
The Occurrence of Suids in the Post-Olduvai to Pre-Jaramillo Pleistocene of Europe and Implications for Late Villafranchian Biochronology and Faunal Dynamics


Abstract
:1. Introduction
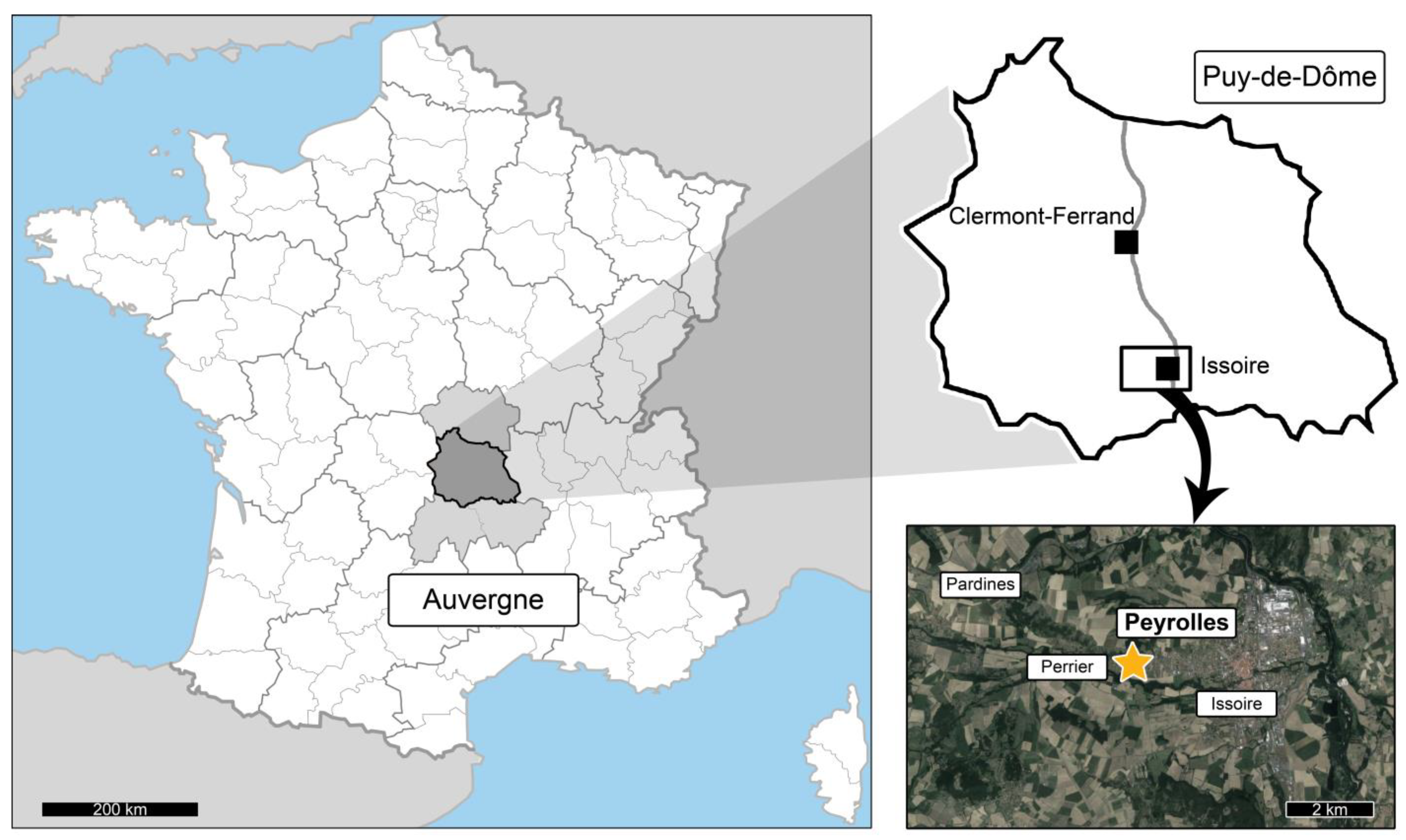
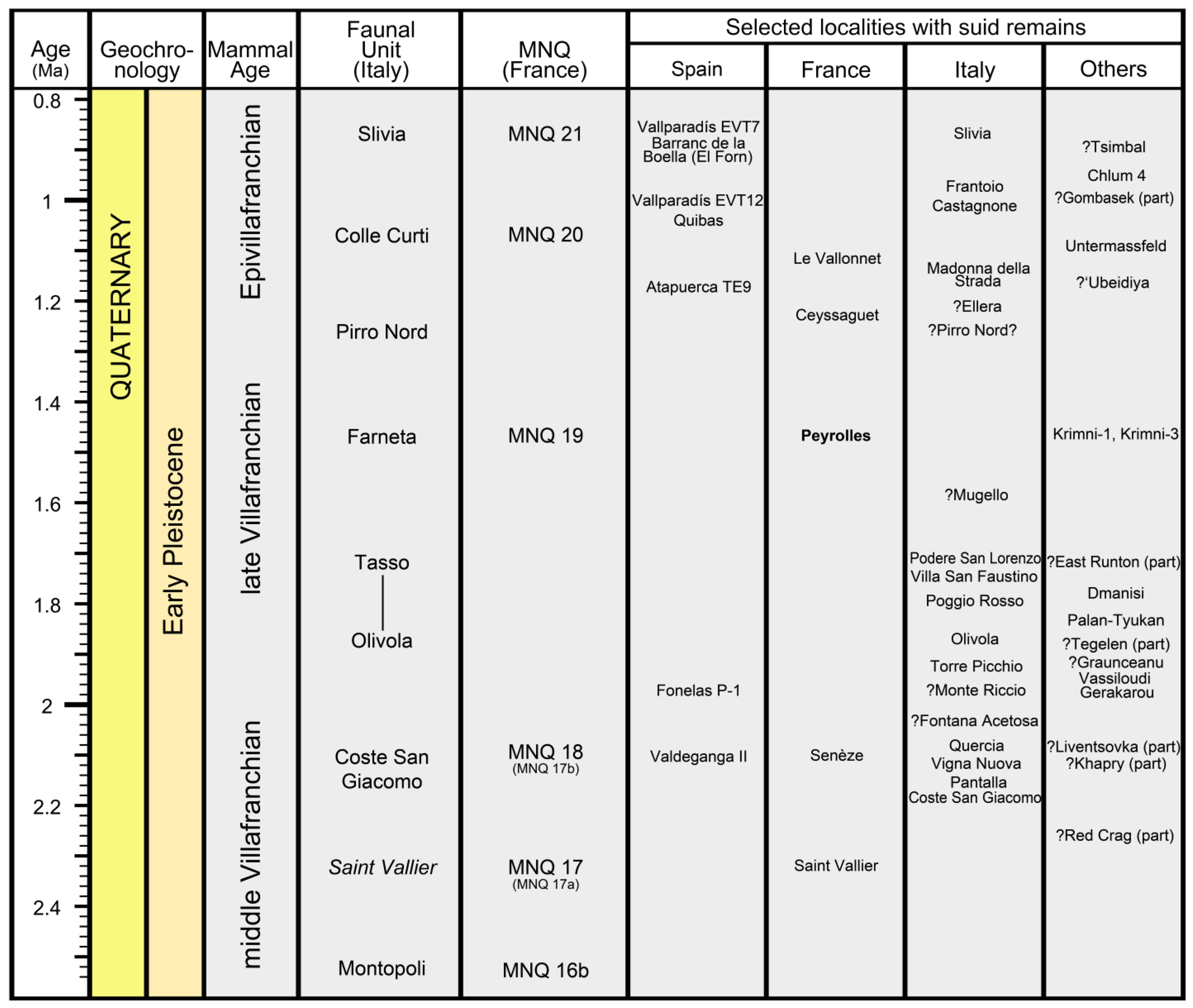
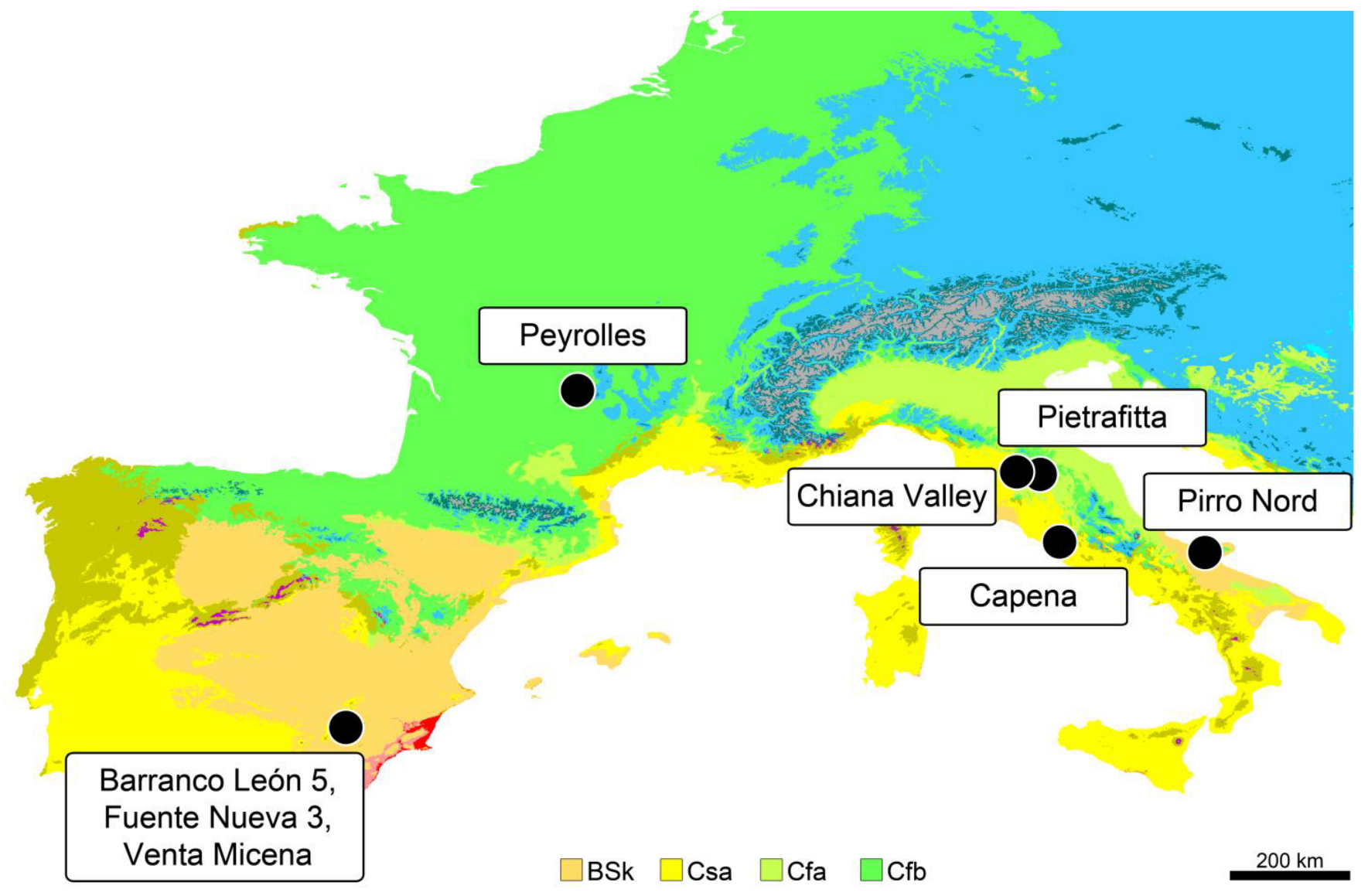
2. Peyrolles
3. Materials and Methods
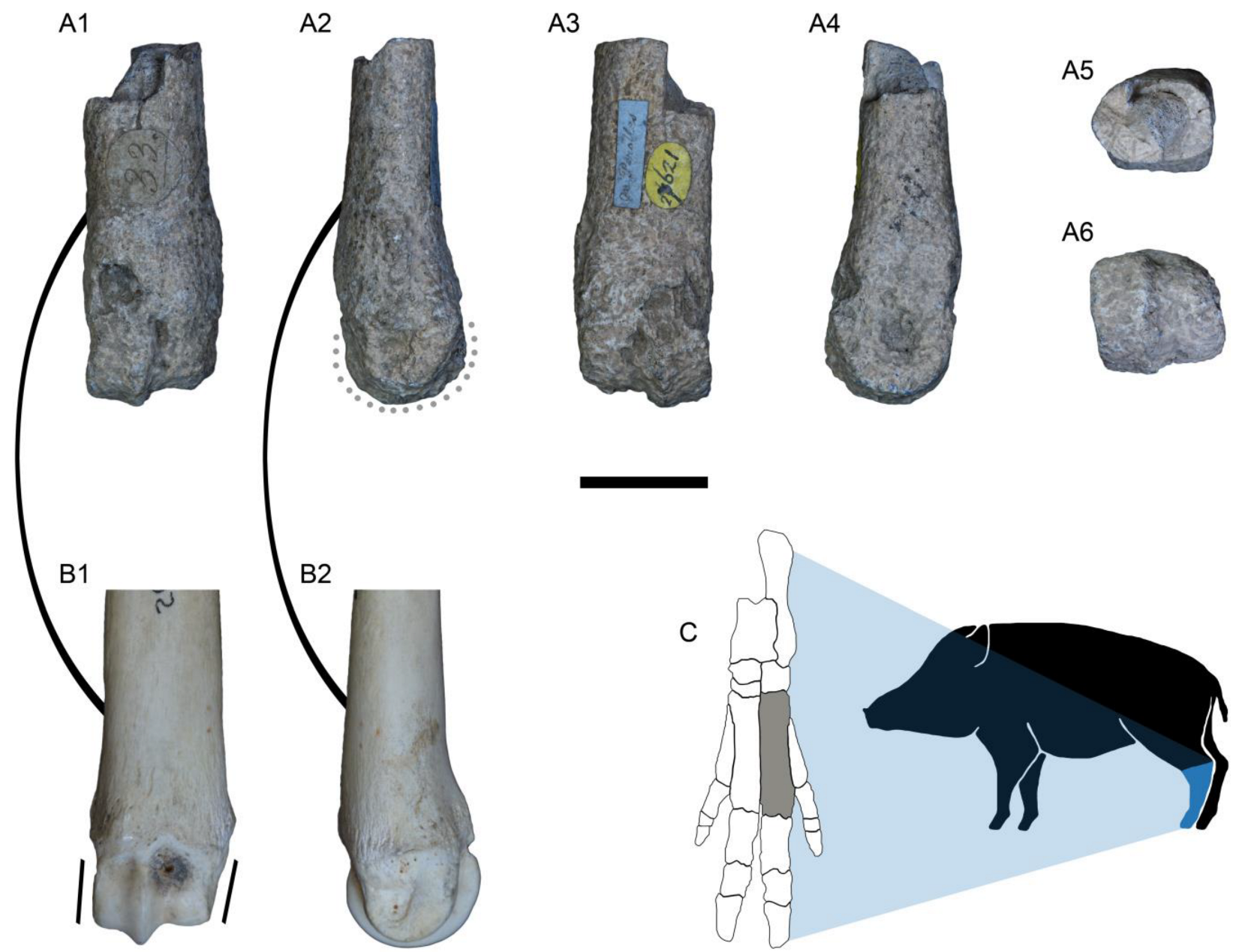
3.1. Fossil Material
3.2. Reproductive Strategies
3.3. Abundance and Frequency of Suids in the Fossil Record
4. Systematic Paleontology
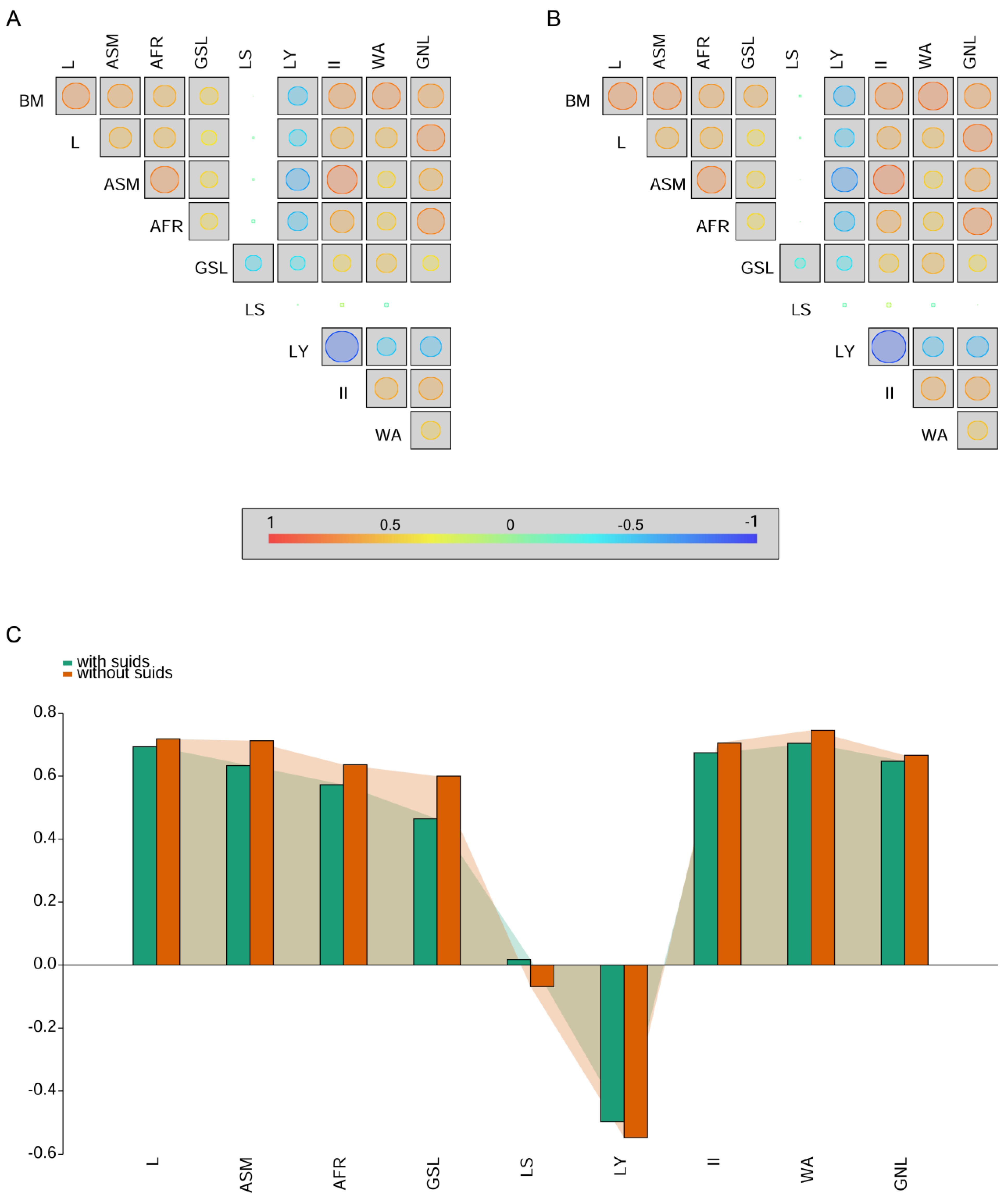
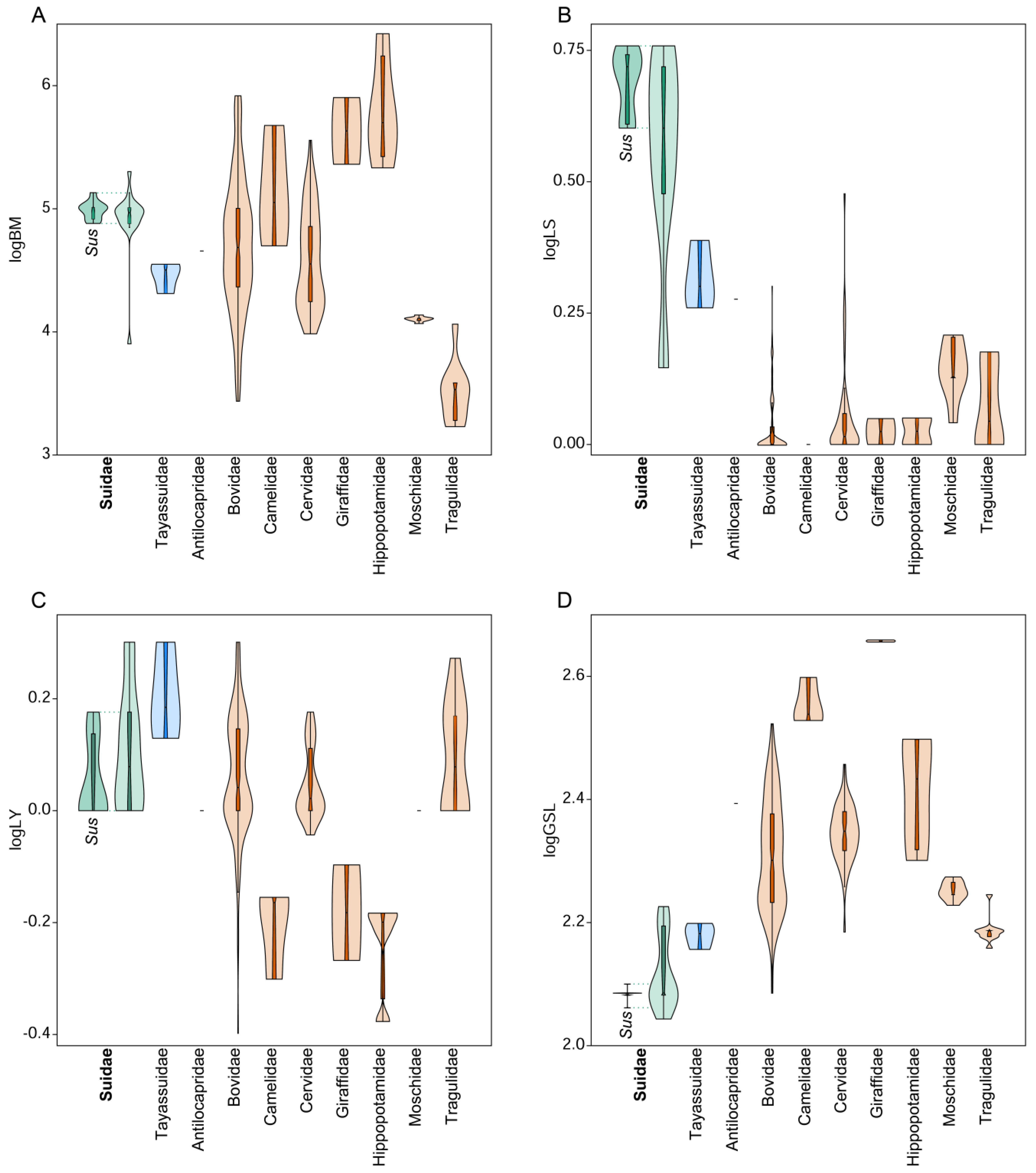
5. Reproductive Strategies
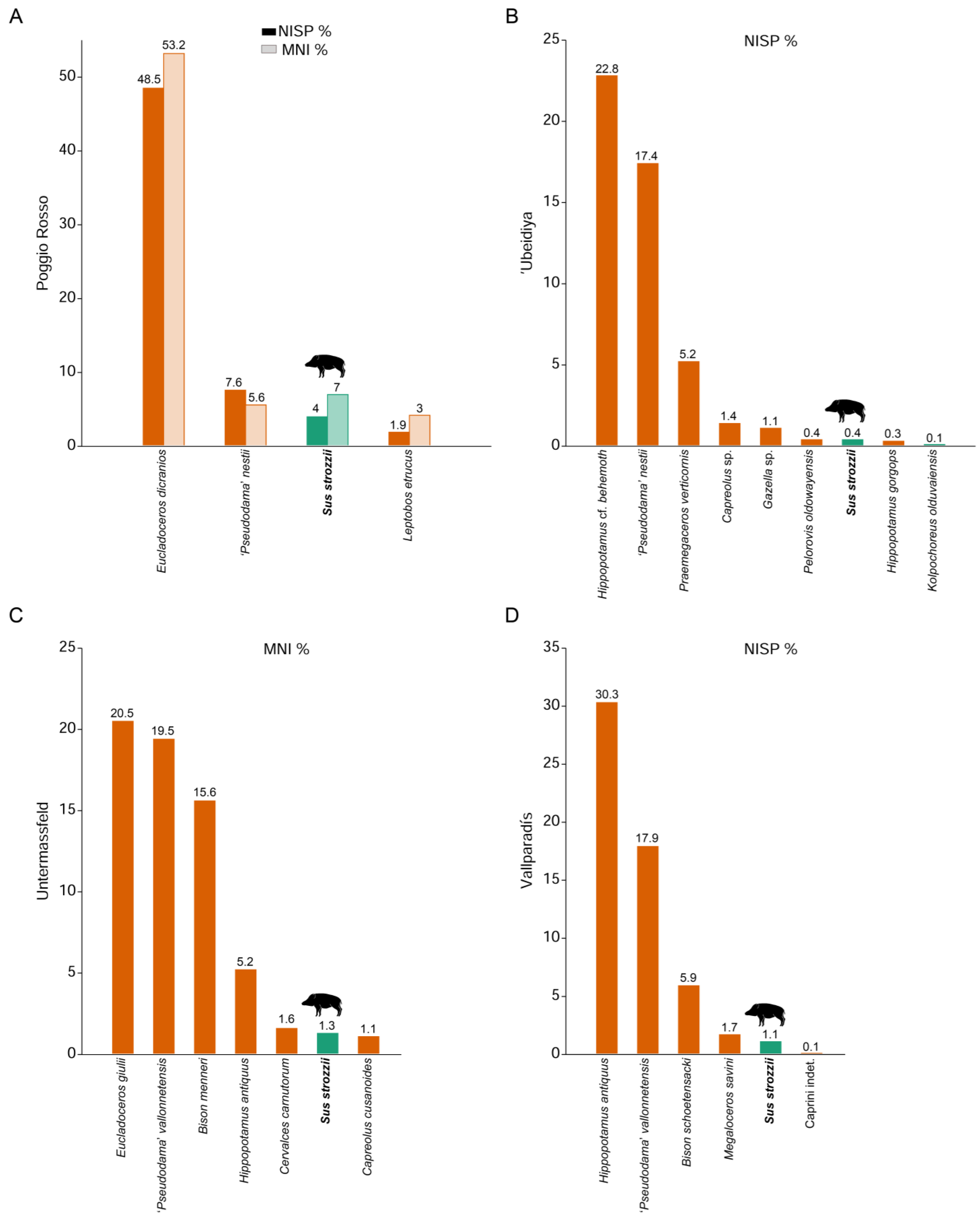
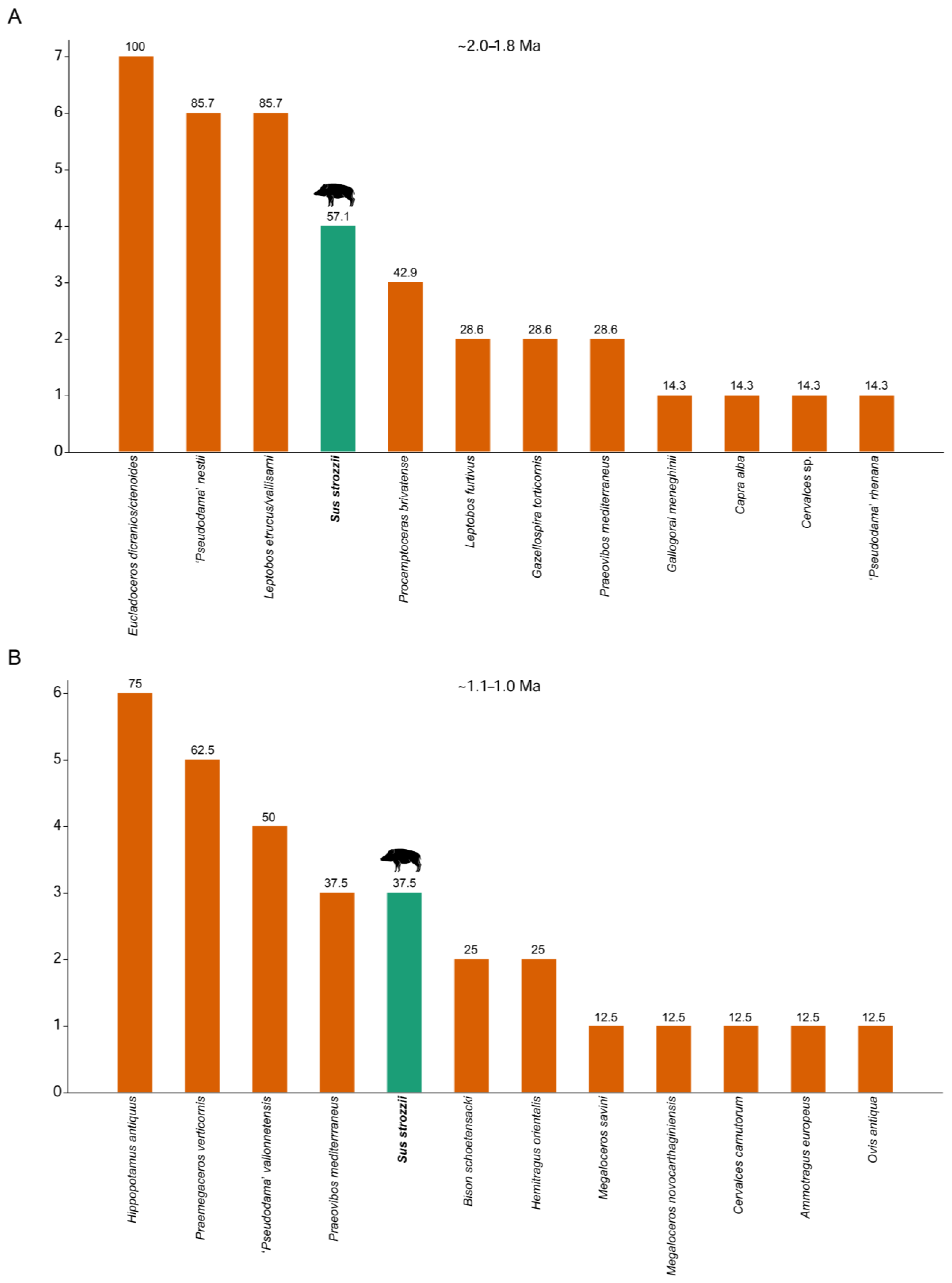
6. Abundance and Frequency in the Fossil Record
7. Discussion
7.1. The Reproductive Strategy of Suids
7.2. Abundance and Frequency of Suids in the Early Pleistocene of Europe
7.3. Fossil Localities and the “Suid Gap”
7.4. Implications for Late Villafranchian Biochronology and Faunal Dynamics
8. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shackleton, N.J. New data on the evolution of Pliocene climatic variability. In Paleoclimate and Evolution, with Emphasis on Human Evolution; Vrba, E.S., Denton, G.H., Patridge, T.C., Burckle, L.H., Eds.; Yale University Press: London, UK, 1995; pp. 242–248. [Google Scholar]
- Kahlke, R.-D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.A.; Spassov, N.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Palombo, M.R. Deconstructing mammal dispersals and faunal dynamics in SW Europe during the Quaternary. Quat. Sci. Rev. 2014, 96, 50–71. [Google Scholar] [CrossRef]
- Gibbard, P.L.; Head, M.J. The Quaternary Period. In Geologic Time Scale 2020; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1217–1255. [Google Scholar]
- Iannucci, A.; Sardella, R. What does the “Elephant-Equus” event mean today? Reflections on mammal dispersal events around the Pliocene-Pleistocene boundary and the flexible ambiguity of biochronology. Quaternary 2023, 6, 16. [Google Scholar] [CrossRef]
- Rightmire, G.P.; Margvelashvili, A.; Lordkipanidze, D. Variation among the Dmanisi hominins: Multiple taxa or one species? Am. J. Phy. Anthrop. 2019, 168, 481–495. [Google Scholar] [CrossRef] [PubMed]
- Tappen, M.; Bukhsianidze, M.; Ferring, R.; Coil, R.; Lordkipanidze, D. Life and death at Dmanisi, Georgia: Taphonomic signals from the fossil mammals. J. Hum. Evol. 2022, 171, 103249. [Google Scholar] [CrossRef]
- Muttoni, G.; Scardia, G.; Kent, D.V. Early hominins in Europe: The Galerian migration hypothesis. Quat. Sci. Rev. 2018, 180, 1–29. [Google Scholar] [CrossRef]
- Falguères, C. The first human settlements out africa into Europe: A chronological perspective. Quat. Sci. Rev. 2020, 247, 106551. [Google Scholar] [CrossRef]
- Azzaroli, A. Quaternary mammals and the “end-Villafranchian” dispersal event—A turning point in the history of Eurasia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1983, 44, 117–139. [Google Scholar] [CrossRef]
- Azzaroli, A.; De Giuli, C.; Ficcarelli, G.; Torre, D. Late Pliocene to early mid-Pleistocene mammals in Eurasia: Faunal succession and dispersal events. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 66, 77–100. [Google Scholar] [CrossRef]
- Iannucci, A.; Mecozzi, B.; Sardella, R. Beware of the “Wolf event”–Remarks on large mammal dispersals in Europe and the late Villafranchian faunal turnover. Alp. Mediterr. Quat. 2023, 36, 75–90. [Google Scholar] [CrossRef]
- Iannucci, A. Sus strozzii (Suidae, Mammalia) from the historical locality of Quercia (Early Pleistocene, Italy). Geobios 2023, 77, 27–36. [Google Scholar] [CrossRef]
- Forsyth Major, C.I. Studi sugli avanzi pleistocenici del genere Sus. Atti Soc. Toscana Sci. Nat. P. V. 1881, 2, 227. [Google Scholar]
- Linnaeus, C. Systema Naturae Per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis; Tomus, I., Editio Decima, R., Eds.; Laurentius Salvius: Stockholm, Sweden, 1758. [Google Scholar]
- Faure, M.; Guérin, C. Sus strozzii et Sus scrofa, deux mammiferes artiodactyles, marqueurs des paleoenvironnements. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1984, 48, 215–228. [Google Scholar] [CrossRef]
- Iannucci, A.; Gasparik, M.; Sardella, R. First report of Sus strozzii (Suidae, Mammalia) from the Early Pleistocene of Hungary (Dunaalmás) and species distinction based on deciduous teeth. Sci. Nat. 2020, 107, 5. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Navarro, B.; Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Medin, T.; Hortola, P.; Palmqvist, P. The Epivillafranchian and the arrival of pigs into Europe. Quat. Int. 2015, 389, 131–138. [Google Scholar] [CrossRef]
- Kahlke, R.-D. Untermassfeld. A Late Early Pleistocene (Epivillafranchian) Fossil Site near Meiningen (Thuringia, Germany) and Its Position in the Development of the European Mammal Fauna; British Archaeological Reports, International Series 1578; BAR Publishing: Oxford, UK, 2006. [Google Scholar]
- Kahlke, R.-D. Late Early Pleistocene European large mammals and the concept of an Epivillafranchian biochron. Cour. Forsch.-Inst. Senckenberg 2007, 259, 265–278. [Google Scholar]
- Kahlke, R.-D. Les communautés de grands mammifères du Pléistocène inférieur terminal et le concept d’un biochrone Épivillafranchien. Quaternaire 2009, 20, 415–427. [Google Scholar] [CrossRef]
- Bellucci, L.; Sardella, R.; Rook, L. Large mammal biochronology framework in Europe at Jaramillo: The Epivillafranchian as a formal biochron. Quat. Int. 2015, 389, 84–89. [Google Scholar] [CrossRef]
- Bona, F.; Sala, B. Villafranchian-Galerian mammal faunas transition in South-Western Europe. The case of the late Early Pleistocene mammal fauna of the Frantoio locality, Arda River (Castell’Arquato, Piacenza, northern Italy). Geobios 2016, 49, 329–347. [Google Scholar] [CrossRef]
- Van der Made, J.; Rosell, J.; Blasco, R. Faunas from Atapuerca at the Early-Middle Pleistocene limit: The ungulates from level TD8 in the context of climatic change. Quat. Int. 2017, 433, 296–346. [Google Scholar] [CrossRef]
- Cherin, M.; Alba, D.M.; Crotti, M.; Menconero, S.; Moullé, P.-É.; Sorbelli, L.; Madurell-Malapeira, J. The post-Jaramillo persistence of Sus strozzii (Suidae, Mammalia) in Europe: New evidence from the Vallparadís Section (NE Iberian Peninsula) and other coeval sites. Quat. Sci. Rev. 2020, 233, 106234. [Google Scholar] [CrossRef]
- Van der Made, J.; Moyà-Solà, S. European Suinae (Artiodactyla) from the Late Miocene onwards. Boll. Soc. Paleontol. Ital. 1989, 28, 329–339. [Google Scholar]
- Gliozzi, E.; Abbazzi, L.; Argenti, P.; Azzaroli, A.; Caloi, L.; Capasso Barbato, L.; Di Stefano, G.; Esu, D.; Ficcarelli, G.; Girotti, O.; et al. Biochronology of selected mammals, molluscs and ostracods from the middle Pliocene to the late Pleistocene in Italy. The state of the art. Riv. Ital. Paleontol. Stratigr. 1997, 103, 369–388. [Google Scholar] [CrossRef]
- Guérin, C.; Faure, M. The Wild Boar (Sus scrofa priscus) from the Post-Villafranchian Lower Pleistocene of Untermaßfeld. In Das Pleistozän von Untermaßfeld bei Meiningen (Thüringen). Teil 1.; Kahlke, R.-D., Ed.; Monographien des Römisch-Germanisches Zentralmuseum: Mainz, Germany, 1997; pp. 375–383. [Google Scholar]
- Lister, A.M.; Parfitt, S.A.; Owen, F.J.; Collinge, S.E.; Breda, M. Metric analysis of ungulate mammals in the early Middle Pleistocene of Britain, in relation to taxonomy and biostratigraphy: II: Cervidae, Equidae and Suidae. Quat. Int. 2010, 228, 157–179. [Google Scholar] [CrossRef]
- Antoine, P.; Moncel, M.H.; Limondin-Lozouet, N.; Locht, J.L.; Bahain, J.J.; Moreno, D.; Voinchet, P.; Auguste, P.; Stoetzel, E.; Dabkowski, J.; et al. Palaeoenvironment and dating of the Early Acheulean localities from the Somme River basin (Northern France): New discoveries from the high terrace at Abbeville-Carrière Carpentier. Quat. Sci. Rev. 2016, 149, 338–371. [Google Scholar] [CrossRef]
- Cherin, M.; Sorbelli, L.; Crotti, M.; Iurino, D.A.; Sardella, R.; Souron, A. New material of Sus strozzii (Suidae, Mammalia) from the Early Pleistocene of Italy and a phylogenetic analysis of suines. Quat. Sci. Rev. 2018, 194, 94–115. [Google Scholar] [CrossRef]
- Iannucci, A.; Sardella, R.; Strani, F.; Mecozzi, B. Size shifts in late Middle Pleistocene to Early Holocene Sus scrofa (Suidae, Mammalia) from Apulia (southern Italy): Ecomorphological adaptations? Hystrix 2020, 31, 10–20. [Google Scholar] [CrossRef]
- Luzón, C.; Yravedra, J.; Courtenay, L.A.; Saarinen, J.; Blain, H.A.; DeMiguel, D.; Viranta, S.; Azanza, B.; Rodrigez-Alba, J.J.; Herranz-Rodrigo, D.; et al. Taphonomic and spatial analyses from the early Pleistocene site of Venta Micena 4 (Orce, Guadix-Baza Basin, southern Spain). Sci. Rep. 2021, 11, 13977. [Google Scholar] [CrossRef]
- Valli, A.M.F.; Caron, J.-B.; Debard, E.; Guérin, C.; Pastre, J.-F.; Argant, J. Le gisement paléontologique villafranchien terminal de Peyrolles (Issoire, Puy-de-Dôme, France): Résultats de nouvelles prospections. Geodiversitas 2006, 28, 297–317. [Google Scholar]
- Bout, P.; Azzaroli, A. Stratigraphie et faune du Creux de Peyrolles près Perrier (Puy-de-Dôme). Annal. Paloentol. 1952, 38, 37–56. [Google Scholar]
- Bout, P. Le Villafranchien du Velay et du Basin Hydrographique Moyen et Supérieur de L’allier; Imprimerie Jeanne d’Arc: Le Puy-en-Velay, France, 1960. [Google Scholar]
- Couthures, J.; Pastre, J.F. Chronostratigraphie du Plio-PIéistocène d’Auvergne et du Velay: Nouveaux apports des datations radiométriques et du paléomagnétisme. Quaternaire 1983, 20, 9–18. [Google Scholar] [CrossRef]
- Poidevin, J.L.; Cantagrel, J.M.; GUERPA. Un site unique du Plio-Pleistocene en Europe: Le pluteau de Perrier (Puy-de-Dôme)—Confrontation des donneés volcanologiques, stratigraphiques et paléontologiques. Rev. Sc. Nat. d’Auvergne 1984, 50, 87–95. [Google Scholar]
- Nomade, S.; Pastre, J.-F.; Guillou, H.; Faure, M.; Guérin, C.; Delson, E.; Debard, E.; Voinchet, P.; Msessager, E. 40Ar/39Ar constraints on some French landmark Late Pliocene to Early Pleistocene large mammalian paleofaunas: Paleoenvironmental and paleoecological implications. Quat. Geochronol. 2014, 21, 2–15. [Google Scholar] [CrossRef]
- Lo Bello, P. Géochronologie par la Méthode 39Ar-40Ar de Ponces Quaternaires Contaminées: Exemple des Ponces du Mont-Dore (Massif Central, France). Utilisation d’un Laser Continu Pour la Datation des Min´Eraux Individuels. Unpublished Ph.D. Thesis, Université de Nice, Nice, France, 1988. [Google Scholar]
- Palombo, M.R.; Valli, A.M.F. Remarks on the biochronology of mammalian faunal complexes from the Pliocene to the Middle Pleistocene in France. Geol. Romana 2004, 37, 145–163. [Google Scholar]
- Thouveny, N.; Taieb, M.; Bonnet, A. Etude magnétostratigraphique du remplissage du Pléistocene inférieur de la grotte de La Sartanette (Remoulins, Gard). Bull. Soc. Geol. Fr. 1984, 26, 1385–1388. [Google Scholar] [CrossRef]
- Paquette, J.L.; Médard, E.; Poidevin, J.L.; Barbet, P. Precise dating of middle to late Villafranchian mammalian paleofaunae from the Upper Allier River valley (French Massif Central) using U-Pb geochronology on volcanic zircons. Quat. Geochronol. 2021, 65, 101198. [Google Scholar] [CrossRef]
- Heintz, E.; Guérin, C.; Martin, P.; Prat, F. Principaux gisements Villafranchiens de France: Listes fauniques et biostratigraphie. Mem. BRGM 1974, 78, 411–417. [Google Scholar]
- Iannucci, A.; Mecozzi, B.; Sardella, R.; Iurino, D.A. The extinction of the giant hyena Pachycrocuta brevirostris and a reappraisal of the Epivillafranchian and Galerian Hyaenidae in Europe: Faunal turnover during the Early–Middle Pleistocene Transition. Quat. Sci. Rev. 2021, 272, 107240. [Google Scholar] [CrossRef]
- Cherin, M.; Breda, M.; Esattore, B.; Hart, V.; Turek, J.; Porciello, F.; Angeli, G.; Holpin, S.; Iurino, D.A. A Pleistocene Fight Club revealed by the palaeobiological study of the Dama-like deer record from Pantalla (Italy). Sci. Rep. 2022, 12, 13898. [Google Scholar] [CrossRef]
- Soria, C.D.; Pacifici, M.; Di Marco, M.; Stephen, S.M.; Rondinini, C. COMBINE: A coalesced mammal database of intrinsic and extrinsic traits. Ecology 2021, 102, e03344. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 4. [Google Scholar]
- Mazza, P.P.A.; Bertini, A.; Magi, M. The late Pliocene site of Poggio Rosso (central Italy): Taphonomy and paleoenvironment. Palaios 2004, 19, 227–248. [Google Scholar] [CrossRef]
- Barash, A.; Belmaker, M.; Bastir, M.; Soudack, M.; O’Brien, H.D.; Woodward, H.; Prendergast, A.; Barzilai, O.; Been, E. The earliest Pleistocene record of a large-bodied hominin from the Levant supports two out-of-Africa dispersal events. Sci. Rep. 2022, 12, 1721. [Google Scholar] [CrossRef] [PubMed]
- Kahlke, R.-D. (Ed.) The Pleistocene of Untermassfeld—A synopsis of site origin, palaeobiodiversity, taphonomic characteristics, palaeoenvironment, chronostratigraphy, and significance in western Palaearctic faunal history. In The Pleistocene of Untermassfeld near Meiningen (Thüringen, Germany); Part 5; Monographien des Römisch-Germanisches Zentralmuseum: Mainz, Germany, 2022; pp. 1635–1734. [Google Scholar]
- Madurell-Malapeira, J.; Alba, D.M.; Espigares, M.-P.; Vinuesa, V.; Palmqvist, P.; Martínez-Navarro, B.; Moyà-Solà, S. Were large carnivorans and great climatic shifts limiting factors for hominin dispersals? Evidence of the activity of Pachycrocuta brevirostris during the Mid-Pleistocene Revolution in the Vallparadís Section (Vallès-Penedès Basin, Iberian Peninsula). Quat. Int. 2017, 431, 42–52. [Google Scholar] [CrossRef]
- Tchernov, E. The age of the ‘Ubeidiya Formation, an Early Pleistocene hominid site in the Jordan Valley, Israel. Isr. J. Earth Sci. 1987, 36, 3–30. [Google Scholar]
- Martínez-Navarro, B.; Belmaker, M.; Bar-Yosef, O. The large carnivores from ‘Ubeidiya (early Pleistocene, Israel): Biochronological and biogeographical implications. J. Hum. Evol. 2009, 56, 514–524. [Google Scholar] [CrossRef]
- Belmaker, M. Insights from carnivore community composition on the paleoecology of early Pleistocene Eurasian sites: Implications for the dispersal of hominins out of Africa. Quat. Int. 2018, 464, 3–17. [Google Scholar] [CrossRef]
- Kostopoulos, D.S.; Konidaris, G.E.; Amanatidou, M.; Chitoglou, K.; Fragkioudakis, E.; Gerakakis, N.; Giannakou, V.; Gkeme, A.; Kalaitzi, C.; Tsakalidis, C.; et al. The new fossil site Krimni-3 in Mygdonia Basin and the first evidence of a giant ostrich in the Early Pleistocene of Greece. PalZ 2023, 97, 147–161. [Google Scholar] [CrossRef]
- De Giuli, C.; Masini, F. Late Villafranchian faunas in Italy: The Casa Frata local fauna (Upper Valdarno, Tuscany). Palaeontogr. Ital. 1986, 74, 1–9. [Google Scholar]
- Napoleone, G.; Albanielli, A.; Azzaroli, A.; Bertini, A.; Magi, M.; Mazzini, M. Calibration of the Upper Valdarno Basin to the Plio-Pleistocene for correlating the Apennine continental sequences. Ital. J. Quat. Sci. 2003, 16, 131–166. [Google Scholar]
- Iannucci, A.; Conti, J.; Curcio, F.; Iurino, D.A.; Mancini, M.; Mecozzi, B.; Strani, F.; Sardella, R. Middle Pleistocene mammal faunas of the area of Rome: Recent results and ongoing work on the MUST collection. J. Mediterr. Earth Sci. 2023, 15, 19–37. [Google Scholar] [CrossRef]
- Duval, M.; Arnold, L.J.; Demuro, M.; Parés, J.M.; Campaña, I.; Carbonell, E.; de Castro, J.M.B. New chronological constraints for the lowermost stratigraphic unit of Atapuerca Gran Dolina (Burgos, N Spain). Quat. Geochron. 2022, 71, 101292. [Google Scholar] [CrossRef]
- Iannucci, A.; Cherin, M.; Sardella, R. The lost hyena from Paciano (Umbria, Italy) reconsidered. Alp. Mediterr. Quat. 2022, 35, 105–117. [Google Scholar]
- Masini, F.; Palombo, M.R.; Rozzi, R. A reappraisal of the early to middle Pleistocene Italian Bovidae. Quat. Int. 2013, 288, 45–62. [Google Scholar] [CrossRef]
- Sorbelli, L.; Cherin, M.; Kostopoulos, D.S.; Sardella, R.; Mecozzi, B.; Plotnikov, V.; Prat-Vericat, M.; Azzarà, B.; Bartolini-Lucenti, S.; Madurell-Malapeira, J. Earliest bison dispersal in Western Palearctic: Insights from the Eobison record from Pietrafitta (Early Pleistocene, central Italy). Quat. Sci. Rev. 2023, 301, 107923. [Google Scholar] [CrossRef]
- Siori, M.S.; Sala, B. The mammal fauna from the late Early Biharian site of Castagnone (Northern Monferrato, Piedmont, NW Italy). Geobios 2007, 40, 207–217. [Google Scholar] [CrossRef]
- Guérin, C.; Dewolf, Y.; Lautridou, J.P. Révision d’un site paléontologique célèbre: Saint-Prest (Chartres, France). Geobios 2003, 36, 55–82. [Google Scholar] [CrossRef]
- Owen, R. Description of teeth and portions of jaws of two extinct anthracotherioid quadrupeds (Hyopotamus vectianus and Hyop. bovinus) discovered by the Marchioness of Hastings in the Eocene deposits on the N. W. coast of the Isle of Wight: With an attempt to develop Cuvier’s idea of the classification of pachyderms by the number of their toes. Q. J. Geol. Soc. Lond. 1848, 4, 103–141. [Google Scholar]
- Gray, J.E. On the natural arrangement of vertebrose animals. London Medical Reposit. Monthly J. Rev. 1821, 15, 296–310. [Google Scholar]
- Iannucci, A. New results on suids from the Early Pleistocense site of Untermassfeld. In The Pleistocene of Untermassfeld near Meiningen (Thüringen, Germany); Part 5; Kahlke, R.-D., Ed.; Monographien des Römisch-Germanisches Zentralmuseum: Mainz, Germany, 2022; pp. 1339–1354. [Google Scholar]
- Schaub, S. Die oberpliocaene Säugetierfauna von Senèze (Haute-Loire) und ihre verbreitungsgeschichtliche Stellung. Eclogae Geol. Helv. 1943, 36, 270–289. [Google Scholar]
- Azzaroli, A. Revisione dei terreni fluvio-lacustri del Valdarno Superiore. V. Filogenesi e biologia di Sus strozzii e Sus minor. Palaeontogr. Ital. 1954, 48, 41–76. [Google Scholar]
- Dubost, G.; Charron, F.; Courcoul, A.; Rodier, A. The Chinese water deer, Hydropotes inermis—A fast-growing and productive ruminant. Mammal. Biol. 2011, 76, 190–195. [Google Scholar] [CrossRef]
- Frauendorf, M.; Gethöffer, F.; Siebert, U.; Keuling, O. The influence of environmental and physiological factors on the litter size of wild boar (Sus scrofa) in an agriculture dominated area in Germany. Sci. Total Environ. 2016, 541, 877–882. [Google Scholar] [CrossRef]
- Pianka, E.R. On r- and K-Selection. Am. Nat. 1970, 104, 592–597. [Google Scholar] [CrossRef]
- Tuomi, J. Mammalian reproductive strategies: A generalized relation of litter size to body size. Oecologia 1980, 45, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Stearns, S.C. Life history tactics: A review of the ideas. Quart. Rev. Biol. 1976, 51, 3–47. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D.; Bryant, M.J.; Bashey, F. r-and K-selection revisited: The role of population regulation in life-history evolution. Ecology 2002, 83, 1509–1520. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. An equilibrium theory of insular zoogeography. Evolution 1963, 17, 373–387. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Bieber, C.; Ruf, T. Population dynamics in wild boar Sus scrofa: Ecology, elasticity of growth rate and implications for the management of pulsed resource consumers. J. Appl. Ecol. 2005, 42, 1203–1213. [Google Scholar] [CrossRef]
- Geisser, H.; Reyer, H.-U. The influence of food and temperature on population density of wild boar Sus scrofa in the Thurgau (Switzerland). J. Zool. 2005, 267, 89–96. [Google Scholar] [CrossRef]
- Keuling, O.; Baubet, E.; Duscher, A.; Ebert, C.; Fischer, C.; Monaco, A.; Podgórski, T.; Prevot, C.; Ronnenberg, K.; Sodeikat, G.; et al. Mortality rates of wild boar Sus scrofa L. in central Europe. Eur. J. Wildl. Res. 2013, 59, 805–814. [Google Scholar] [CrossRef]
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef]
- Vetter, S.G.; Ruf, T.; Bieber, C.; Arnold, W. What is a mild winter? Regional differences in within-species responses to climate change. PLoS ONE 2015, 10, e0132178. [Google Scholar] [CrossRef] [PubMed]
- Melletti, M.; Meijaard, E. Ecology, Conservation and Management of Wild Pigs and Peccaries; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- White, T.D.; Harris, J.M. Suid evolution and correlation of African hominid localities. Science 1977, 198, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Cooke, H.B.S.; Wilkinson, A.F. Suidae and Tayassuidae. In Evolution of African Mammals; Maglio, V.J., Cooke, H.B.S., Eds.; Harvard University Press: Cambridge, UK, 1978; pp. 435–482. [Google Scholar]
- Harris, J.M.; Cerling, T.E. Dietary adaptations of extant and Neogene African suids. J. Zool. 2002, 256, 45–54. [Google Scholar] [CrossRef]
- Harris, J.M.; White, T.D. Evolution of the Plio-Pleistocene African Suidae. Trans. Am. Philos. Soc. 1979, 69, 1–128. [Google Scholar] [CrossRef]
- Souron, A. Diet and ecology of extant and fossil wild pigs. In Ecology, Conservation and Management of Wild Pigs and Peccaries; Melletti, M., Meijaard, E., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 29–38. [Google Scholar]
- Yang, D.; Pisano, A.; Kolasa, J.; Jashashvili, T.; Kibii, J.; Cano, A.R.G.; Viriot, N.; Grine, F.E.; Souron, A. Why the long teeth? Morphometric analysis suggests different selectivse pressures on functional occlusal traits in Plio-Pleistocene African suids. Paleobiology 2022, 48, 655–676. [Google Scholar] [CrossRef]
- Cherin, M.; Basilici, G.; Duval, M.; Shao, Q.; Sier, M.J.; Parés, J.M.; Gliozzi, E.; Mazzini, I.; Magri, D.; Di Rita, F.; et al. The dawn of the Late Villafranchian: Paleoenvironment and age of the Pantalla paleontological site (Italy; Early Pleistocene). Quat. Sci. Rev. 2023, 316, 108279. [Google Scholar] [CrossRef]
- Strani, F.; Bellucci, L.; Iannucci, A.; Iurino, D.A.; Mecozzi, B.; Sardella, R. Palaeoenvironments of the MIS 15 site of Cava di Breccia—Casal Selce 2 (central Italian Peninsula) and niche occupation of fossil ungulates during Middle Pleistocene interglacials. Hist. Biol. 2022, 34, 555–565. [Google Scholar] [CrossRef]
- Magurran, A.E.; Henderson, P.A. Explaining the excess of rare species in natural species abundance distributions. Nature 2003, 422, 714–716. [Google Scholar] [CrossRef]
- Sorbelli, L.; Alba, D.M.; Cherin, M.; Moullé, P.É.; Brugal, J.P.; Madurell-Malapeira, J. A review on Bison schoetensacki and its closest relatives through the early-Middle Pleistocene transition: Insights from the Vallparadís Section (NE Iberian Peninsula) and other European localities. Quat. Sci. Rev. 2021, 261, 106933. [Google Scholar] [CrossRef]
- Hermier, R.; Merceron, G.; Kostopoulos, D.S. The emblematic Eurasian Villafranchian antelope Gazellospira (Mammalia: Bovidae): New insights from the Lower Pleistocene Dafnero fossil sites (Northern Greece). Geobios 2020, 61, 11–29. [Google Scholar] [CrossRef]
- Palombo, M.R. Discrete dispersal bioevents of large mammals in Southern Europe in the post-Olduvai Early Pleistocene: A critical overview. Quat. Int. 2017, 431, 3–19. [Google Scholar] [CrossRef]
- Breda, M.; Marchetti, M. Systematical and biochronological review of Plio-Pleistocene Alceini (Cervidae; Mammalia) from Eurasia. Quat. Sci. Rev. 2005, 24, 775–805. [Google Scholar] [CrossRef]
- Mazza, P.P.A. Poggio Rosso (Upper Valdarno, central Italy), a window on latest Pliocene wildlife. Palaios 2006, 21, 493–498. [Google Scholar] [CrossRef]
- Moncel, M.-H.; Lemorini, C.; Eramo, G.; Fioretti, G.; Daujeard, C.; Curci, A.; Berto, C.; Hardy, B.; Pineda, A.; Rineau, V.; et al. A taphonomic and spatial distribution study of the new levels of the middle Pleistocene site of Notarchirico (670–695 ka, Venosa, Basilicata, Italy). Archaeol. Anthropol. Sci. 2023, 15, 106. [Google Scholar] [CrossRef]
- Ollé, A.; Lombao, D.; Asryan, L.; Gárcia-Medano, P.; Arroyo, A.; Fernández-Marchena, J.L.; Yesilova, G.C.; Cáceres, I.; Huguet, R.; López-Polín, L.; et al. The earliest European Acheulean: New insights into the large shaped tools from the late Early Pleistocene site of Barranc de la Boella (Tarragona, Spain). Front. Earth Sci. 2023, 11, 1188663. [Google Scholar] [CrossRef]
- Mayer, J.J.; Brisbin, I.E. Wild Pigs: Biology, Damage, Control Techniques and Management; Savannah River National Laboratory: Aiken, SC, USA, 2009. [Google Scholar]
- Pazzaglia, F.; Barchi, M.R.; Buratti, N.; Cherin, M.; Pandolfi, L.; Ricci, M. Pleistocene calcareous tufa from the Ellera basin (Umbria, central Italy) as a key for an integrated paleoenvironmental and tectonic reconstruction. Quat. Int. 2013, 292, 59–70. [Google Scholar] [CrossRef]
- Abbazzi, L.; Benvenuti, M.; Rook, L.; Masini, F. Biochronology of the Mugello intermontane basin (northern Apennines, Italy). Il Quat. 1995, 8, 5–10. [Google Scholar]
- Fidalgo, D.; Rosas, A.; Madurell-Malapeira, J.; Pineda, A.; Huguet, R.; García-Tabernero, A.; Cáceres, I.; Olle, A.; Vallverdú, J.; Saladie, P. A review on the Pleistocene occurrences and palaeobiology of Hippopotamus antiquus based on the record from the Barranc de la Boella Section (Francolí Basin, NE Iberia). Quat. Sci. Rev. 2023, 307, 108034. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Palombo, M.R. The late Early to early Middle Pleistocene stenonoid horses from Italy. Quat. Int. 2013, 288, 25–44. [Google Scholar] [CrossRef]
- De Giuli, C. Late Villafranchian faunas of Italy: The Selvella local fauna in the southern Chiana Valley-Umbria. Palaeontogr. Ital. 1986, 74, 11–50. [Google Scholar]
- Geraads, D.; Guérin, C.; Faure, M. Les suidés du Pléistocène ancien d’Oubeidiyeh (Israël). Mem. Travaux Cent. Rech. Fr. Jerus. 1986, 5, 93–105. [Google Scholar]
- Rink, W.J.; Bartoll, J.; Schwarcz, H.P.; Shane, P.; Bar-Yosef, O. Testing the reliability of ESR dating of optically exposed buried quartz sediments. Radiat. Meas. 2007, 42, 1618–1626. [Google Scholar] [CrossRef]
- Freudenthal, M. Neogene vertebrates from the Gargano Peninsula, Italy. Scripta Geol. 1971, 3, 1–10. [Google Scholar]
- De Giuli, C.; Masini, F.; Torre, D. The latest Villafranchian faunas in Italy: The Pirro Nord fauna (Apricena, Gargano). Palaeontogr. Ital. 1986, 74, 51–62. [Google Scholar]
- Arzarello, M.; Marcolini, F.; Pavia, G.; Pavia, M.; Petronio, C.; Petrucci, M.; Rook, L.; Sardella, R. Evidence of earliest human occurrence in Europe: The site of Pirro Nord (southern Italy). Naturwissenschaften 2007, 94, 107–112. [Google Scholar] [CrossRef]
- Arzarello, M.; Pavia, G.; Peretto, C.; Petronio, C.; Sardella, R. Evidence of an early Pleistocene hominin presence at Pirro Nord (Apricena, Foggia, southern Italy): P13 site. Quat. Int. 2012, 267, 56–61. [Google Scholar] [CrossRef]
- Arzarello, M.; De Weyer, L.; Peretto, C. The first European peopling and the Italian case: Peculiarities and “opportunism”. Quat. Int. 2016, 393, 41–50. [Google Scholar] [CrossRef]
- Pavia, M.; Zunino, M.; Coltorti, M.; Angelone, C.; Arzarello, M.; Bagnus, C.; Bellucci, L.; Colombero, S.; Marcolini, F.; Peretto, C.; et al. Stratigraphical and palaeontological data from the Early Pleistocene Pirro 10 site of Pirro Nord (Puglia, south eastern Italy). Quat. Int. 2012, 267, 40–55. [Google Scholar] [CrossRef]
- Petrucci, M.; Cipullo, A.; Martínez-Navarro, B.; Rook, L.; Sardella, R. The Late Villafranchian (Early Pleistocene) carnivores (Carnivora, Mammalia) from Pirro Nord (Italy). Palaeontogr. Abt. A 2013, 298, 113–145. [Google Scholar] [CrossRef]
- Carpentieri, M.; Arzarello, M. For our world without sound: The opportunistic debitage in the Italian context—A methodological evaluation of the lithic assemblages of Pirro Nord, Cà Belvedere di Montepoggiolo, Ciota Ciara Cave and Riparo Tagliente. J. Paleolithic Archaeol. 2022, 5, 12. [Google Scholar] [CrossRef]
- Stefanelli, D.; Mecozzi, B.; Breda, M.; Girone, A.; La Perna, R.; Maiorano, P.; Marino, M.; Sardella, R. Paleodays 2023—XXIII Edizione Delle Giornate di Paleontologia Lecce, 7–9 Giugno 2023, Abstract Book; Cherin, M., Collareta, A., Eds.; Rediscovering the Fossil Collection from the Late Early Pleistocene Site of Pirro Nord (Foggia, Southern Italy) at the Bari University; University of Salento: Lecce, Italy, 2023; p. 76. [Google Scholar]
- Tsoukala, E.; Bonifay, M.-F. The Early Pleistocene Carnivores (Mammalia) from Ceyssaguet (Haute-Loire). Paléo 2004, 16, 193–242. [Google Scholar]
- Brugal, J.P.; Argant, A.; Boudadi-Maligne, M.; Crégut-Bonnoure, E.; Croitor, R.; Fernandez, P.; Fourvel, J.-P.; Fosse, P.; Gaudelli, J.-L.; Labe, B.; et al. Pleistocene herbivores and carnivores from France: An updated overview of the literature, sites and taxonomy. Ann. Paleontol. 2020, 106, 102384. [Google Scholar] [CrossRef]
- Koufos, G.D.; Kostopoulos, D.S. The Plio-Pleistocene large mammal record of Greece: Implications for early human dispersals into Europe In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 269–280. [Google Scholar]
- Konidaris, G.E.; Kostopoulos, D.S.; Maron, M.; Schaller, M.; Ehlers, T.A.; Aidona, E.; Marini, M.; Tourloukis, V.; Muttoni, G.; Koufos, G.D.; et al. Dating of the Lower Pleistocene vertebrate site of Tsiotra Vryssi (Mygdonia Basin, Greece): Biochronology, magnetostratigraphy, and cosmogenic radionuclides. Quaternary 2021, 4, 1. [Google Scholar] [CrossRef]
- Alifieri, E. Dietary Adaptations of the Early Pleistocene Gerakarou Mammal Community (Mygdonia Basin, Greece). Unpublished Master’s Thesis, Interinstitutional Program of Postgraduate Studies in Palaeontology-Geobiology, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2021. [Google Scholar]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef]
- Vekua, A. Die Wirbeltierfauna des Villafranchium von Dmanisi und ihre biostratigraphische Bedeutung. Jahrb. RGZM 1995, 42, 77–180. [Google Scholar]
- Channell, J.E.T.; Singer, B.S.; Jicha, B.R. Timing of Quaternary geomagnetic reversals and excursions in volcanic and sedimentary archives. Quat. Sci. Rev. 2020, 228, 106114. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
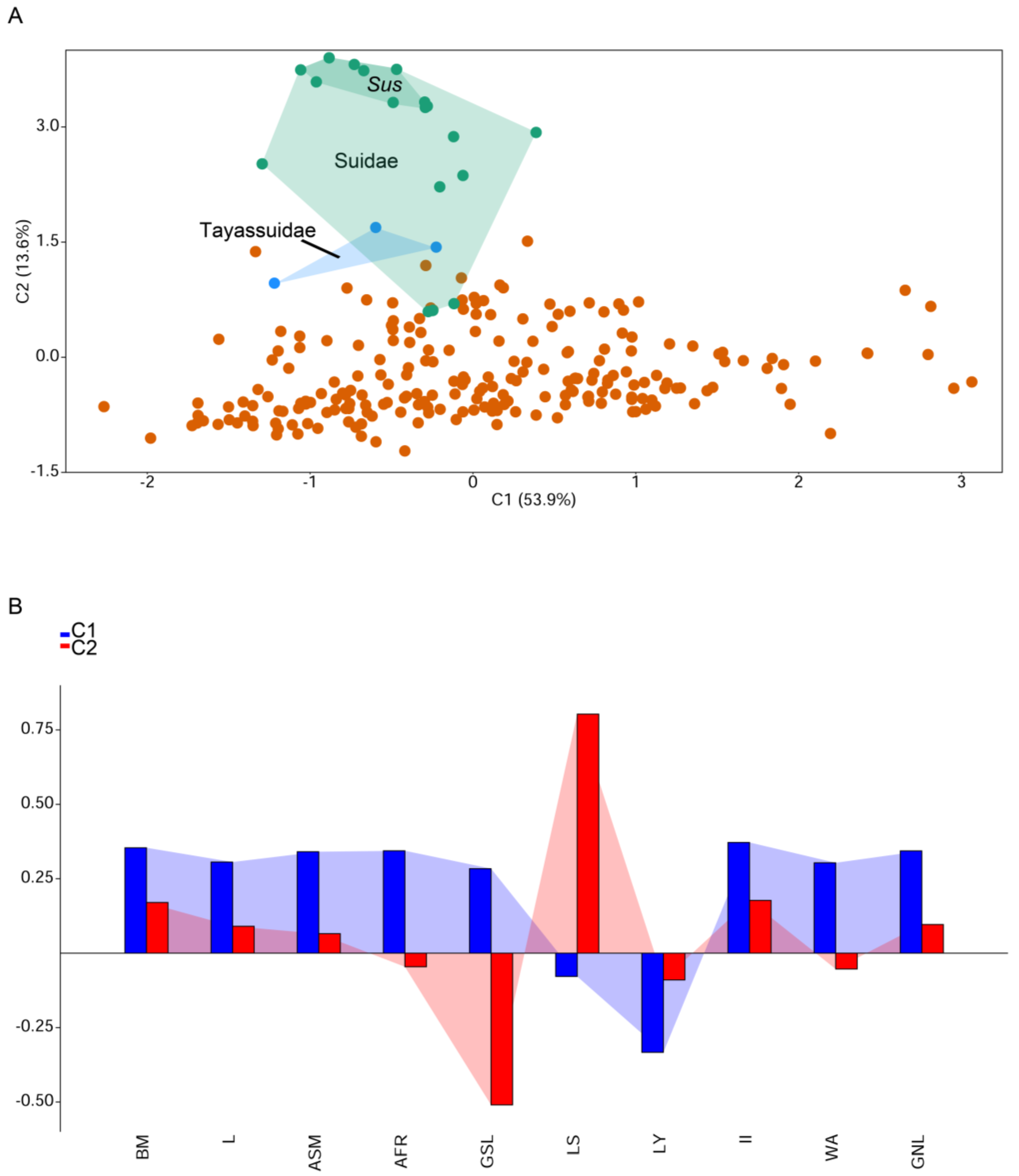
| Variable | Abbreviation | Description |
|---|---|---|
| Body mass | BM | Body mass of an adult individual in grams |
| Longevity | L | Maximum reported age at death for the species in days |
| Age at female sexual maturity | ASM | The amount of time needed for a female to reach sexual maturity in days |
| Age at first reproduction | AFR | Age at which females give birth to their first litter or their young attach to teats in days |
| Gestation length | GSL | Length of time of fetal growth in days |
| Litter size | LS | Number of offspring born per litter per female |
| Litters per year | LY | Number of litters per female per year |
| Interbirth interval | II | Time between reproduction events in days |
| Weaning age | WA | Age at which primary nutritional dependency on the mother ends and independent foraging begins in days |
| Generation length | GNL | Average age of parents of the current cohort in days |
| PCA | PC1 | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|
| % Variance | 5.38938 | 1.35783 | 0.75328 | 0.677878 | 0.506962 |
| BM | 0.35411 | 0.17025 | 0.2838 | −0.16842 | 0.37325 |
| L | 0.30613 | 0.090422 | 0.45011 | 0.40485 | −0.43725 |
| ASM | 0.34086 | 0.065646 | −0.42337 | 0.091518 | 0.39679 |
| AFR | 0.344 | −0.04563 | −0.204 | 0.38143 | 0.35725 |
| GSL | 0.28391 | −0.50972 | 0.040859 | −0.12475 | −0.06277 |
| LS | −0.078169 | 0.80313 | 0.057934 | −0.074333 | 0.044958 |
| LY | −0.33322 | −0.089843 | 0.382 | 0.29656 | 0.54834 |
| II | 0.37192 | 0.17733 | −0.28008 | −0.25519 | −0.18045 |
| WA | 0.30359 | −0.053162 | 0.49521 | −0.52393 | 0.17881 |
| GNL | 0.3437 | 0.096293 | 0.1462 | 0.45299 | −0.11515 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iannucci, A. The Occurrence of Suids in the Post-Olduvai to Pre-Jaramillo Pleistocene of Europe and Implications for Late Villafranchian Biochronology and Faunal Dynamics. Quaternary 2024, 7, 11. https://doi.org/10.3390/quat7010011
Iannucci A. The Occurrence of Suids in the Post-Olduvai to Pre-Jaramillo Pleistocene of Europe and Implications for Late Villafranchian Biochronology and Faunal Dynamics. Quaternary. 2024; 7(1):11. https://doi.org/10.3390/quat7010011
Chicago/Turabian StyleIannucci, Alessio. 2024. "The Occurrence of Suids in the Post-Olduvai to Pre-Jaramillo Pleistocene of Europe and Implications for Late Villafranchian Biochronology and Faunal Dynamics" Quaternary 7, no. 1: 11. https://doi.org/10.3390/quat7010011
APA StyleIannucci, A. (2024). The Occurrence of Suids in the Post-Olduvai to Pre-Jaramillo Pleistocene of Europe and Implications for Late Villafranchian Biochronology and Faunal Dynamics. Quaternary, 7(1), 11. https://doi.org/10.3390/quat7010011