceront Site (Castelldefels, Barcelona)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
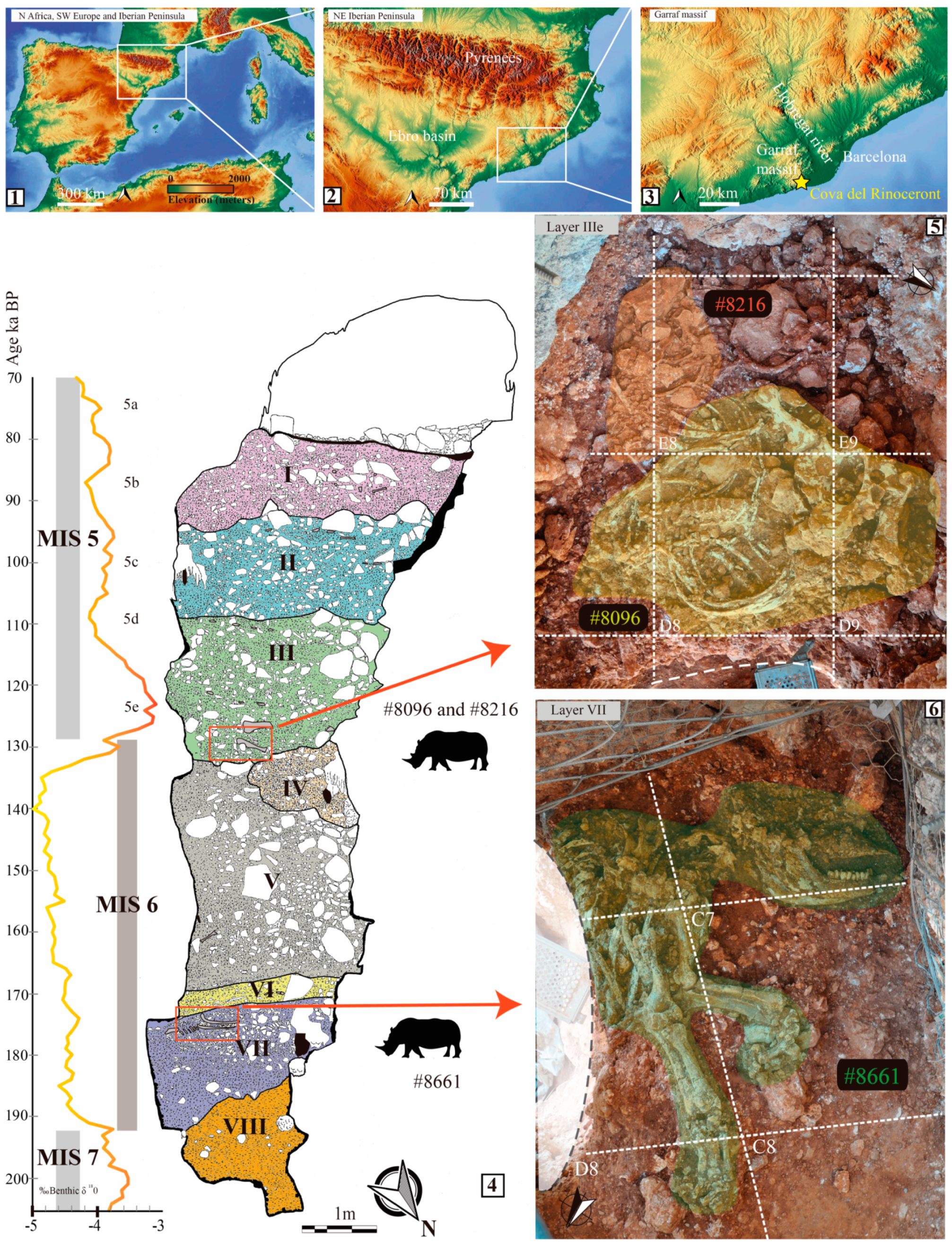
2. Cova del Rinoceront
2.1. Site Description
2.2. Stratigraphic Provenience and Chronological Framework of Rhinocerotid Remains
3. Materials and Methods
Anatomical Abbreviations
4. Systematic Palaeontology
4.1. Referred Material
4.2. Description and Comparison
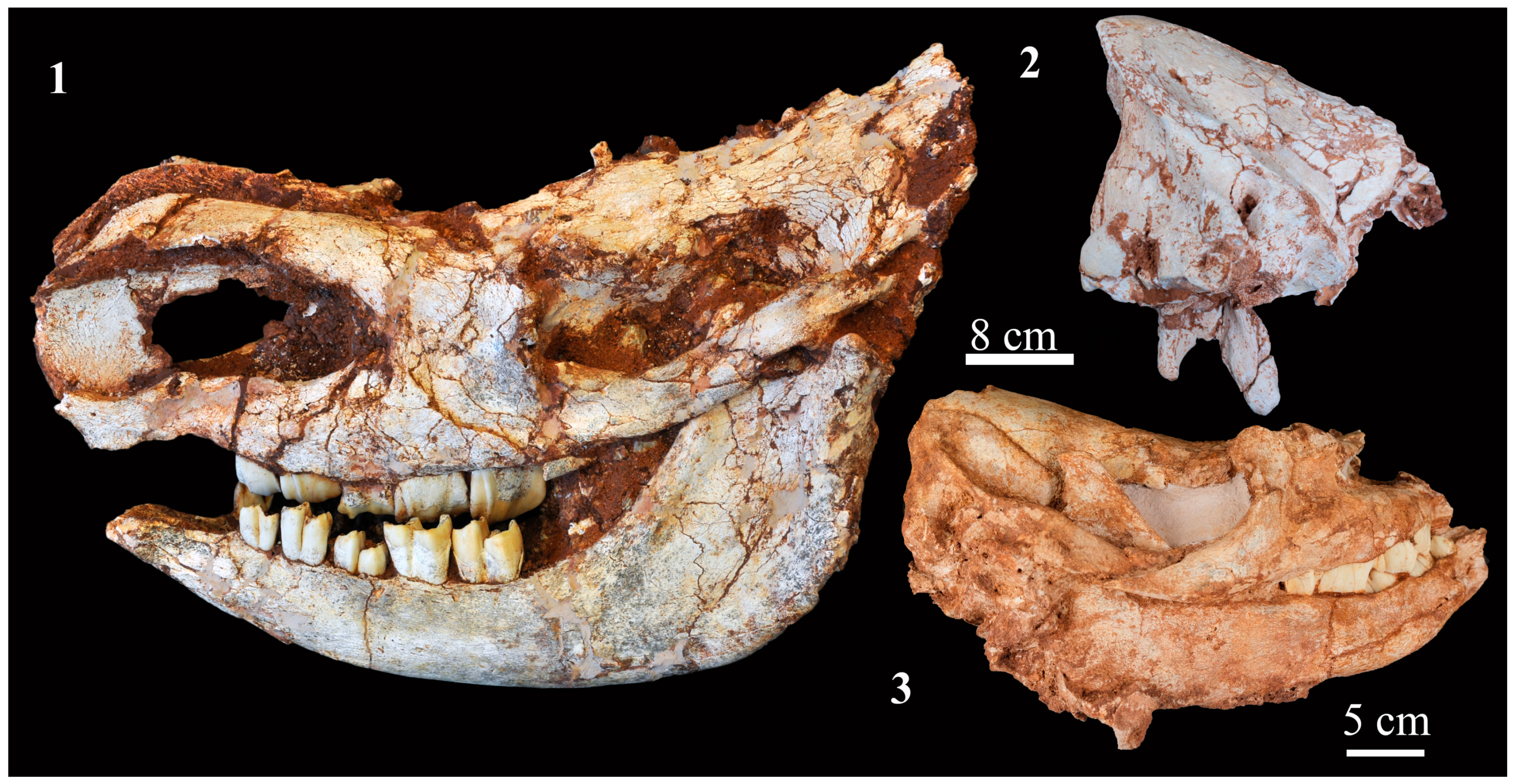
4.2.1. Skull
4.2.2. Mandible
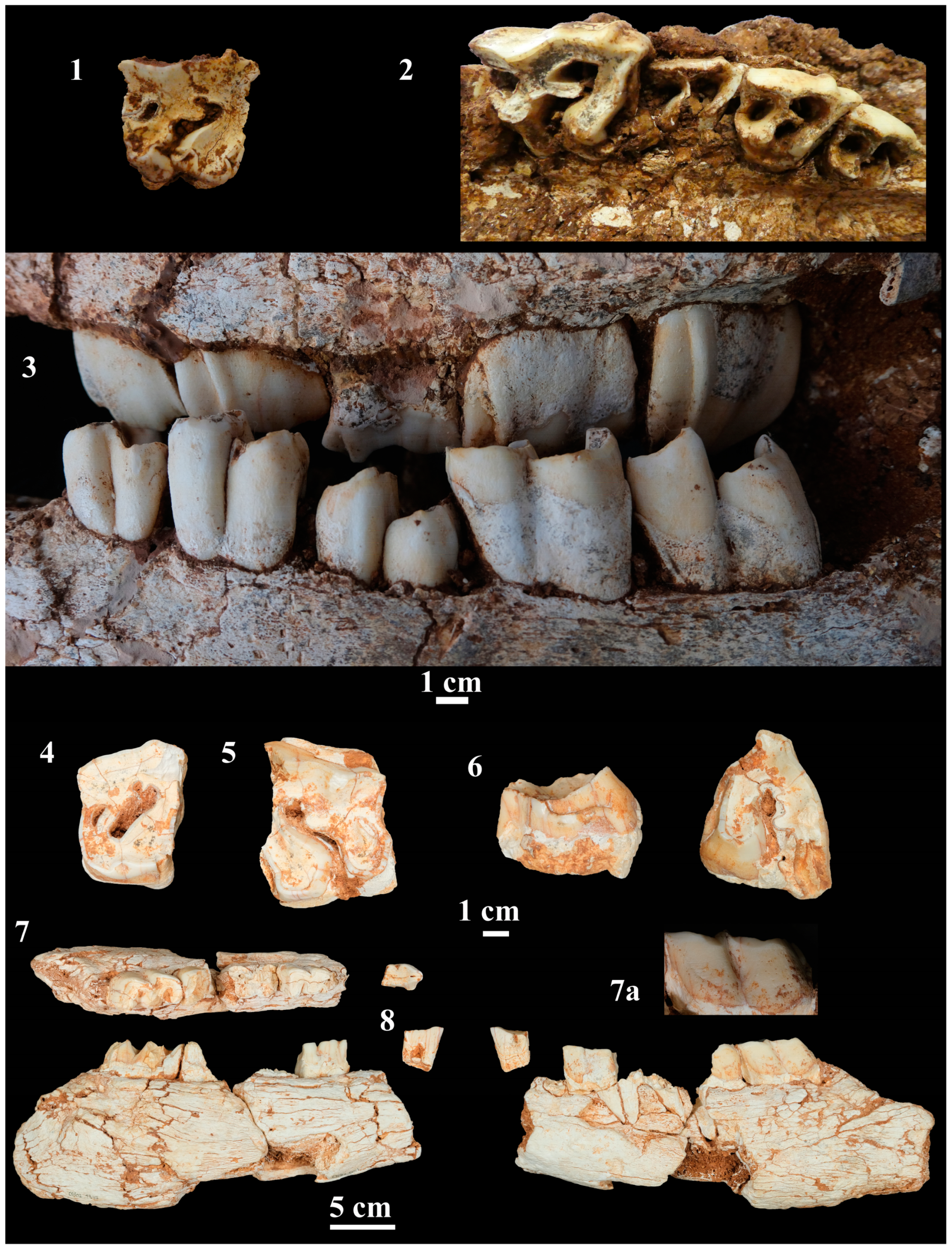
4.2.3. Upper Dentition
4.2.4. Lower Dentition
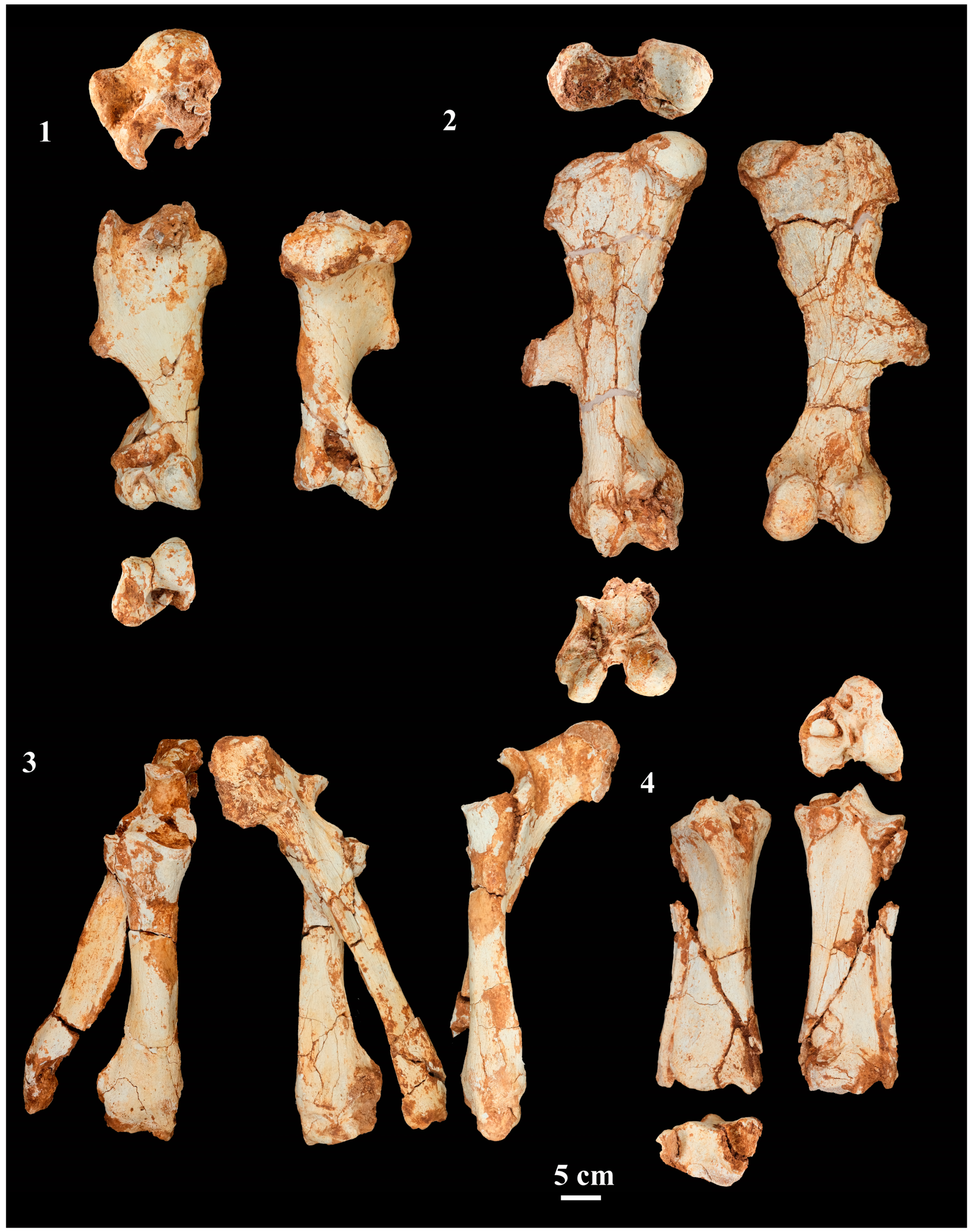
4.2.5. Fore- and Hindlimb Long Bones
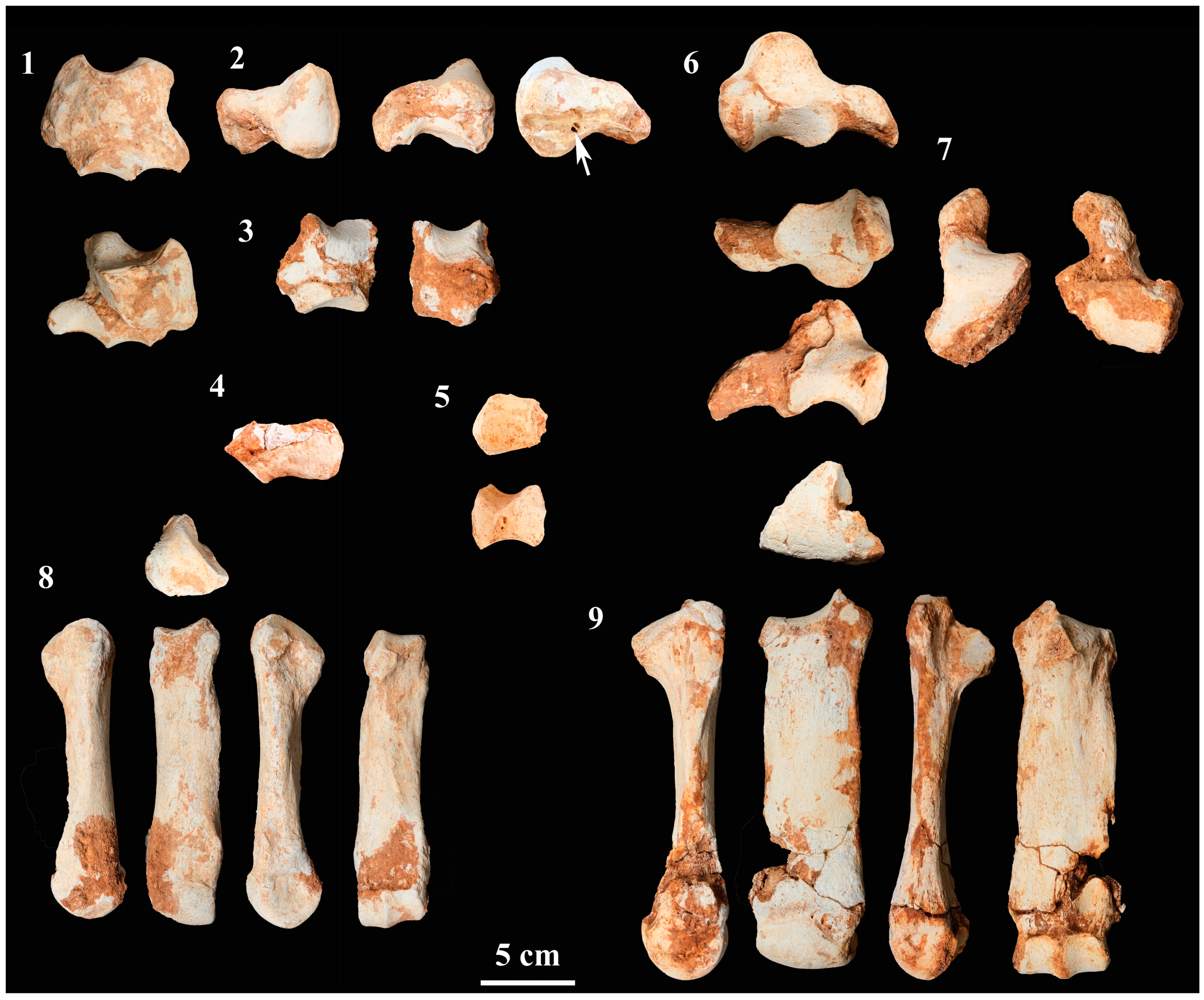
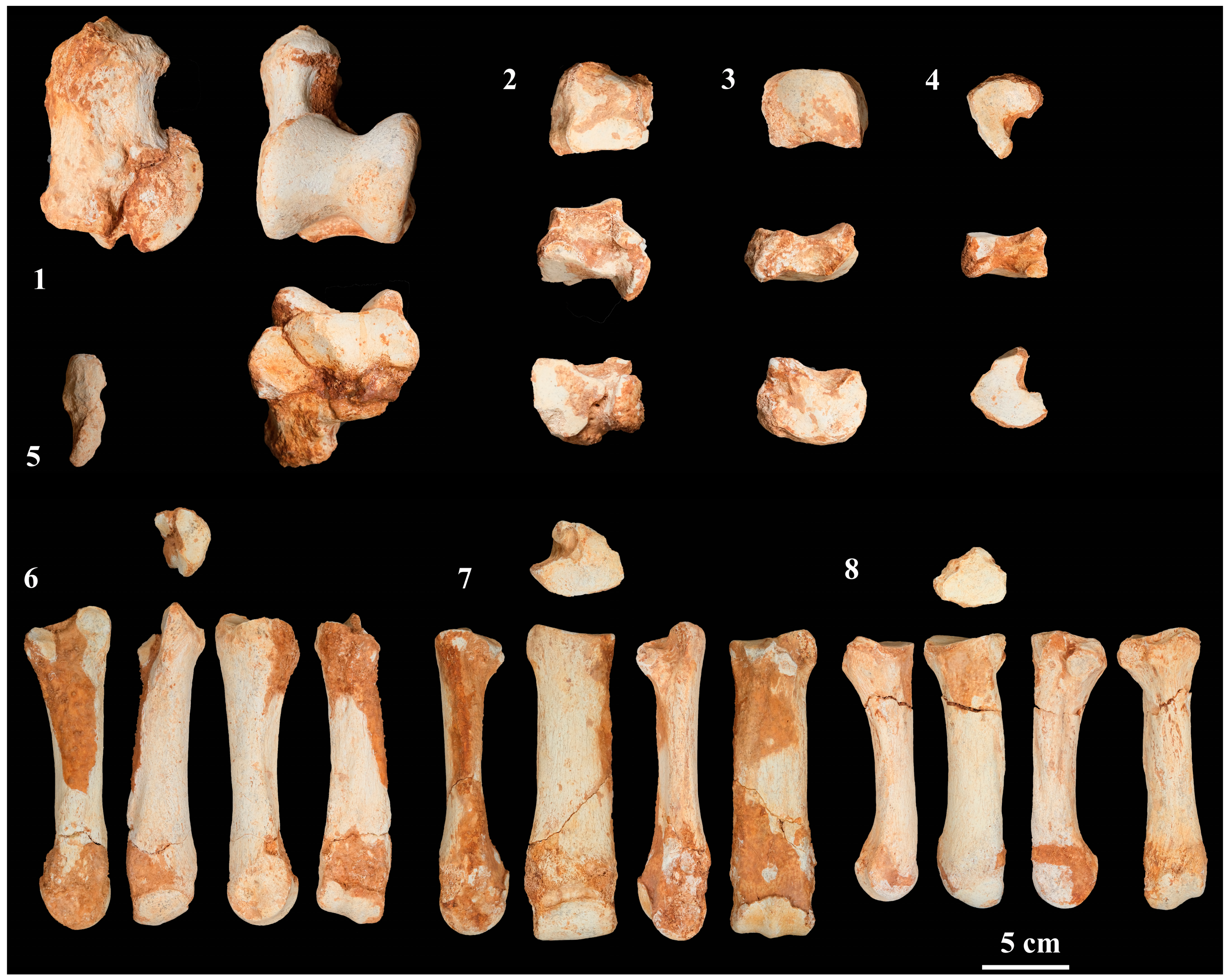
4.2.6. Carpal Bones
4.2.7. Metacarpals
4.2.8. Tarsal Bones
4.2.9. Metatarsals
4.2.10. Phalanges
5. Discussion
5.1. Taxonomy
5.2. Biostratigraphy
5.3. Geographical Distribution
5.4. Palaeobiological Remarks
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pandolfi, L. Sistematica e Filogenesi dei Rhinocerotini (Mammalia, Rhinocerotidae). Ph.D. Dissertation, Scuola Dottorale in Geologia dell’Ambiente e delle Risorse, Università degli Studi Roma Tre, Roma, Italy, 2015; p. 338. [Google Scholar]
- Pandolfi, L.; Erten, H. Stephanorhinus hundsheimensis (Mammalia, Rhinocerotidae) from the late early Pleistocene deposits of the Denizli Basin (Anatolia, Turkey). Geobios 2017, 50, 65–73. [Google Scholar] [CrossRef]
- Cerdeño, E. Stephanorhinus hemitoechus (Falc.) (Rhinocerotidae, Mammalia) del Pleistoceno medio y superior de España. Est. Geol. 1990, 46, 465–479. [Google Scholar] [CrossRef]
- Riquelme Cantal, J.A.; Barroso Ruiz, C.; Botella Ortega, D.; Caparrós, M.; Moine, A.M.; García Solano, J.A. Un yacimiento del Pleistoceno medio y superior en el sur de la Península Ibérica: La Cueva del Ángel (Lucena, Córdoba). CPAG 2010, 20, 201–221. [Google Scholar]
- Pandolfi, L.; Marra, F. Rhinocerotidae (Mammalia, Perissodactyla) from the chrono-stratigraphically constrained Pleistocene deposits of the urban area of Rome (Central Italy). Geobios 2015, 699, 147–167. [Google Scholar] [CrossRef]
- Zilhão, J.; Angelucci, D.E.; Arnold, L.J.; Demuro, M.; Hoffmann, D.L.; Pike, A.W.G. A revised, Last Interglacial chronology for the Middle Palaeolithic sequence of Gruta da Oliveira (Almonda karst system, Torres Novas, Portugal). Quat. Sci. Rev. 2021, 258, 106885. [Google Scholar] [CrossRef]
- Zilhão, J.; Angelucci, D.E.; Araújo Igreja, M.; Arnold, L.J.; Badal, E.; Callapez, P.; Cardoso, J.L.; d’Errico, F.; Daura, J.; Demuro, M.; et al. Last Interglacial Iberian Neandertals as fisher-hunter-gatherers. Science 2020, 367, eaaz7943. [Google Scholar] [CrossRef] [PubMed]
- Demuro, M.; Arnold, L.J.; Aranburu, A.; Gómez-Olivencia, A.; Arsuaga, J.L. Single-grain OSL dating of the Middle Palaeolithic site of Galería de las Estatuas, Atapuerca (Burgos, Spain). Quat. Geochr. 2019, 49, 254–261. [Google Scholar] [CrossRef]
- Arsuaga, J.L.; Baquedano, E.; Pérez-González, A.; Sala, M.T.; Quam, R.M.; Rodríguez, L.; García, R.; García, N.; Álvarez-Lao, D.; Laplana, C.; et al. Understanding the ancient habitats of the last-interglacial (late MIS 5) Neanderthals of central Iberia: Paleoenvironmental and taphonomic evidence from the Cueva del Camino (Spain) site. Quat. Intern. 2012, 275, 55–75. [Google Scholar] [CrossRef]
- Daura, J.; Sanz, M.; Julià, R.; García-Fernández, D.; Fornós, J.J.; Vaquero, M.; Allué, M.; López-García, J.M.; Blain, H.A.; Ortiz, J.E.; et al. Cova del Rinoceront (Castelldefels, Barcelona): A terrestrial record for the Last Interglacial period (MIS 5) in the Mediterranean coast of the Iberian Peninsula. Quat. Sci. Rev. 2015, 114, 203–227. [Google Scholar] [CrossRef]
- Pandolfi, L.; Tagliacozzo, A. Stephanorhinus hemitoechus (Mammalia, Rhinocerotidae) from the Late Pleistocene of Valle Radice (Sora, Central Italy) and re-evaluation of the morphometric variability of the species in Europe. Geobios 2015, 48, 169–191. [Google Scholar] [CrossRef]
- Van der Made, J. The rhinos from the Middle Pleistocene of Neumark-Nord (Saxony-Anhalt). Veröffentlich. Land. Denkm. Archäol. 2010, 62, 433–527. [Google Scholar]
- Álvarez-Lao, D.J.; García, N. Southern dispersal and Palaeoecological implications of woolly rhinoceros (Coelodonta antiquitatis): Review of the Iberian occurrences. Quat. Sci. Rev. 2011, 30, 2002–2017. [Google Scholar] [CrossRef]
- Alba, D.M.; Aurell, J.; Madurell, J.; Gómez, M.; Moyà-Solà, S.; Berástegui, X. Paleontologia i Geologia del jaciment del Pleistocè inferior de Vallparadís (Terrassa, Vallès Occidental). Trib. d’Arqueol. 2008, 2008, 29–44. [Google Scholar]
- Madurell-Malapeira, J.; Minwer-Barakat, R.; Alba, D.M.; Garcés, M.; Gómez, M.; Aurell-Garrido, J.; Ros-Montoya, S.; Moyà-Solà, S.; Berástegui, X. The Vallparadís section (Terrassa, Iberian Peninsula) and the latest Villafranchian faunas of Europe. Quat. Sci. Rev. 2010, 29, 3972–3982. [Google Scholar] [CrossRef]
- Madurell-Malapeira, J.; Ros-Montoya, M.P.; Espigares, D.M.; Alba, J.; Aurell-Garrido, J. Villafranchian large mammals from the Iberian Peninsula: Paleobiography, paleoecology and dispersal events. J. Iber. Geol. 2014, 40, 167–178. [Google Scholar] [CrossRef]
- Alba, D.M.; Madurell-Malapeira, J.; Nelson, E.; Vinuesa, V.; Susana, I.; Patrocinio Espigares, M.; Ros-Montoya, S.; Martínez-Navarro, B. First record of macaques from the Early Pleistocene of Incarcal (NE, Iberian Peninsula). J. Hum. Evol. 2016, 96, 139–144. [Google Scholar] [CrossRef]
- García-Fernández, D.; Galobart, A.; Ros, X.; Cerdeño, E. Stephanorhinus etruscus (Rhinocerotidae) en el Villafranquiense de Crespià (Girona, NE de la Península Ibérica). Paleontol. Evol. 2001, 34, 279–296. [Google Scholar]
- García-Fernández, D.; Galobart, A.; Cerdeño, E. Perisodáctilos del Pleistoceno inferior de los yacimientos de Incarcal (Girona, NE de la Península Ibérica). Paleontol. Evol. 2003, 34, 175–183. [Google Scholar]
- Cerdeño, E. Remarks on the Spanish Plio-Pleistocene rhinocerotid Stephanorhinus etruscus. Comptes Rendus Acad. Sci. Paris 1993, 317, 1363–1367. [Google Scholar]
- Ros Montoya, S. Los Proboscideos del Plio-Pleistoceno de las Cuencas de Guádix-Baza y Granada. Doctoral Thesis, Departamento de Estratigrafía y Paleontología, Universidad de Granada, Granada, Spain, 2010. [Google Scholar]
- Lacombat, F. Estudio paleontológico de Stephanorhinus hundsheimensis de Fuente Nueva 3 y Barranco León. In Ocupaciones Humanas en el Pleistoceno Inferior y Medio de la Cuenca Guadix-Baza; Toro, I., Martínez-Navarro, B., Agustí, J., Eds.; Junta de Andalucía, Arqueología-Monografías: Sevilla, Spain, 2010; pp. 237–246. [Google Scholar]
- Duval, M.; Falgueres, C.; Bahain, J.J.; Rainer Gru, N.; Qinfeng, S.; Maxime, A.; Hellstrom, J.; Dolo, J.M.; Agustí, J.; Martínez-Navarro, B.; et al. The challenge of dating Early Pleistocene fossil teeth by the combined uranium series–electron spin resonance method: The Venta Micena palaeontological site (Orce, Spain). J. Quat. Sci. 2011, 26, 603–615. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, G.; Rodríguez, J.; Mateos, A.; Martín-González, J.A.; Goikoetxea, I. Food Web Structure during the European Pleistocene. J. Taphon. 2012, 10, 165–184. [Google Scholar]
- Daura, J.; Sanz, M. El jaciment prehistòric de la pedrera de Ca n’Aymerich de Castelldefels. IV Trobades d’Estudiosos del Massís del Garraf (21 de novembre de 2002, Vilanova i al Geltrú). Diput. Barc. Serv. Parcs Nat. Monogr. 2004, 37, 165–168. [Google Scholar]
- Sanz, M.; Daura, J. Taphonomic analysis of an ungulate-dominated accumulation at the Pleistocene Cova del Rinoceront site near Barcelona, Spain (northeastern Iberian Peninsula). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 498, 24–38. [Google Scholar] [CrossRef]
- Croitor, R.; Sanz, M.; Daura, J. The endemic deer Haploidoceros mediterraneus (Bonifay) (Cervidae, Mammalia) from the Late Pleistocene of Cova del Rinoceront (Iberian Peninsula): Origin, ecomorphology, and paleobiology. Hist. Biol. 2020, 32, 409–427. [Google Scholar] [CrossRef]
- Palombo, M.R.; Sanz, M.; Daura, J. The complete skeleton of a straight-tusked elephant calf from Cova del Rinoceront (Late Pleistocene, NE Iberian Peninsula): New insights into ontogenetic growth in Palaeoloxodon antiquus. Quat. Sci. Rev. 2021, 274, 107257. [Google Scholar] [CrossRef]
- Guérin, C. Les Rhinocerotidae (Mammalia-Perissodactyla) du Miocene Terminal au Pléistocène supérieur d’Europe Occidentale comparación avec les especes actuelles. Doc. Lab. Géol. Lyon 1980, 79, 1–1185. [Google Scholar]
- Fortelius, M.; Mazza, P.; Sala, B. Stephanorhinus (Mammalia-Rhinocerotidae) of the Western European Pleistocene, with a revision of S. etruscus (Falconer, 1868). Palaeontogr. Ital. 1993, 80, 63–155. [Google Scholar]
- Lacombat, F. Les Rhinocéros Fossiles des Sites Préhistoriques de l’Europe Méditerranéene et du Massif Central, Paléontologie et Implications Bichronologiques; Bristish Archaeological Reports, International Series, 1419; BAA: Oxford, UK, 2005; pp. 1–175. [Google Scholar]
- Geraads, D.; Cerdeño, E.; García Fernández, D.; Pandolfi, L.; Billia, E.; Athanassiou, A.; Albayrak, E.; Codrea, V.; Obada, T.; Deng, T.; et al. A Database of Old-World Neogene and Quaternary Rhino-Bearing Localities. 2020. Available online: http://www.rhinoresourcecenter.com/about/fossil-rhino-database.php (accessed on 15 February 2023).
- von Jäger, G.F. Über die Fossilen Säugethiere, Welche in Württemberg in Verschiedenen Formationen Aufgefunden Worden Sind, Nebst Geognostichen Bemerkungen über Diese Formationen; Carl Erhard: Stuttgart, Germany, 1839; p. 214. [Google Scholar]
- Toula, F. Das Nashorn von Hundsheim Rhinoceros (Ceratorhinus Osborn) Hundsheimensis nov. form. Mit Ausführunger über die Verhältnisse von elf Schädeln von Rhinoceros (Ceratorhinus) sumatrensis. Abh. KK Geol. Reich. 1902, 19, 1–92. [Google Scholar]
- Mazza, P. The Tuscan Early Pleistocene Dicerorhinus etruscus. Palaeontogr. Ital. 1988, 75, 1–87. [Google Scholar]
- Azzaroli, A. Validita della specie Rhinoceros hemitoechus Falc. Palaeont. Ital. 1962, 57, 21–34. [Google Scholar]
- Pandolfi, L.; Bartolini-Lucenti, S.; Cirilli, O.; Bukhsianidze, M.; Lordkipanidze, D.; Rook, L. Paleoecology, biochronology, and paleobiogeography of Eurasian Rhinocerotidae during the Early Pleistocene: The contribution of the fossil material from Dmanisi (Georgia, Southern Caucasus). J. Hum. Evol. 2021, 156, 103013. [Google Scholar] [CrossRef] [PubMed]
- Kotowski, A.; Stefaniak, K.; Kahlke, R.-D. A rhinocerotid skull from the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld near Meiningen (Thüringen, Germany); Kahlke, R.-D., Ed.; Römisch-Germanisches Zentralmuseum: Mainz, Germany, 2020; Part 4; pp. 273–1294. [Google Scholar]
- Loose, H.K. Pleistocene Rhinocerotidae of W. Europe with reference to the recent two-horned species of Africa and S.E. Asia. Scr. Geol. 1975, 33, 1–60. [Google Scholar]
- Schreiber, H.D. Osteological investigations on skeleton material of Rhinoceroses (Rhinocerotidae, Mammalia) from the Early Middle Pleistocene locality of Mauer near Heildelberg (SW-Germany). Quat. Hors Série 2005, 2, 103–111. [Google Scholar]
- Cigala-Fulgosi, F. Dicerorhinus hemitoechus (Falconer) del post-villafranchiano fluviolacustre del Torrente Stirone (Salsomagggiore, Parma). Boll. Soc. Paleontol. Ital. 1976, 15, 59–72. [Google Scholar]
- Bona, F.; Sala, B. Villafranchian-Galerian mammal faunas transition in South-Western Europe. The case of the Late Early Pleistocene mammal fauna of the Frantoio locality, Arda River (Castell’Arquato, Piacenza, Northern Italy). Geobios 2016, 49, 329–347. [Google Scholar] [CrossRef]
- Pandolfi, L.; Gaeta, M.; Petronio, C. The skull of Stephanorhinus hemitoechus (Mammalia, Rhinocerotidae) from the Middle Pleistocene of Campagna Romana (Rome, central Italy): Biochronological and paleobiogeographic implications. Bull. Geosci. 2013, 88, 51–62. [Google Scholar] [CrossRef]
- Schroeder, H. Die Wirbelthier-Fauna des Mosbacher Sandes. 1. Gattung Rhinoceros. Abh. Kön. Preuss. Geol. Landes. 1903, 18, 1–143. [Google Scholar]
- Kahlke, H.D. Die Rhinocerotiden-Reste aus den Tonen von Voigtstedt in Thüringen. Palaeontol. Abh. A 1965, 2, 451–520. [Google Scholar]
- Sala, B.; Fortelius, M. The Rhinoceros of Isernia La Pineta (early Middle Pleistocene, Southern Italy). Palaeontogr. Ital. 1993, 80, 157–174. [Google Scholar]
- Mazza, P. The Middle Pleistocene rhinoceros remains from Cesi (Colfiorito Basin, Macerata, Central Italy). Boll. Soc. Palaeontol. Ital. 1996, 35, 349–355. [Google Scholar]
- Kahlke, H.D. Die Rhinocerotiden-Reste aus den Kiesen von Süssenborn bei Weimar. Palaeontol. Abh. A 1969, 3, 667–709. [Google Scholar]
- Kahlke, H.D. Die Rhinocerotiden-Reste aus dem unterpleistozän von Untermassfeld. In Das Pleistozän von Untermassfeld bei Meiningen, Thüringen; Kahlke, R.-D., Ed.; Monographien Römisch-Germanischen Zentralmus: Mainz, Germany, 2001; Volume 40, pp. 501–556. [Google Scholar]
- Moullé, P.E.; Lacombat, F.; Echassoux, A. Apport des grands mammifères de la Grotte du Vallonnet (Roquebrune-Cap Martin, Alpes Maritimes, France) à la connaissance du cadre biochronologique de la seconde moitié du Pléistocène inférieur d’Europe. L’Anthropologie 2006, 110, 837–849. [Google Scholar] [CrossRef]
- Ballatore, M.; Breda, M. Stephanorhinus hundsheimensis (Rhinocerotidae, Mammalia) teeth from the early Middle Pleistocene of Isernia La Pineta (Molise, Italy) and comparison with coeval British material. Quat. Intern. 2013, 302, 169–183. [Google Scholar] [CrossRef]
- Vislobokova, I.A.; Agadjanian, A.K. New data on large mammals of the Pleistocene Trlica Fauna, Montenegro, the Central Balkans. Paleontol. J. 2015, 49, 651–667. [Google Scholar] [CrossRef]
- Cerdeño, E. Presencia de rinoceronte en la fauna de Cueva Millán (Burgos). Geogaceta 1987, 2, 9–10. [Google Scholar]
- Lacombat, F. Morphological and biometrical differentiation of the teeth from the Pleistocene species of Stephanorhinus (Mammalia, Perissodactyla, Rhinocerotidae) in Mediterranean Europe and the Massif Central, France. Palaeontographica A 2006, 274, 71–111. [Google Scholar] [CrossRef]
- Alférez, F.; Íñigo, C. Los restos de Dicerorhinus hemitoechus (Perissodactyla; Mammalia) del Pleistoceno Medio de Pinilla del Valle (Madrid). In Actas de Paleontología; Civis Llovera, J., Flores Villarejo, A., Eds.; Universidad de Salamanca: Salamanca, Spain, 1990; pp. 25–40. [Google Scholar]
- Íñigo, C. El rinoceronte del Pleistoceno Superior de la Cueva del Búho (Segovia). Bol. Geol. Min. 1995, 106, 107–110. [Google Scholar]
- Van der Made, J.; Montoya, P. Rinocerontes del Pleistoceno de El Bardello, Plaça de La República y El Molinar en Alcoy, España. Rev. Mus. Alcoi 2007, 16, 7–18. [Google Scholar]
- Abdala, V.; Vera, M.C.; Amador, L.I.; Fontanarrosa, G.; Fratani, J.; Ponssa, M.L. Sesamoids in tetrapods: The origin of new skeletal morphologies. Biol. Rev. 2019, 94, 2011–2032. [Google Scholar] [CrossRef]
- Baryshnikov, G.; Guérin, C.; Mezhlumyan, S.K. Rhinoceros Dicerorhinus etruscus brachycephalus from the Yerevan Mousterian site. Proc. Zool. Inst. 1989, 198, 103–110. [Google Scholar]
- Santafé, J.V.; Casanovas, M.L. Dicerorhinus hemitoechus (Falconer, 1868) (Mammalia, Perissodactyla) del yacimiento Pleistocénico de la Cova del Gegant (Garraf, Barcelona). Empúries 1986, 48–50, 310–322. [Google Scholar]
- Mazza, P.; Sala, B.; Fortelius, M. A small latest Villafranchian (late Early Pleistocene) rhinoceros from Pietrafitta (Perugia, Umbria, Central Italy) with notes on the Pirro and Westerhoven rhinoceroses. Palaeontogr. Ital. 1993, 8, 25–50. [Google Scholar]
- Kaniewski, D.; Renault-Miskovskiy, J.; de Lumley, H. Palaeovegetation from a Homo neanderthalensis occupation in Western Liguria: Archaeopalynology of Madonna dell’Arma (San Remo, Italy). J. Archaeol. Sci. 2005, 32, 827–840. [Google Scholar] [CrossRef]
- Berto, C.; Rubinato, G. The upper Pleistocene mammal record from Caverna degli Orsi (San Dorligo della Valle–Dolina, Trieste, Italy): A faunal complex between eastern and western Europe. Quat. Int. 2013, 284, 7–14. [Google Scholar] [CrossRef]
- Petronio, C.; Pandolfi, L. Stephanorhinus hemitoechus (Falconer, 1868) del Pleistocene superiore dell’area di Melpignano-Cursi e S. Sidero (Lecce, Italia). Geol. Romana 2008, 41, 1–12. [Google Scholar]
- Ronchitelli, A.; Abbzazzi, L.; Accorsi, A.A.; Bandini Mazzanti, M.; Bernardi, M.; Masini, F.; Mercuri, A.; Mezzabotta, C.; Rook, L. The Grotta Grande of Scario (Salerno-southern Italy): Stratigraphy, archaeological finds, pollen and mammals. In Proceedings of the 1st International Congress on ‘Science and Technology for the Safeguard of Cultural Heritage in the Mediterranean Basin’, Siracusa, Italy, 27 November–2 December 1995. [Google Scholar]
- Mazza, B.; Rustioni, M.; Agostini, S.; Rossi, A. An unexpected Late Pleistocene macaque remain from Grotta degli Orsi Volanti (Rapino, Chieti, Central Italy). Geobios 2005, 3, 211–217. [Google Scholar] [CrossRef]
- van der Made, J.; Mazo, A.V. Los Grandes Mamíferos del yacimiento de PRERESA; Museo Arqueológico Regional, Alcalá de Henares: Comunidad de Madrid, Spain, 2014; pp. 40–67. [Google Scholar]
- Crégut-Bonnoure, E.; Fernandez, P. Perspectives morphométriques et phylogéniques du genre Capra au Pléistocène (Mammalia, Artiodactyla, Caprinae). Quaternaire 2018, 29, 243–254. [Google Scholar] [CrossRef]
- Lacombat, F. Biochronologie et grands mammifères au Pléistocène moyen et supérieur en Europe Occidentale: L’apport des Rhinocerotidae (Genre Stephanorhinus). Quaternaire 2009, 20, 429–435. [Google Scholar] [CrossRef]
- Breda, M.; Collinge, S.E.; Parfitt, S.A.; Lister, A.M. Metric analysis of ungulate mammals in the early Middle Pleistocene of Britain, in relation to taxonomy and biostratigraphy. I. Rhinocerotidae and Bovidae. Quat. Intern. 2010, 228, 136–156. [Google Scholar] [CrossRef]
- Guérin, C.; Baryshnikov, G.F. Le rhinocéros acheuléen de la grotte de Koudaro I (Géorgie, URSS) et le problème des espèces relictes du Pléistocène du Caucase. Geobios 1987, 20, 389–396. [Google Scholar] [CrossRef]
- Baryshnikov, G.F. Local biochronology of Middle and Late Pleistocene mammals from the Caucasus. J. Theriol. 2002, 1, 61–67. [Google Scholar] [CrossRef]
- Hoffecker, J.F.; Baryshnikov, G.F.; Doronichev, V.B. Large mammal taphonomy of the Middle Pleistocene hominid occupation at Treugol’naya Cave (Northern Caucasus). Quat. Sci. Rev. 2003, 22, 595–607. [Google Scholar] [CrossRef]
- Kahlke, H.D.; Kaiser, T. Generalism as a subsistence strategy; advantages and limitations of the higlhy flexible feeding traits of Pleistocene Stephanorhinus hudsheimensis (Rhinocerotidae, Mammalia). Quat. Sci. Rev. 2011, 30, 2250–2261. [Google Scholar] [CrossRef]
- Pandolfi, L.; Petronio, C. Stephanorhinus etruscus (Falconer, 1868) from Pirro Nord (Apricena, Foggia, Southern Italy) with notes on the older late Early Pleistocene rhinoceros remains of Italy. Riv. Ital. Paleontol. Stratigr. 2011, 117, 173–187. [Google Scholar]
- Anders, U.; von Koenigswald, W.; Ruf, I.; Smith, B.H. Generalized individual dental age stages for fossil and extant placental mammals. Paläontol. Z. 2011, 85, 321–339. [Google Scholar] [CrossRef]
- Hitchins, P.M. Age determination of the black rhinoceros (Diceros bicornis LINN.) in Zululand. S. Afr. J. Wildl. Res. 1978, 8, 71–80. [Google Scholar]
- Petronio, C. Una mandibola di rinoceronte di Ponte Galeria. Atti Soc. Sci. Nat. 1988, 129, 173–178. [Google Scholar]
- Bonifay, M.F. Dicerorhinus etruscus Falc. du Pléistocène moyen des grottes de Lunel-Viel (Hérault). Ann. Paléont. 1973, 59, 79–111. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Fernández, D.; Cerdeño, E.; Sanz, M.; Daura, J. The Latest Occurrence of Stephanorhinus hundsheimensis (Rhinocerotidae) in Europe: The Skeletons from the Cova del Rinoceront Site (Castelldefels, Barcelona). Quaternary 2023, 6, 60. https://doi.org/10.3390/quat6040060
García-Fernández D, Cerdeño E, Sanz M, Daura J. The Latest Occurrence of Stephanorhinus hundsheimensis (Rhinocerotidae) in Europe: The Skeletons from the Cova del Rinoceront Site (Castelldefels, Barcelona). Quaternary. 2023; 6(4):60. https://doi.org/10.3390/quat6040060
Chicago/Turabian StyleGarcía-Fernández, David, Esperanza Cerdeño, Montserrat Sanz, and Joan Daura. 2023. "The Latest Occurrence of Stephanorhinus hundsheimensis (Rhinocerotidae) in Europe: The Skeletons from the Cova del Rinoceront Site (Castelldefels, Barcelona)" Quaternary 6, no. 4: 60. https://doi.org/10.3390/quat6040060
APA StyleGarcía-Fernández, D., Cerdeño, E., Sanz, M., & Daura, J. (2023). The Latest Occurrence of Stephanorhinus hundsheimensis (Rhinocerotidae) in Europe: The Skeletons from the Cova del Rinoceront Site (Castelldefels, Barcelona). Quaternary, 6(4), 60. https://doi.org/10.3390/quat6040060