
An Updated Review of The Quaternary Hippopotamus Fossil Records from the Iberian Peninsula

 ,
,  ,
,  and
and 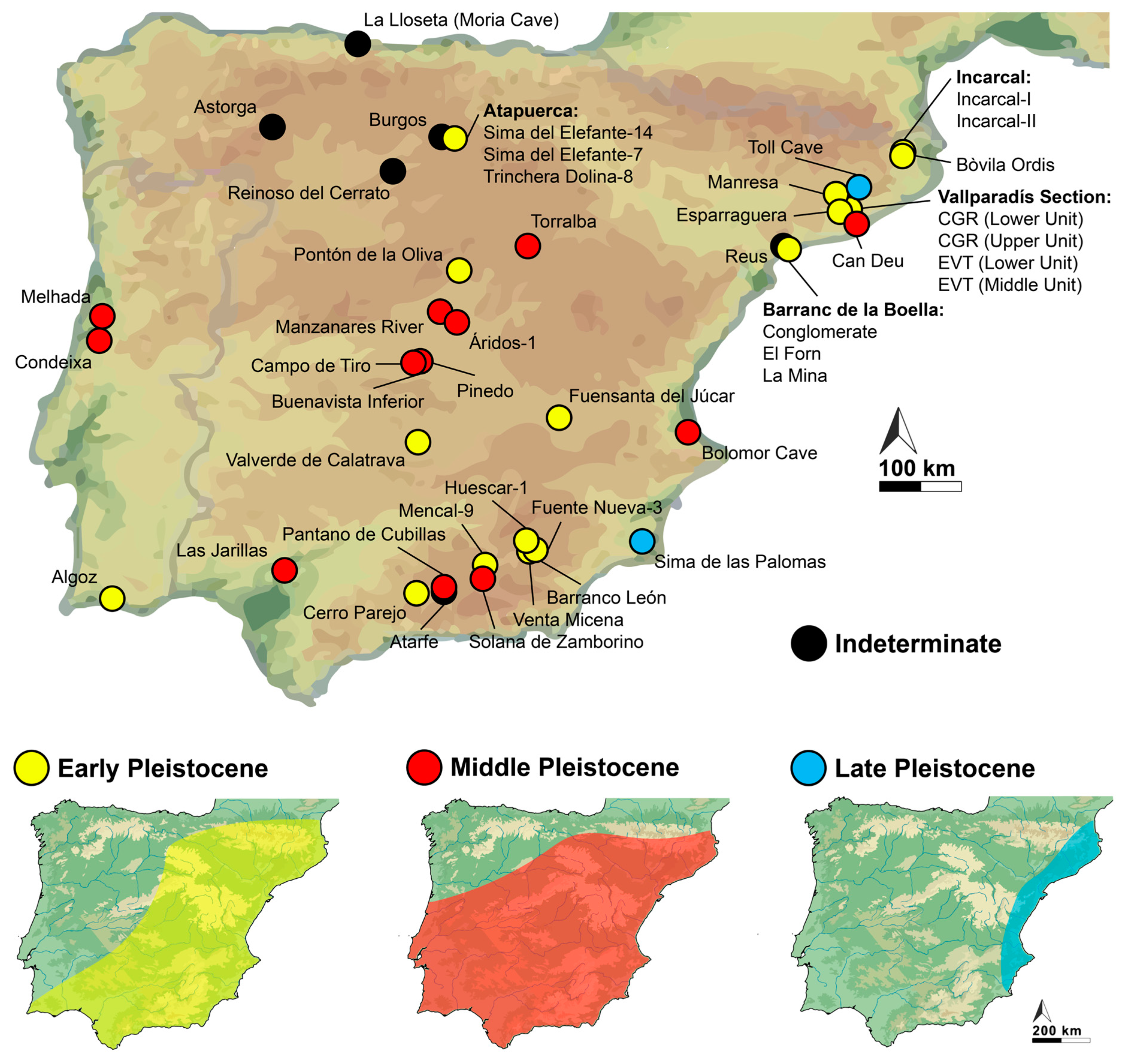{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
- Compilation and consultation of all of the available bibliographies on hippopotamus fossils on the Iberian Peninsula and the sites where they have been found. The key references for each case are listed in Table S1.
- Direct consultation with the researchers responsible for excavations on the findings. Data from this type of consultation are referenced as “pers. comm.”.
- Review of previously published historical collections and unpublished material deposited in numerous Iberian institutions. Museo Nacional de Ciencias Naturales (MNCN), Museo Geominero (IGME), Museo Arqueológico y Paleontológico de Madrid (MAR), Museo Arqueológico Nacional (MAN), Museo de San Isidro (MSI), Catedral de Astorga, Museo de la Rinconada, Institut Català de Paleoecologia Humana i Evolució Social (IPHES), Institut Català de Paleontologia (ICP), Museu Arqueològic Comarcal de Banyoles, Museu de Ciències Naturals de Barcelona, Museu del Seminari Conciliar de Barcelona, Museu d’Arqueologia de Catalunya, Museu d’Història de València, Museo Paleontològico de Elche, Museo de Arqueología de Múrcia, Museo Arqueológico de Cartagena, Museo Arqueológico y Etnológico de Granada, and Museu Geológico de Lisboa (MG).
- Collaboration in the evaluation of new finds. Barranc de la Boella, Vallparadís Estació, Cal Guardiola, Incarcal Complex (I, II and V), Las Jarillas, Bòbila Ordis, Barranco León D, Fuente Nueva-3, and Venta Micena.
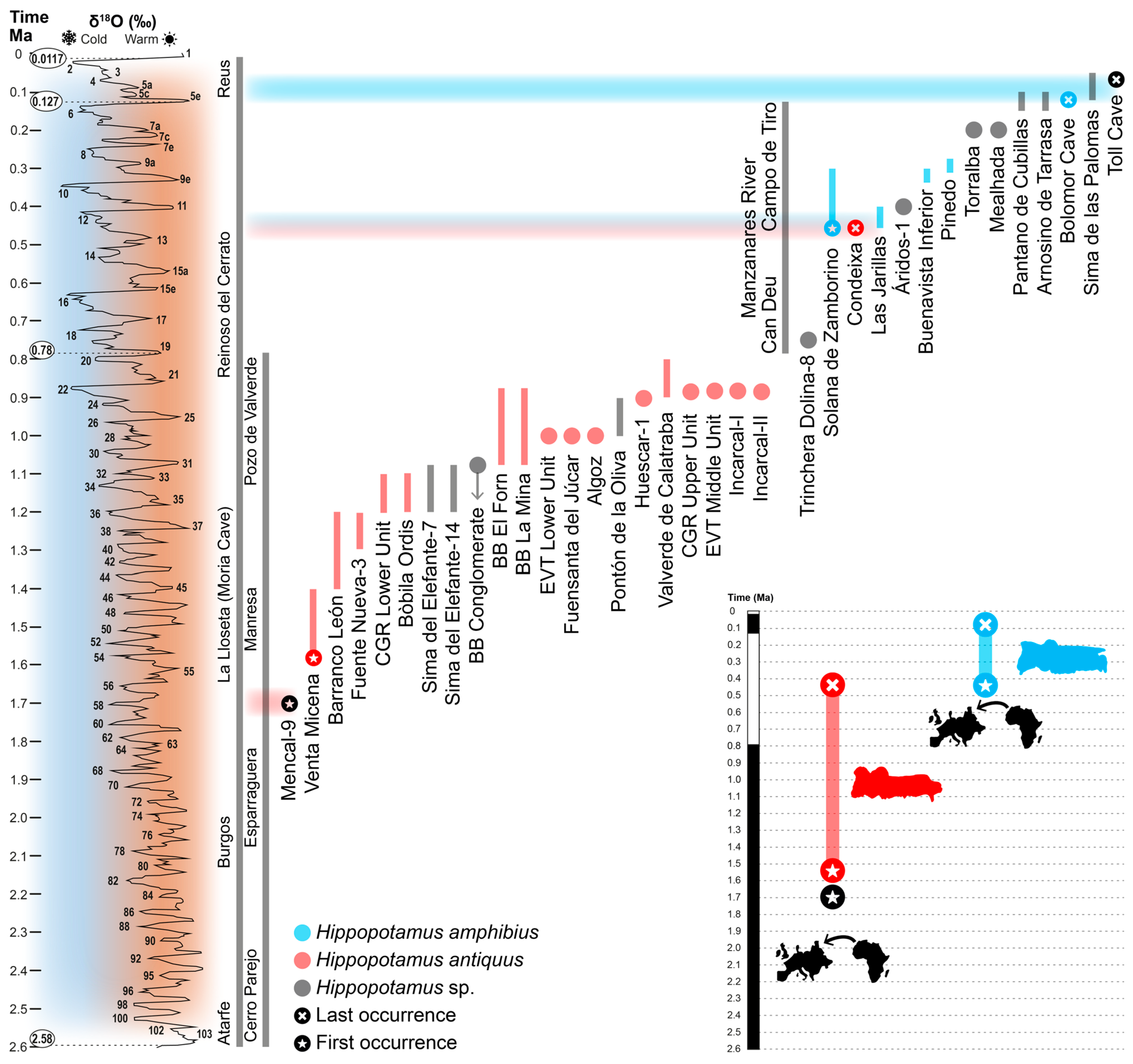
3. Results
4. Discussion
4.1. Phylogenetic and Biogeographical Relationships of Hippopotamus of the Iberian Peninsula
4.2. Paleoecological Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fidalgo, D.; Galli, E.; Madurell-Malapeira, J.; Rosas, A. Earliest Pleistocene European hippos: A review. Comun. Geol. 2021, 108, 69–73. [Google Scholar]
- Martino, R.; Pandolfi, L. The Quaternary Hippopotamus records from Italy. Hist. Biol. 2022, 34, 1146–1156. [Google Scholar] [CrossRef]
- Iannucci, A.; Mecozzi, B.; Sardella, R. Beware of the “Wolf event”–Remarks on large mammal dispersals in Europe and the late Villafranchian faunal turnover. Alp. Mediterr. Quat. 2023, 36, 75–90. [Google Scholar]
- Mazza, P. Interrelations between pleistocene hippopotami of Europe and Africa. Bollet. Soc. Paleont. Ital. 1991, 30, 153–186. [Google Scholar]
- Stuart, A.J.; Gibbard, P.L. Pleistocene occurrence of Hippopotamus in Britain. Quartärpaläontologie 1986, 6, 209–218. [Google Scholar]
- Pandolfi, L.; Petronio, C. A brief review of the occurrences of Pleistocene Hippopotamus (Mammalia, Hippopotamidae) in Italy. Geol. Croat. 2015, 68, 313–319. [Google Scholar] [CrossRef]
- Athanassiou, A. The fossil record of continental hippopotamids (Mammalia: Artiodactyla: Hippopotamidae) in Greece. In Fossil Vertebrates of Greece Vol. 2: Laurasiatherians, Artiodactyles, Perissodactyles, Carnivorans, and Island Endemics; Vlachos, E., Ed.; Springer: Cham Switzerland, 2022; pp. 281–299. [Google Scholar]
- Martino, R.; Ríos, M.I.; Mateus, O.; Pandolfi, L. Taxonomy, chronology, and dispersal patterns of Western European Quaternary hippopotamuses: New insight from Portuguese fossil material. Quat. Int. 2023, 674–675, 121–137. [Google Scholar] [CrossRef]
- Petronio, C. Note on the taxonomy of Pleistocene hippopotamuses. Ibex 1995, 3, 53–55. [Google Scholar]
- Fidalgo, D.; Rosas, A.; Madurell-Malapeira, J.; Pineda, A.; Huguet, R.; García-Tabernero, A.; Cáceres, I.; Ollé, A.; Vallverdú, J.; Saladie, P. A review on the Pleistocene occurrences and palaeobiology of Hippopotamus antiquus based on the record from the Barranc de la Boella Section (Francolí Basin, NE Iberia). Quat. Sci. Rev. 2023, 307, 108034. [Google Scholar] [CrossRef]
- Mazza, P. New evidence on the Pleistocene hippopotamuses of western Europe. Geol. Rom. 1995, 31, 61–241. [Google Scholar]
- Vekua, A.K. The lower Pleistocene mammalian fauna of Ahkalkalaki (southern Georgia, USSR). Palaeontogr. Ital. 1986, 74, 63–96. [Google Scholar]
- Mazza, P.; Bertini, A. Were Pleistocene hippopotamuses exposed to climate driven body size changes? Boreas 2013, 42, 194–209. [Google Scholar] [CrossRef]
- van der Made, J.; Sahnouni, M.; Boulaghraief, K. Hippopotamus gorgops from El Kherba (Algeria) and the Context of its Biogeography. In Proceedings of the II MAP, Burgos, Spain, 15–16 April 2015; pp. 135–169. [Google Scholar]
- Caloi, L.; Palombo, M.R.; Petronio, C. Resti cranici di Hippopotamus antiquus (= H. major) e Hippopotamus amphibius conservati nel Museo di Paleontologia dell’Università di Roma. Geol. Rom. 1980, 19, 91–119. [Google Scholar]
- Reynolds, S.H. A Monograph on the British Pleistocene Mammalia VOL. III, Part I. Hippopotamus. Pages 1–38; plates I–VI. Monogr. Palaeontogr. Soc. 1922, 74, 1–38. [Google Scholar] [CrossRef]
- Mecozzi, B.; Iannucci, A.; Mancini, M.; Tentori, D.; Cavasinni, C.; Conti, J.; Yuri Messina, M.; Sarra, A.; Sardella, R. Reinforcing the idea of an early dispersal of Hippopotamus amphibius in Europe: Restoration and multidisciplinary study of theskull from the Middle Pleistocene of Cava Montanari (Rome, central Italy). PLoS ONE 2023, 18, e0293405. [Google Scholar] [CrossRef] [PubMed]
- Bytinski-Salz, H. Recent findings of Hippopotamus in Israel. Isr. J. Zool. 1965, 14, 38–48. [Google Scholar]
- Silva, P.G.; Bardají, T.; Roquero García-Casal, E.; Baena Preysler, J.; Cearreta, A.; Rodríguez Pascua, M.A.; Rosas, A.; Zazo, C.; Goy, J.L. El periodo cuaternario: La historia geológica de la Prehistoria. Cuat. Geomorf. 2017, 31, 113–154. [Google Scholar]
- Espigares, M.P. Análisis y Modelización del Contexto Sedimentario y los Atributos Tafonómicos de los Yacimientos Pleistocénicos del Borde Nororiental de la Cuenca de Guadix-Baza. Ph.D. Thesis, Universidad de Granada, Granada, Spain, 2010. [Google Scholar]
- Madurell-Malapeira, J.; Alba, D.M.; Espigares, M.P.; Vinuesa, V.; Palmqvist, P.; Martínez-Navarro, B.; Moya-Sola, S. Were large carnivorans and great climatic shifts limiting factors for hominin dispersals? Evidence of the activity of Pachycrocuta brevirostris during the Mid-Pleistocene Revolution in the Vallparadís Section (Vallès-Penedès Basin, Iberian Peninsula). Quat. Int. 2017, 431, 42–52. [Google Scholar] [CrossRef]
- Blasco, R.; Peris, J.F. From tortoises to elephants: The impact of elephants in the broad spectrum diet at Bolomor Cave (MIS 9–5 Spain). In Contributions in Paleoanthropology Band 1: Human-Elephant Interactions: From Past to Present; Konidaris, G.E., Barkai, R., Tourloukis, V., Harvati, K., Eds.; Tuebingen Paleoanthropology Book Series: Tuebingen, Germany, 2021; pp. 363–385. [Google Scholar]
- Pla-Pueyo, S.; Viseras, C.; Soria, J.M.; Tent-Manclús, J.E.; Arribas, A. A stratigraphic framework for the pliocene-pleistocene continental sediments of the Guadix basin (betic Cordillera, S. Spain). Quat. Int. 2011, 243, 16–32. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Ruiz Bustos, A. Descripción y significado biostratigráfico y climático del Equus e Hippopotamus. en el yacimiento de Venta Micena (Granada). Estud. Geol. 1985, 41, 251–261. [Google Scholar] [CrossRef]
- Reimann, C.K.; Strauch, F. Ein Hippopotamus-Schädel aus dem Pliozän von Elis (Peloponnes, Griechenland). Neues Jahrbuch Geol. Palaontol.-Abhandlungen 2008, 249, 203–222. [Google Scholar] [CrossRef]
- Bellucci, L.; Bona, F.; Corrado, P.; Magri, D.; Mazzini, I.; Parenti, F.; Scardia, G.; Sardella, R. Evidence of late Gelasian dispersal of African fauna at Coste San Giacomo (Anagni Basin, central Italy): Early Pleistocene environments and the background of early human occupationin Europe. Quat. Sci. Rev. 2014, 96, 72–85. [Google Scholar] [CrossRef]
- Mazza, P.; Rustioni, M. The fossil bear from Senèze (Southern France). Rend. Lincei. 1994, 5, 17–26. [Google Scholar] [CrossRef]
- Martínez-Navarro, B.; Palmqvist, P.; Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Torregrosa, V.; Pérez-Claros, J.A. La fauna de grandes mamíferos de Fuente Nueva-3 y Barranco León-5: Estado de la cuestión. In Ocupaciones Humanas en el Pleistoceno Inferior y Medio de la Cuenca de Guadix-Baza; Toro, I., Martínez-Navarro, B., Agustí, J., Eds.; Junta de Andalucía, Consejería de Cultura: Sevilla, Spain, 2010; pp. 197–236. [Google Scholar]
- Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Alba, D.M.; Aurell-Garrido, J. Villafranchian large mammals from the Iberian Peninsula: Paleobiogeography, paleoecology and dispersal events. J. Iber. Geol. 2014, 40, 167–178. [Google Scholar] [CrossRef]
- Antunes, M.T.; Azzaroli, A.; Faure, M.; Guérin, C.; Mein, P. Mammifères pleistocènes de Algoz, en Algarve: Une révision. Ciênc. Terra. 1986, 8, 73–86. [Google Scholar]
- Galobart, À.; Ros, X.; Maroto, J.; Vila, B. Descripción del material de hipopótamo (Hippopotamus antiquus Desmarest, 1822) de los yacimientos del Pleistoceno inferior de Incarcal (Girona, NE de la Península Ibérica). Paleont. Evol. 2003, 34, 153–173. [Google Scholar]
- Agustí, J.; Leroy, S.A.; Lozano-Fernández, I.; Julià, R. Joint vegetation and mammalian records at the early Pleistocene sequence of Bòbila Ordis (Banyoles-Besalú Basin, NE Spain) and their bearing on early hominin occupation in Europe. Paleobiodivers. Paleoenviron. 2018, 98, 653–662. [Google Scholar] [CrossRef]
- Mazza, P.P.A.; Ventra, D. Pleistocene debris-flow deposition of the hippopotamus-bearing Collecurti bonebed (Macerata, Central Italy): Taphonomic and paleoenvironmental analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 296–314. [Google Scholar] [CrossRef]
- van der Made, J.; Rosell, J.; Blasco, R. Faunas from Atapuerca at the Early- Middle Pleistocene limit: The ungulates from level TD8 in the context of climatic change. Quat. Int. 2017, 433, 296–346. [Google Scholar] [CrossRef]
- Álvarez-Posada, C.; Parés, J.M.; Sala, R.; Viseras, C.; Pla-Pueyo, S. New magnetostratigraphic evidence for the age of Acheulean tools at the archaeo-palaeontological site “Solana del Zamborino” (Guadix–Baza Basin, S Spain). Sci. Rep. 2017, 7, 13495. [Google Scholar] [CrossRef]
- Baena, R.; Cunha, P.P.; Guerrero, I.C.; Fernández, J.J.; Cortés, M.; Silva, P.G.; Simón, M.; Murray, A.S.; Buylaert, J.P. El pleistoceno medio y superior en la secuencia ceneral de las terrazas del Guadalquivir (sgtg): Nuevas dataciones por luminiscencia de la t12 y t13 en la Rinconada (SW de España). In Proceedings of the XV Reunión Nacional Cuaternario, Bilbao, Spain, 1–5 July 2019; pp. 369–372. [Google Scholar]
- Gómez-Alba, J. Catálogo razonado de los vertebrados fósiles de España del Museo de Geología de Barcelona (1882–1982). In Treballs del Museu de Geologia de Barcelona; Ajuntament de Barcelona: Museu de Geologia: Barcelona, Spain, 1997; pp. 1–296. [Google Scholar]
- Blain, H.A.; Santonja, M.; Pérez-González, A.; Panera, J.; Rubio-Jara, S. Climate and environments during Marine Isotope Stage 11 in the central Iberian Peninsula: The herpetofaunal assemblage from the Acheulean site of Áridos-1, Madrid. Quat. Sci. Rev. 2014, 94, 7–21. [Google Scholar] [CrossRef]
- Sese, C.; Soto, E. Vertebrados del Pleistoceno de Madrid. Flora Fauna 2000, 6, 216–243. [Google Scholar]
- Blasco, R.; Rosell, J.; Peris, J.F.; Cáceres, I.; Vergès, J.M. A new element of trampling: An experimental application on the Level XII faunal record of Bolomor Cave (Valencia, Spain). J. Archaeol. Sci. 2008, 35, 1605–1618. [Google Scholar] [CrossRef]
- Faure, M. Hippopotamus incognitus nov. sp., un hippopotame (Mammalia, Artiodactyla) du Pléistocène d’Europe occidentale. Geobios 1984, 17, 427–437. [Google Scholar] [CrossRef]
- Donner, J.J.; Kurtén, B. The floral and faunal succession of “Cueva del Toll”, Spain. E G Quat. Sci. J. 1958, 9, 72–82. [Google Scholar] [CrossRef]
- Trinkaus, E.; Walker, M. The People of Palomas: Neandertals from the Sima de las Palomas del Cabezo Gordo, Southeastern Spain; Texas A&M University Press: College Station, TX, USA, 2017; p. 320. [Google Scholar]
- Pieruccini, P.; Forti, L.; Mecozzi, B.; Iannucci, A.; Yu, T.L.; Shen, C.C.; Bona, F.; Lembo, G.; Muttillo, B.; Sardella, R.; et al. Stratigraphic reassessment of Grotta Romanelli sheds light on Middle-Late Pleistocene palaeoenvironments and human settling in the Mediterranean. Sci. Rep. 2022, 12, 13530. [Google Scholar] [CrossRef]
- Palmqvist, P.; Rodríguez-Gomez, G.; Figueirido, B.; García-Aguilar, J.M.; Perez-Claros, J.A. On the ecological scenario of the first hominin dispersal out of Africa. Anthropologie 2022, 126, 102998. [Google Scholar] [CrossRef]
- Kierdorf, U.; Kahlke, R.D. Pathological findings on remains of hippopotamids from the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld near Meiningen (Thüringen, Germany). Part 4; Kahlke, R.D., Ed.; Senckenberg: Frankfurt, Germany, 2020; Volume 40, pp. 1251–1272. [Google Scholar]
- Adams, N.F.; Candy, I.; Schreve, D.C. An Early Pleistocene hippopotamus from Westbury Cave, Somerset, England: Support for a previously unrecognized temperate interval in the British Quaternary record. J. Quat. Sci. 2022, 37, 28–41. [Google Scholar] [CrossRef]
- Head, M.J.; Gibbard, P.L. Early–Middle Pleistocene transitions: Linking terrestrial and marine realms. Quat. Int. 2015, 389, 7–46. [Google Scholar] [CrossRef]
- Fidalgo, D.; Rosas, A.; Bartolini-Lucenti, S.; Boisserie, J.-R.; Pandolfi, L.; Martínez-Navarro, B.; Palmqvist, P.; Rook, L.; Madurell Malapeira, J. Increase on environmental seasonality through the European Early Pleistocene inferred from dental enamel hypoplasia. Sci. Rep. 2023, 13, 16941. [Google Scholar] [CrossRef]
- Berends, C.J.; Köhler, P.; Lourens, L.J.; Van de Wal, R.S.W. On the cause of the Mid-Pleistocene transition. Rev. Geophys. 2021, 59, e2020RG000727. [Google Scholar] [CrossRef]
- Fidalgo, D.; Rosas, A.; Estalrrich, A.; García-Tabernero, A.; Pastor, F.J.; Pineda, A.; Huguet, R.; Cáceres, I.; Ollé, A.; Vallverdú, J.; et al. First presence of Macaca sylvanus at the late early Pleistocene of barranc de la Boella (La Mina locality, Francolí basin, NE Iberia). J. Hum. Evol. 2023, 179, 103368. [Google Scholar] [CrossRef] [PubMed]
- Boscaini, A.; Madurell-Malapeira, J.; Llenas, M.; Martínez-Navarro, B. The origin of the critically endangered Iberian lynx: Speciation, diet and adaptive changes. Quat. Sci. Rev. 2015, 123, 247–253. [Google Scholar] [CrossRef]
- Duval, M.; Martínez-Navarro, B.; Oms, O.; Rufí, I.; Sorbelli, I.; Prat-Vericat, M.; Madurell-Malapeira, J. The Early Pleistocene fossil assemblage from Incarcal-I: First numerical dating results. In Proceedings of the XXI Inqua Congress, Rome, Italy, 13–20 July 2023. [Google Scholar]
- Madurell-Malapeira, J. Estudi de les Restes d’Hippopotamus antiquus (Hippopotamidae, Mammalia) del Jaciment del Pleistocè Inferior de Cal Guardiola (Terrassa). Master’s Thesis, Universidad Autònoma de Barcelona, Barcelona, Spain, 2006. [Google Scholar]
- Madurell-Malapeira, J.; Minwer-Barakat, R.; Alba, D.M.; Garcés, M.; Gómez, M.; Aurell-Garrido, J.; Ros-Montoya, S.; Moyà-Solà, S.; Berástegui, X. The Vallparadís section (Terrassa, Iberian Peninsula) and the latest Villafranchian faunas of Europe. Quat. Sci. Rev. 2010, 29, 3972–3982. [Google Scholar] [CrossRef]
- Espigares, M.P.; Palmqvist, P.; Rodríguez-Ruiz, M.D.; Ros-Montoya, S.; Pérez-Ramos, A.; Rodríguez-Gómez, G.; Guerra-Merchán, A.; García-Aguilar, J.M.; Granados, A.; Campaña, I.; et al. Sharing food with hyenas: A latrine of Pachycrocuta brevirostris in the Early Pleistocene assemblage of Fuente Nueva-3 (Orce, Baza Basin, SE Spain). Archaeol. Anthropol. Sci. 2023, 15, 1–29. [Google Scholar] [CrossRef]
- Pineda, A.; Saladié, P.; Huguet, R.; Cáceres, I.; Rosas, A.; García-Tabernero, A.; Estalrrich, A.; Mosquera, M.; Ollé, A.; Vallverdú, J. Coexistence among large predators during the Lower Paleolithic at the site of La Mina (Barranc de la Boella, Tarragona, Spain). Quat. Int. 2015, 388, 177–187. [Google Scholar] [CrossRef]
- Konidaris, G.; Tourloukis, V.; Boni, G.; Athanassiou, A.; Giusti, D.; Thompson, N.; Syrides, G.; Panagopoulou, E.; Karkanas, P.; Harvati, K. Marathousa 2: A New Middle Pleistocene Locality in the Megalopolis Basin (Greece) with Evidence of Hominin Exploitation of Megafauna (Hippopotamus). PaleoAnthropology 2023, 2023, 34–55. [Google Scholar]
- Yravedra, J.; Solano, J.A.; Herranz-Rodrigo, D.; Linares-Matás, G.J.; Saarinen, J.; Rodríguez-Alba, J.J.; Titton, S.; Serrano-Ramos, A.; Courtenay, L.A.; Mielgo, C.; et al. Unravelling hominin activities in the zooarchaeological assemblage of Barranco León (Orce, Granada, Spain). J. Paleolit. Archaeol. 2022, 5, 6. [Google Scholar] [CrossRef]
- Alonso Herrero, E.; Fernández-Martínez, E.; Rodríguez-Fernández, L.R.; Matías Rodríguez, R. Guía Geológica Visual de León; Editorial Celarayn: León, Spain, 2004; p. 274. [Google Scholar]
- Rosas, A.; Huguet, R.; Pérez-González, A.; Carbonell, E.; Bermúdez de Castro, J.M.; Vallverdú, J.; van der Made, J.; Allue, E.; Garcia, N.; Martinez-Pérez, R.; et al. The “Sima del Elefante” cave site at Atapuerca (Spain). Estud. Geol. 2006, 62, 327–348. [Google Scholar] [CrossRef]
- Rodríguez, J.; Burjachs, F.; Cuenca-Bescós, G.; García, N.; van der Made, J.; Pérez González, A.P.; Blain, H.-A.; Expósito, I.; López-García, J.M.; García Antón, M.; et al. One million years of cultural evolution in a stable environment at Atapuerca (Burgos, Spain). Quat. Sci. Rev. 2011, 30, 1396–1412. [Google Scholar] [CrossRef]
- Van der Made, J. Ungulados de Gran Dolina, Atapuerca: Nuevos datos e interpretaciones. Atapuerca Evol. Hum. 1998, 97–110. [Google Scholar]
- Aguirre, E. Remarques sur la stratigraphie et la paléontologie du bassin de Granada (Espagne). Comptes Rendus Hebd. Seances L Acad. Des Sci. 1958, 246, 2140–2142. [Google Scholar]
- Sánchez, B.; Pesquero, M.D.; Fraile, S.; Salesa, M.J. Las colecciones de Vertebrados fósiles del Museo Nacional de Ciencias Naturales (CSIC): Aportación del Profesor Emiliano Aguirre a la Paleontología española. Zona Arqueol. 2004, 4, 527. [Google Scholar]
- Crusafont, M. El cuaternario español y su fauna de mamíferos: Ensayo de síntesis. Rev. Fac. Cienc. 1960, 2, 181–199. [Google Scholar]
- Bech, J. Nuevo hallazgo de Hippopotamus ampphibius major en Banyoles (Gerona). Acta Geológica Hispánica 1970, 51–53. [Google Scholar]
- Julià, R. Nuevos datos sobre la posición cronoestratigráfica de los materiales cuaternarios de la cuenca lacustre de Banyoles-Besalú (Girona). Acta Geol. Hisp. 1977, 12, 55–59. [Google Scholar]
- Faure, M. Les hippopotamidés du Pléistocène ancien d’Oubeidiyeh (Israèl). Mem. Travaux Cent. Rech. Fr. Jerus. 1986, 5, 107–142. [Google Scholar]
- Ruiz-Bustos, A.; Fernández, J.; Morales, J.; Rodríguez-Fernández, J.; Vera, J.A. Bioestratigrafia de los materiales Plio-Cuaternarios del borde norte de la Depresion de Granada. Estud. Geol. 1990, 46, 277–290. [Google Scholar] [CrossRef]
- Cardoso, J.L. Contribuição Para o Conhecimento dos Grandes Mamíferos do Plistocénico Superior de Portugal; OEIRAS: Oeiras, Portugal, 1993; pp. 171–567. [Google Scholar]
- Estraviz, Q. Quaternary Fossil Vertebrates from Continental Portugal: Paleobiodiversity, Revision of Specimens and New Localities. Master Thesis, Universidade Nova de Lisboa, Lisboa, Portugal, May 2019. [Google Scholar]
- Mazo, A.V.; Pérez-González, A.; Aguirre, E. Las faunas pleistocenas de Fuensanta del Jucar y El Provencio y su significado en la evolucion del Cuaternario de la Llanura manchega. Bol. Geol. Min. 1990, 101, 404–418. [Google Scholar]
- Mazo, A.V. Los hipopótamos del Pleistoceno medio de Huescar-1 (Granada). Trab. Neog. Cuantern. 1989, 1, 317–325. [Google Scholar]
- Alberdi, M.T.; Alcalá, L.; Azanza, B.; Cerdeño, E.; Mazo, A.V.; Morales, J.; Sesé, C. Consideraciones Bioestratigráficas Sobre la Fauna de Vertebrados Fósiles de la Cuenca de Guadix-Baza (Granada, España); CSIC—Museo Nacional de Ciencias Naturales (MNCN): Madrid, Spain, 1989; Volume 11, pp. 347–355. [Google Scholar]
- Domingo, M.S.; Alberdi, M.T.; Sánchez Chillón, B.; Cerdeño, E. La fauna cuaternaria de la Cornisa Cantábrica en las colecciones del Museo Nacional de Ciencias Naturales. MUNIBE 2005, 57, 5–30. [Google Scholar]
- Mariezkurrena-Gastearena, K. Hallazgos de macromamíferos poco frecuentes en yacimientos arqueológicos y paleontológicos del Pleistoceno de la región cantábrica. Kobie Ser. Paleoanthropol. 2011, 30, 83–110. [Google Scholar]
- Baena Escudero, R.; Fernández Caro, J.J.; Guerrero Amador, I.; Posada Simeón, J.C. La Terraza compleja del río Gualdaquivir en “Las Jarillas” (La Rinconada, Sevilla. SW de España): Cronoestratiagrafía, industria lítica y macro-fauna asociada. Cuat. Geomorf. 2014, 28, 107–125. [Google Scholar]
- Soto, E. Estudio paleontologico. In El yacimiento Achelense de Pinedo (Toledo); Querol, M.A., Santonja, M., Eds.; Ministerio de Cultura: Madrid, Spain, 1979; pp. 39–42. [Google Scholar]
- López-Recio, M.; Silva, P.G.; Roquero, E.; Cunha, P.P.; Tapias, F.; Alcaraz-Castaño, M.; Baena, J.; Cuartero, F.; Morín, J.; Torres, T.; et al. Geocronología de los yacimientos achelenses de Pinedo y Cien Fanegas (Valle del Tajo) e implicaciones en la evolución fluvial en el entorno de Toledo (España). Estud. Geol. 2015, 71, e029. [Google Scholar] [CrossRef]
- Nolte, E. Miscelánea Arqueológica. Kobie 1977, 7, 9–27. [Google Scholar]
- Vilaseca, S. Reus y su Entorno en la Prehistoria; Asociación de Estudios Reusenses: Reus, Spain, 1973; pp. 1–282. [Google Scholar]
- Jiménez-Arenas, J.M.; Santonja, M.; Botella, M.; Palmqvist, P. The oldest handaxes in Europe: Fact or artefact? J. Archaeol. Sci. 2011, 38, 3340–3349. [Google Scholar] [CrossRef]
- Pineda, A.; Saladié, P. The Middle Pleistocene site of Torralba (Soria, Spain): A taphonomic view of the Marquis of Cerralbo and Howell faunal collections. Archaeol. Anthropol. Sci. 2019, 11, 2539–2556. [Google Scholar] [CrossRef]
- Bonadonna, F.P.; Alberdi, M.T. Equus stenonis Cocchi as a biostratigraphical marker in the Neogene-Quaternary of the Western Mediterranean basin: Consequence on Galerian-Villafranchian chronostratigraphy. Quat. Sci. Rev. 1987, 6, 55–66. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fidalgo, D.; Madurell-Malapeira, J.; Martino, R.; Pandolfi, L.; Rosas, A. An Updated Review of The Quaternary Hippopotamus Fossil Records from the Iberian Peninsula. Quaternary 2024, 7, 4. https://doi.org/10.3390/quat7010004
Fidalgo D, Madurell-Malapeira J, Martino R, Pandolfi L, Rosas A. An Updated Review of The Quaternary Hippopotamus Fossil Records from the Iberian Peninsula. Quaternary. 2024; 7(1):4. https://doi.org/10.3390/quat7010004
Chicago/Turabian StyleFidalgo, Darío, Joan Madurell-Malapeira, Roberta Martino, Luca Pandolfi, and Antonio Rosas. 2024. "An Updated Review of The Quaternary Hippopotamus Fossil Records from the Iberian Peninsula" Quaternary 7, no. 1: 4. https://doi.org/10.3390/quat7010004
APA StyleFidalgo, D., Madurell-Malapeira, J., Martino, R., Pandolfi, L., & Rosas, A. (2024). An Updated Review of The Quaternary Hippopotamus Fossil Records from the Iberian Peninsula. Quaternary, 7(1), 4. https://doi.org/10.3390/quat7010004