
The Effect of Seeding Treatments and Climate on Fire Regimes in Wyoming Sagebrush Steppe


Abstract
:1. Introduction
2. Materials and Methods
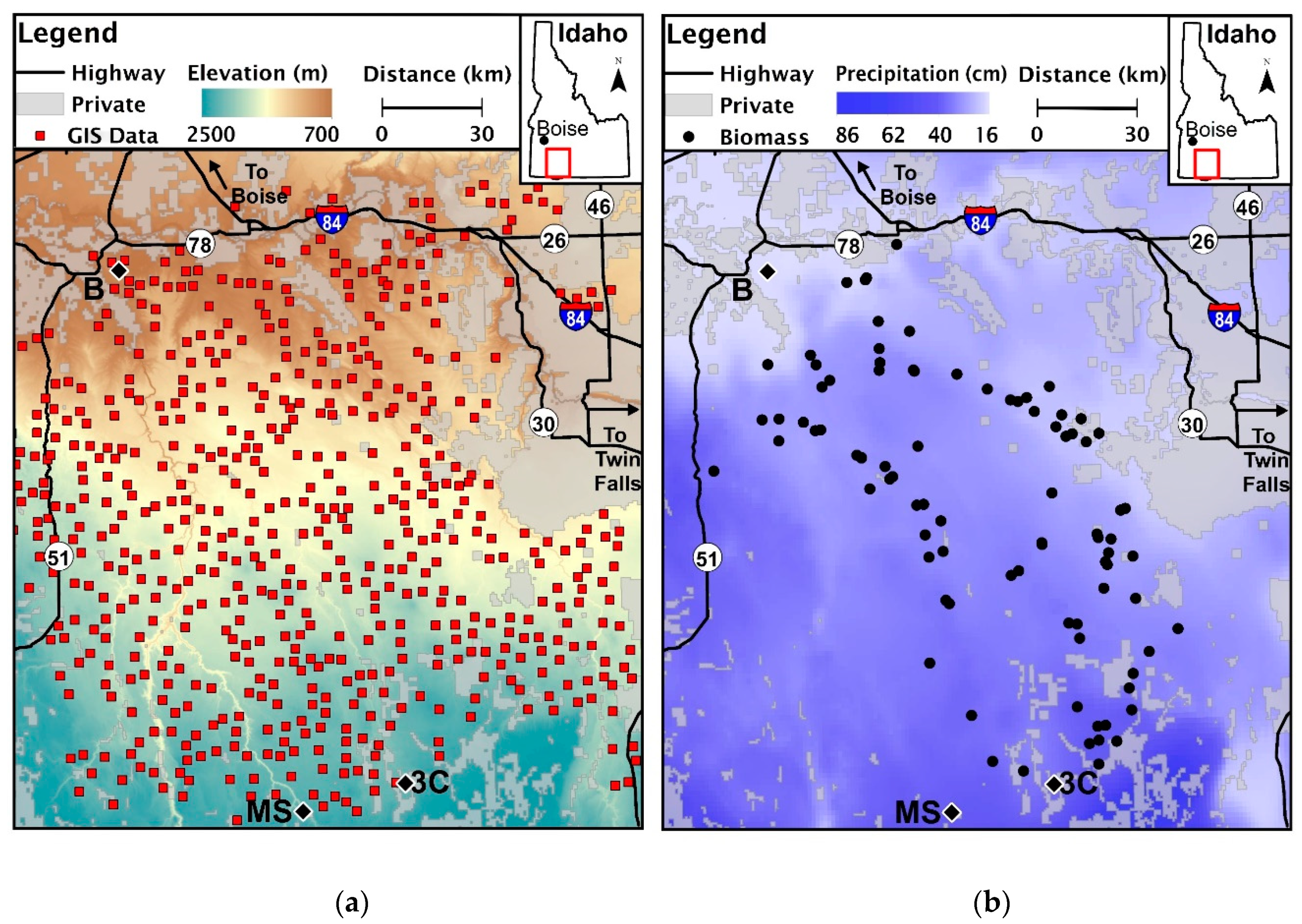
2.1. Site Description
2.2. Data Extraction
2.3. Field Collection
2.4. Statistical Analysis
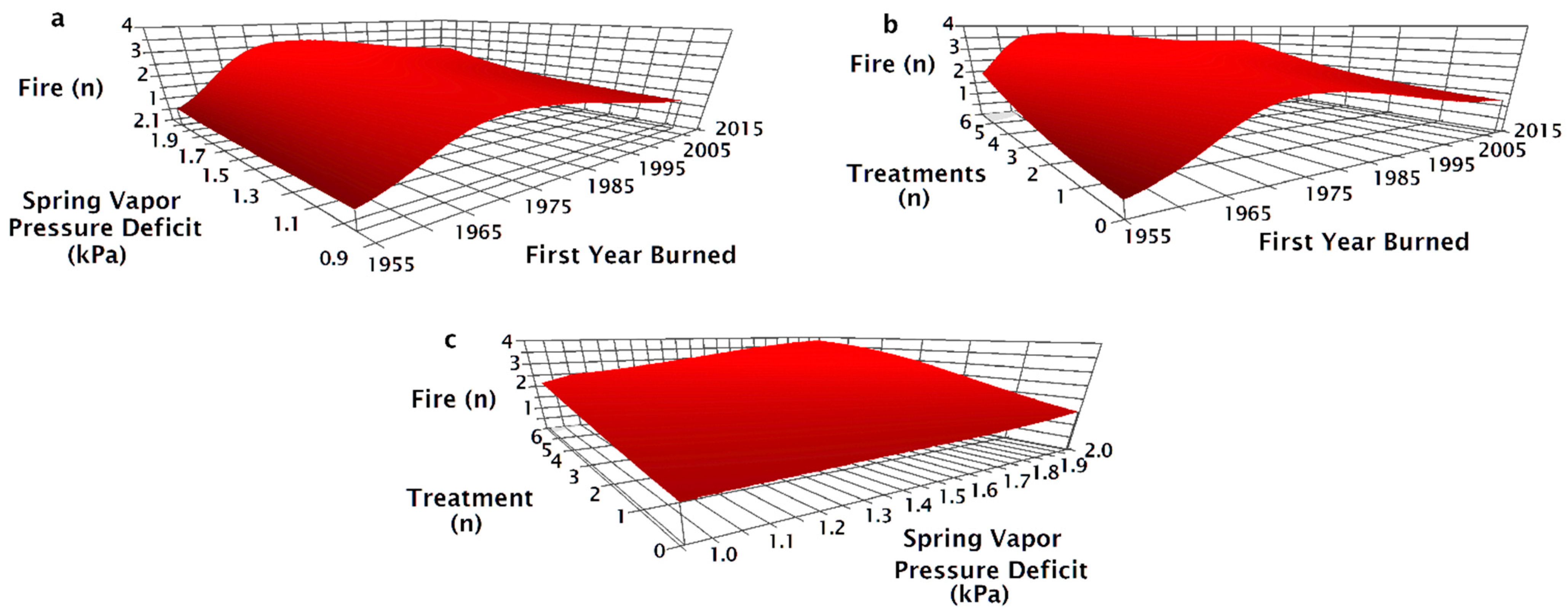
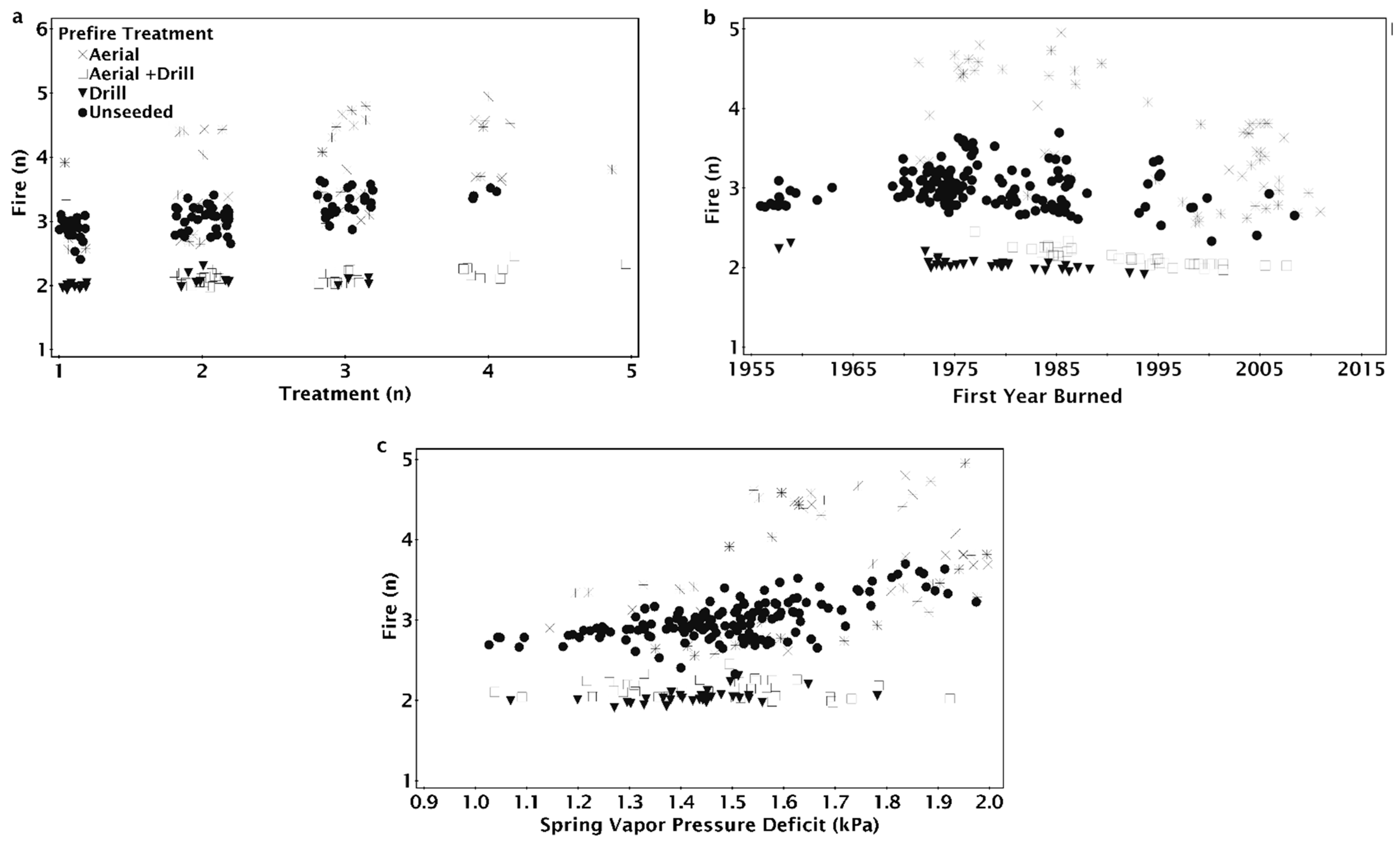
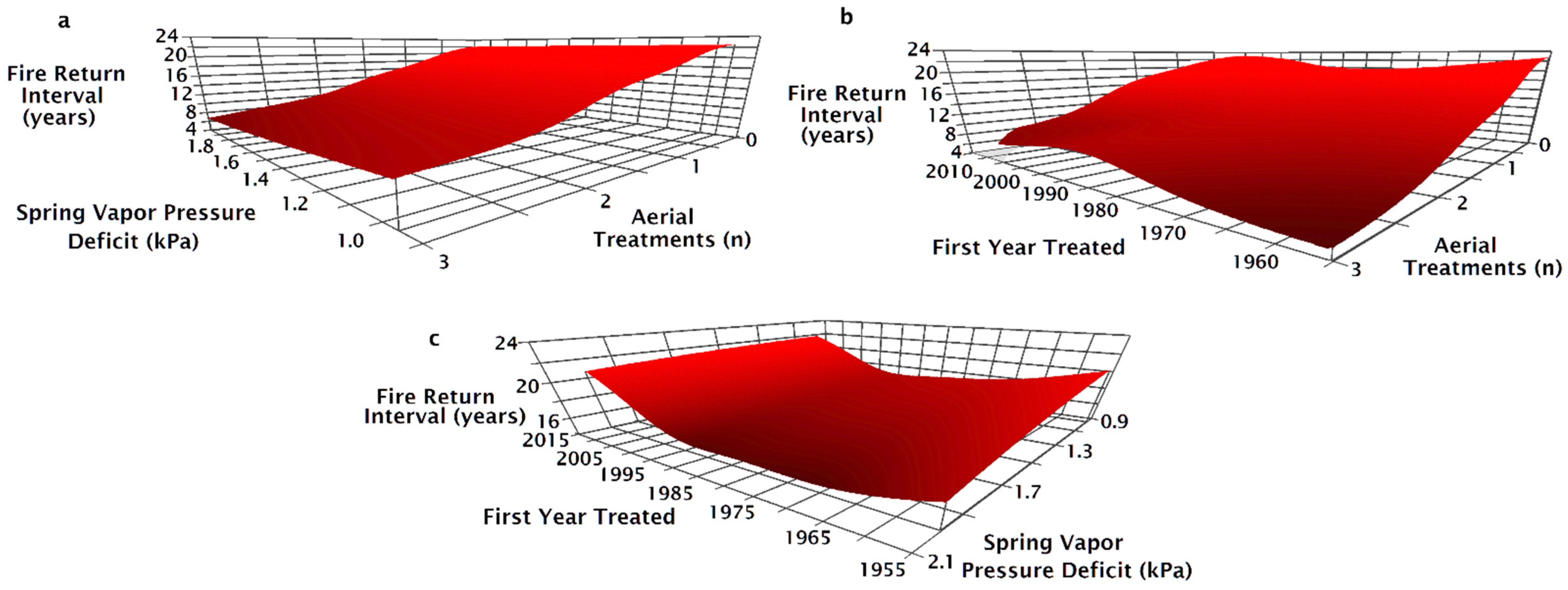
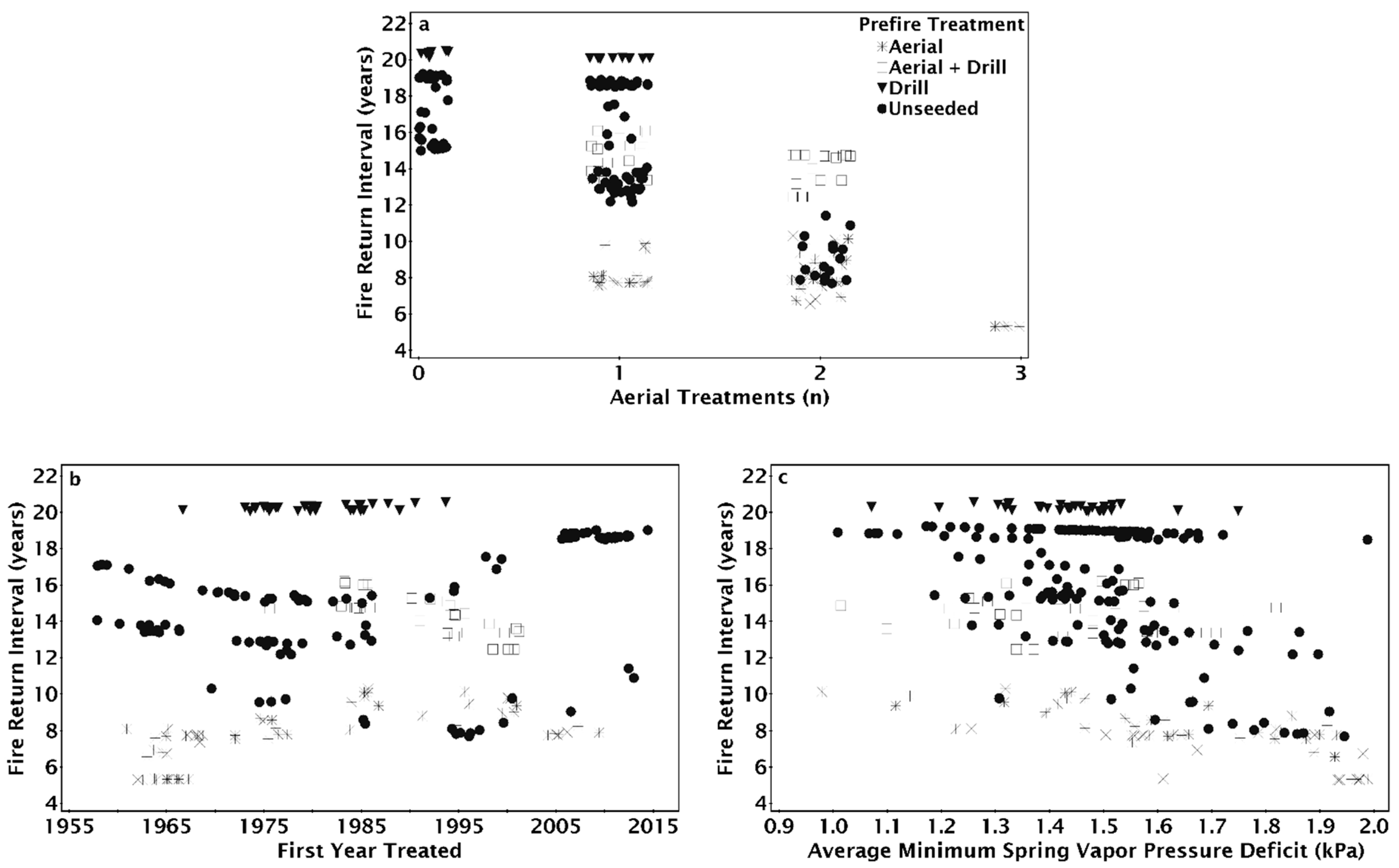
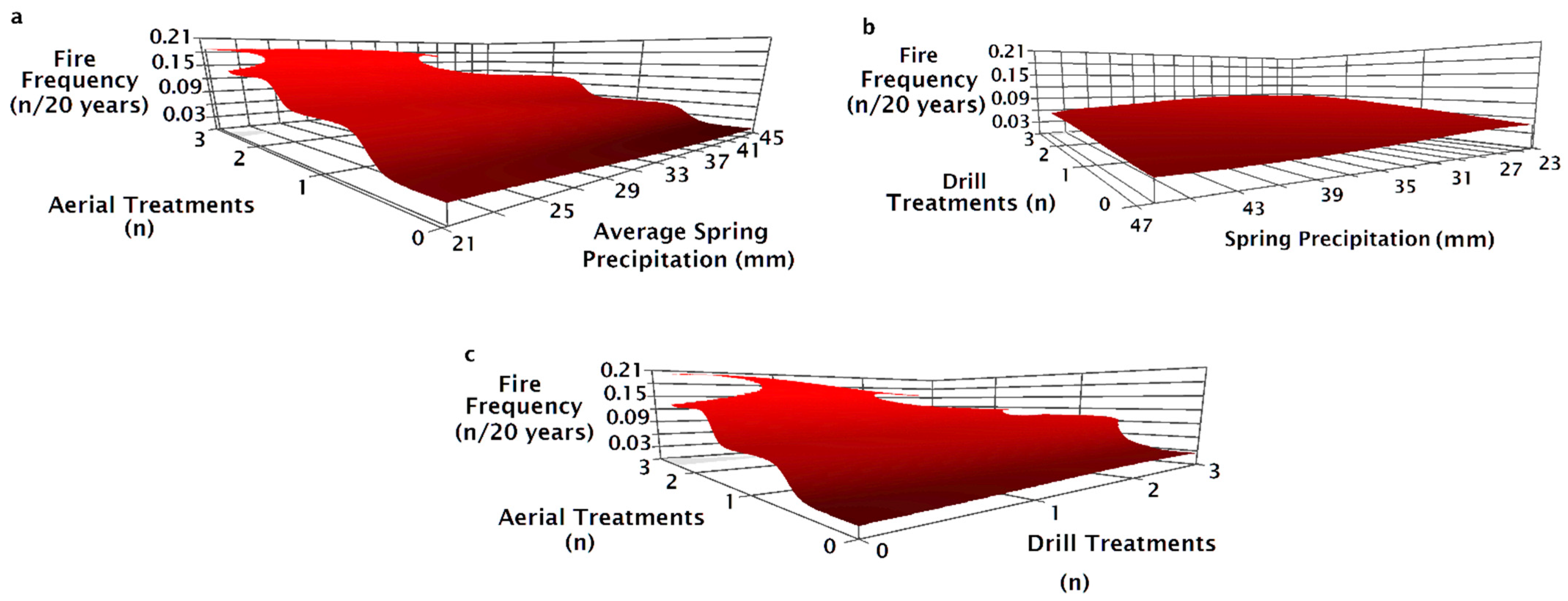
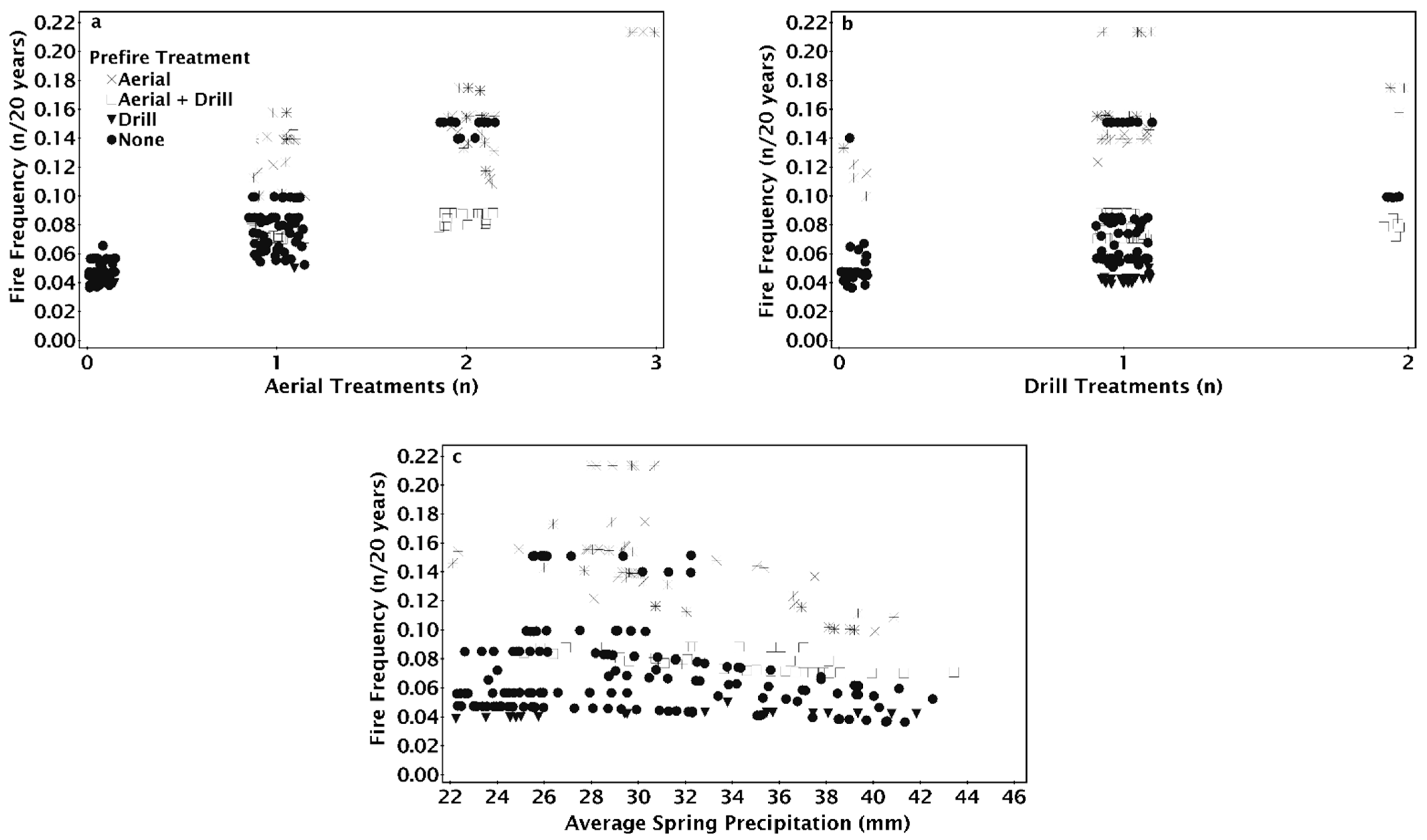
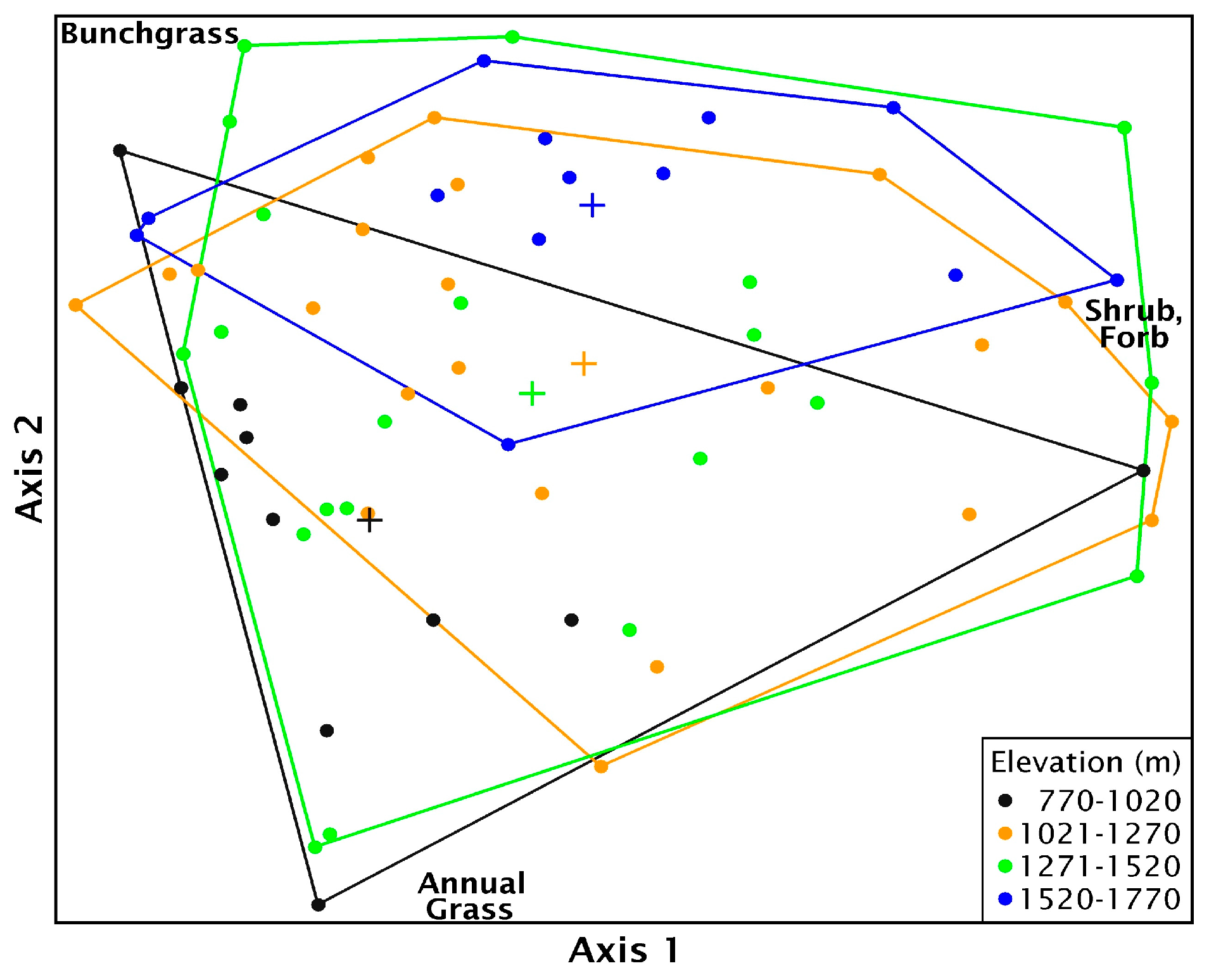
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goetz, S.J.; Mack, M.C.; Gurney, K.R.; Randerson, J.T.; Houghton, R.A. Ecosystem responses to recent climate change and fire disturbance at northern high latitudes: Observations and model results contrasting northern Eurasia and North America. Environ. Res. Lett. 2007, 2, 9. [Google Scholar] [CrossRef]
- Liu, Y.; Stanturf, J.; Goodrick, S. Trends in global wildfire potential in a changing climate. For. Ecol. Manag. 2010, 259, 685–697. [Google Scholar] [CrossRef]
- Littell, J.S.; McKenzie, D.; Peterson, D.L.; Westerling, A.L. Climate and wildfire area burned in western U.S. Ecoprovinces, 1916–2003. Ecol. Appl. 2009, 19, 1003–1021. [Google Scholar] [CrossRef] [PubMed]
- Gedalof, Z.E.; Peterson, D.L.; Mantua, N.J. Atmospheric, climatic, and ecological controls on extreme wildfire years in the northwestern United States. Ecol. Appl. 2005, 15, 154–174. [Google Scholar] [CrossRef]
- Cleetus, R.; Mulik, K. Playing with Fire: How Climate Change and Development Patterns Are Contributing to the Soaring Costs of Western Wildfires; Union of Concerned Scientists: Cambridge, MA, USA, 2014. [Google Scholar]
- Mack, R.N. Invasion of Bromus tectorum L. into Western North America: An ecological chronicle. Agro-Ecosystems 1981, 7, 145–165. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire management impacts on invasive plants in the western United States. Conserv. Biol. 2006, 20, 375–384. [Google Scholar] [CrossRef]
- Bradley, B.A.; Mustard, J.F. Characterizing the landscape dynamics of an invasive plant and risk of invasion using remote sensing. Ecol. Appl. 2006, 16, 1132–1147. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Wimberly, M.C. Direct and indirect effects of climate change on projected future fire regimes in the western United States. Sci. Total Environ. 2016, 542, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Brooks, M.L.; D’antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. BioScience 2004, 54, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Dennison, P.E.; Brewer, S.C.; Arnold, J.D.; Moritz, M.A. Large wildfire trends in the western United States, 1984–2011. Geophys. Res. Lett. 2014, 41, 2928–2933. [Google Scholar] [CrossRef]
- Fusco, E.J.; Finn, J.T.; Balch, J.K.; Nagy, R.C.; Bradley, B.A. Invasive grasses increase fire occurrence and frequency across US ecoregions. Proc. Natl. Acad. Sci. USA 2019, 116, 23594–23599. [Google Scholar] [CrossRef]
- Miller, J.D.; Skinner, C.N.; Safford, H.D.; Knapp, E.E.; Ramirez, C.M. Trends and causes of severity, size, and number of fires in northwestern California, USA. Ecol. Appl. 2011, 22, 184–203. [Google Scholar] [CrossRef]
- Hurteau, M.D.; Bradford, J.B.; Fulé, P.Z.; Taylor, A.H.; Martin, K.L. Climate change, fire management, and ecological services in the southwestern US. For. Ecol. Manag. 2014, 327, 280–289. [Google Scholar] [CrossRef]
- Barbero, R.; Abatzoglou, J.T.; Larkin, N.K.; Kolden, C.A.; Stocks, B. Climate change presents increased potential for very large fires in the contiguous United States. Int. J. Wildland Fire 2015, 24, 892–899. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Kolden, C.A. Climate change in western US deserts: Potential for increased wildfire and invasive annual grasses. Rangel. Ecol. Manag. 2011, 64, 471–478. [Google Scholar] [CrossRef]
- McKenzie, D.; Gedalof, Z.E.; Peterson, D.L.; Mote, P. Climatic change, wildfire, and conservation. Conserv. Biol. 2004, 18, 890–902. [Google Scholar] [CrossRef]
- Blaisdell, J.P.; Murray, R.B.; McArthur, E.D. Managing Intermountain Rangelands—Sagebrush-Grass Ranges; US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1982; 41p.
- West, N.E. Managing for Biodiversity of Rangelands, in Biodiversity in Agroecosystems; Collins, W.W., Qualset, C.O., Eds.; CRC Press LLC: Boca Raton, FL, USA, 1999; pp. 101–126. [Google Scholar]
- Bradley, B.A.; Curtis, C.A.; Fusco, E.J.; Abatzoglou, J.T.; Balch, J.K.; Dadashi, S.; Tuanmu, M.N. Cheatgrass (Bromus tectorum) distribution in the intermountain western United States and its relationship to fire frequency, seasonality, and ignitions. Biol. Invasions 2018, 20, 1493–1506. [Google Scholar] [CrossRef] [Green Version]
- Balch, J.K.; Bradley, B.A.; D’Antonio, C.M.; Gómez-Dans, J. Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Glob. Chang. Biol. 2013, 19, 173–183. [Google Scholar] [CrossRef]
- Baker, W.L. Is wildland fire increasing in sagebrush landscapes of the western United States? Ann. Assoc. Am. Geogr. 2013, 103, 5–19. [Google Scholar] [CrossRef]
- Baker, W.L. Fire and restoration of sagebrush ecosystems. Wildl. Soc. Bull. 2006, 34, 177–185. [Google Scholar] [CrossRef]
- Bukowski, B.E.; Baker, W.L. Historical fire regimes, reconstructed from land-survey data, led to complexity and fluctuation in sagebrush landscapes. Ecol. Appl. 2013, 23, 546–564. [Google Scholar] [CrossRef] [PubMed]
- Bradley, B.A. Regional analysis of the impacts of climate change on cheatgrass invasion shows potential risk and opportunity. Glob. Chang. Biol. 2009, 15, 196–208. [Google Scholar] [CrossRef]
- Link, S.O.; Keeler, C.W.; Hill, R.W.; Hagen, E. Bromus tectorum cover mapping and fire risk. Int. J. Wildland Fire 2006, 15, 113–119. [Google Scholar] [CrossRef]
- West, N.E.; Hassan, M.A. Recovery of sagebrush-grass vegetation following wildfire. J. Range Manag. 1985, 38, 131–134. [Google Scholar] [CrossRef]
- Peterson, E.B. Mapping Percent-Cover of the Invasive Species Bromus Tectorum (Cheatgrass) over a Large Portion of Nevada from Satellite Imagery; Nevada Natural Heritage Program: Carson City, NV, USA, 2003. [Google Scholar]
- Jessop, B.D.; Anderson, V.J. Cheatgrass invasion in salt desert shrublands: Benefits of postfire reclamation. Rangel. Ecol. Manag. 2007, 60, 235–243. [Google Scholar] [CrossRef]
- Jewell, S. Secretarial Order 3336; Department of the Interior: Washington, DC, USA, 2015; p. 5.
- BLM. Burned Area Emergency Stabilization and Rehabilitation Manual, (BLM Handbook H-1742-1); U.S. Department of Interior, Bureau of Land Management: Washington, DC, USA, 2007; 80p.
- Merriam, K.E.; Keeley, J.E.; Beyers, J.L. Fuel breaks affect nonnative species abundance In Californian plant communities. Ecol. Appl. 2006, 16, 515–527. [Google Scholar] [CrossRef]
- Ott, J.E.; Cox, R.D.; Shaw, N.L. Comparison of postfire seeding practices for Wyoming big sagebrush. Rangel. Ecol. Manag. 2017, 70, 625–632. [Google Scholar] [CrossRef]
- Rogler, G.A.; Russell, J.L. Crested wheatgrass: Early history in the United States. J. Range Manag. 1983, 36, 91–93. [Google Scholar] [CrossRef]
- Vale, T.R. Sagebrush conversion projects: An element of contemporary environmental change in the Western United States. Biol. Conserv. 1974, 6, 274–284. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Welty, J.L.; Toevs, G.R. Seventy-five years of vegetation treatments on public rangelands in the Great Basin of North America. Rangelands 2017, 39, 1–9. [Google Scholar] [CrossRef]
- Copeland, S.M.; Munson, S.M.; Pilliod, D.S.; Welty, J.L.; Bradford, J.B.; Butterfield, B.J. Long-term trends in restoration and associated land treatments in the southwestern United States. Restor. Ecol. 2018, 26, 311–322. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Welty, J.L. Land Treatment Digital Library: U.S. Geological Survey Data Series 806. 2013. Available online: http://pubs.er.usgs.gov/publication/ds806;https://ltdl.wr.usgs.gov/ (accessed on 1 March 2013).
- Asay, K.H.; Dewey, D.R.; Gomm, F.B.; Johnson, D.A.; Carlson, J.R. Registration of ‘Hycrest’ crested wheatgrass. Crop Sci. 1985, 25, 368–369. [Google Scholar] [CrossRef]
- Knutson, K.C.; Pyke, D.A.; Wirth, T.A.; Arkle, R.S.; Pilliod, D.S.; Brooks, M.L.; Chambers, J.C.; Grace, J.B. Long-term effects of seeding after wildfire on vegetation in Great Basin shrubland ecosystems. J. Appl. Ecol. 2014, 51, 1414–1424. [Google Scholar] [CrossRef]
- Clinton, W. Executive Order 13112- Invasive Species. Fed. Regist. 1999, 64, 6183–6186. [Google Scholar]
- Knick, S.T.; Rotenberry, J.T. Ghosts of habitats past: Contribution of landscape change to current habitats used by shrubland birds. Ecology 2000, 81, 220–227. [Google Scholar] [CrossRef]
- Knick, S.T.; Dobkin, D.S.; Rotenberry, J.T.; Schroeder, M.A.; Vander Haegen, W.M.; Van Riper, C., III. Teetering on the Edge or Too Late? Conservation and Research Issues for Avifauna of Sagebrush Habitats. Condor 2003, 105, 611–634. [Google Scholar] [CrossRef]
- Knick, S.T. Historical development, principal federal legislation, and current management of sagebrush habitats: Implications for conservation, in Greater Sage-Grouse: Ecology and conservation of a landscape species and its habitats. Stud. Avian Biol. 2011, 38, 13–31. [Google Scholar]
- Connelly, J.W.; Schroeder, M.A.; Sands, A.R.; Braun, C.E. Guidelines to manage sage grouse populations and their habitats. Wildl. Soc. Bull. 2000, 28, 967–985. [Google Scholar]
- BLM. Management Considerations for Sagebrush (Artemisia) in the Western United States; U.S. Department of Interior, Bureau of Land Management: Washington, DC, USA, 2002; 73p.
- Arkle, R.S.; Pilliod, D.S.; Hanser, S.E.; Brooks, M.L.; Chambers, J.C.; Grace, J.B.; Knutson, K.C.; Pyke, D.A.; Welty, J.L.; Wirth, T.A. Quantifying restoration effectiveness using multi-scale habitat models: Implications for sage-grouse in the Great Basin. Ecosphere 2014, 5, 1–32. [Google Scholar] [CrossRef]
- Hulet, A.; Boyd, C.S.; Davies, K.W.; Svejcar, T.J. Prefire (preemptive) management to decrease fire-induced bunchgrass mortality and reduce reliance on postfire seeding. Rangel. Ecol. Manag. 2015, 68, 437–444. [Google Scholar] [CrossRef]
- Bradford, J.B.; Lauenroth, W.K. Controls over invasion of Bromus tectorum: The importance of climate, soil, disturbance and seed availability. J. Veg. Sci. 2006, 17, 693–704. [Google Scholar]
- Ponzetti, J.M.; McCune, B.; Pyke, D.A. Biotic soil crusts in relation to topography, cheatgrass and fire in the Columbia Basin, Washington. Bryologist 2007, 110, 706–722. [Google Scholar] [CrossRef]
- Reisner, M.D.; Grace, J.B.; Pyke, D.A.; Doescher, P.S. Conditions favouring Bromus tectorum dominance of endangered sagebrush steppe ecosystems. J. Appl. Ecol. 2013, 50, 1039–1049. [Google Scholar] [CrossRef] [Green Version]
- Martinson, E.J.; Omi, P.N. Fuel Treatments and Fire Severity: A Meta-Analysis; Res. Pap. RMRS-RP-103WWW; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2013; 35p.
- Hudak, A.T.; Rickert, I.; Morgan, P.; Strand, E.; Lewis, S.A.; Robichaud, P.; Hoffman, C.; Holden, Z.A. Review of Fuels Treatment Effectiveness in Forests and Rangelands and a Case Study from the 2007 Megafires in Central Idaho, USA; Gen. Tech. Rep. RMRS-GTR-252; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; 60p.
- Miller, R.F.; Chambers, J.C.; Pyke, D.A.; Pierson, F.B.; Williams, C.J. A Review of Fire Effects on Vegetation and Soils in the Great Basin Region: Resposnse and Ecological Site Characteristics; Gen. Tech. Rep. RMRS-GTR-308; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2013; 126p.
- NWCG. Glossary of Wildland Fire Terminology; PMS 205; N.W.C. Group: Boise, ID, USA, 2012; p. 190.
- Strand, E.K.; Vierling, L.A.; Bunting, S.C.; Gessler, P.E. Quantifying successional rates in western aspen woodlands: Current conditions, future predictions. For. Ecol. Manag. 2009, 257, 1705–1715. [Google Scholar] [CrossRef]
- Strand, E.K.; Vierling, L.A.; Bunting, S.C. A spatially explicit model to predict future landscape composition of aspen woodlands under various management scenarios. Ecol. Model. 2009, 220, 175–191. [Google Scholar] [CrossRef]
- Sunderman, S.O.; Weisberg, P.J. Predictive modelling of burn probability and burn severity in a desert spring ecosystem. Int. J. Wildland Fire 2012, 21, 1014–1024. [Google Scholar] [CrossRef]
- Roth, A.D.; Bunting, S.C.; Strand, E.K. Relationships between landscape patterns and fire occurrence within a successional gradient in sagebrush steppe–juniper woodland. Int. J. Wildland Fire 2011, 20, 69–77. [Google Scholar] [CrossRef]
- Peeler, J.L.; Smithwick, E.A.H. Exploring invasibility with species distribution modeling: How does fire promote cheatgrass (Bromus tectorum) invasion within lower montane forests? Divers. Distrib. 2018, 24, 1308–1320. [Google Scholar] [CrossRef] [Green Version]
- Ottmar, R.D.; Sandberg, D.V.; Riccardi, C.L.; Prichard, S.J. An overview of the fuel characteristic classification system—Quantifying, classifying, and creating fuelbeds for resource planning. Can. J. For. Res. 2007, 37, 2383–2393. [Google Scholar] [CrossRef]
- Welch, B.L. Big Sagebrush: A Sea Fragmented into Lakes, Ponds, and Puddles; General Technical Report RMRS-GTR-144; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005; 210p.
- BLM. Historic Fire Perimeter; U.S. Department of the Interior (DOI), Bureau of Land Management, Idaho State Office: Boise, ID, USA, 2015. Available online: https://catalog.data.gov/dataset/fire-perimeters-historic-polygon (accessed on 1 March 2015).
- NOC. Completed Vegetation Treatments; U.S. Department of the Interior (DOI), Bureau of Land Management, Idaho State Office, GIS Staff: Denver, CO, USA, 2014. Available online: https://catalog.data.gov/dataset/vegetation-treatment-area-completed-polygonbd74a (accessed on 1 March 2015).
- BLM. Bureau of Land Management EPlanning Project Search. 2019. Available online: https://eplanning.blm.gov/ (accessed on 1 October 2019).
- Division, Geospatial. TIGER 2015 Roads; USDA Geospatial Data Gateway; U.S. Department of Commerce: Washington, DC, USA, 2015.
- Excellence, National Geospatial Center of National Hydrography Dataset; USA Natural Resources Conservation Service: Washington, DC, USA, 2012.
- BLM. BLM Idaho Surface Management Agency (Surface Ownership); Bureau of Land Management; U.S. Department of Interior, Idaho State Office: Boise, ID, USA, 2015.
- LandFire, 2014 Biophysical Settings. Available online: https://www.landfire.gov/bps.php (accessed on 1 January 2014).
- NAIP. National Agriculture Imagery Program; USA Farm Service Agency: Washington, DC, USA, 2015.
- Hilty, J. DOI-BLM-ID-T010-2012-0020-DNA (Kinyon Road Fire (G1CH) Emergency Stabilization and Burned Area Rehabilitation (ES&BAR) Plan). 2012. Available online: https://eplanning.blm.gov/epl-front-office/eplanning/projectSummary.do?methodName=renderDefaultProjectSummary&projectId=33702 (accessed on 1 June 2013).
- PRISM. PRISM Climate Group. Oregon State University. 2014. Available online: http://prism.oregonstate.edu (accessed on 1 May 2014).
- Center, E.D. National Elevation Dataset; U.S. Geological Survey; 2013. Available online: https://datagateway.nrcs.usda.gov (accessed on 1 March 2013).
- McCune, B.; Mefford, M.J. HyperNiche: Nonparametric Multiplicative Habitat Modeling; Version 2.30; MjM Software: Gleneden Beach, OR, USA, 2009. [Google Scholar]
- McCune, B. Non-parametric habitat models with automatic interactions. J. Veg. Sci. 2006, 17, 819–830. [Google Scholar] [CrossRef]
- McCune, B. Nonparametric Multiplicative Regression for Habitat Modeling. 2011. Available online: http://www.pcord.com/NPMRintro.pdf (accessed on 1 May 2016).
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Narum, S.R. Beyond Bonferroni: Less conservative analyses for conservation genetics. Conserv. Genet. 2006, 7, 783–787. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M. PC-ORD. Multivariate Analysis of Ecological Data; Version 7.02; MjM Software: Gleneden Beach, OR, USA, 2016. [Google Scholar]
- Welch, B.L.; Criddle, C. Countering Misinformation Concerning Big Sagebrush; Research Paper RMRS-RP-40; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2003; p. 28.
- Shinneman, D.J.; McIlroy, S.K. Identifying key climate and environmental factors affecting rates of post-fire big sagebrush Artemisia tridentata recovery in the northern Columbia Basin, USA. Int. J. Wildland Fire 2016, 25, 933–945. [Google Scholar] [CrossRef]
- Yoder, C.; Caldwell, M. Effects of perennial neighbors and nitrogen pulses on growth and nitrogen uptake by Bromus tectorum. Plant Ecol. 2002, 158, 77–84. [Google Scholar] [CrossRef]
- Blank, R.R.; Morgan, T. Suppression of Bromus tectorum L. by established perennial grasses: Potential mechanisms part one. Appl. Environm. Soil Sci. 2012, 2012, 9. [Google Scholar]
- Davies, K.W.; Johnson, D.D. Established perennial vegetation provides high resistance to reinvasion by exotic annual grasses. Rangel.Ecol. Manag. 2017, 70, 748–754. [Google Scholar] [CrossRef]
- Svejcar, T.; Boyd, C.; Davies, K.; Hamerlynck, E.; Svejcar, L. Challenges and limitations to native species restoration in the Great Basin, USA. Plant Ecol. 2017, 218, 81–94. [Google Scholar] [CrossRef]
- James, J.J.; Svejcar, T.J.; Rinella, M.J. Demographic processes limiting seedling recruitment in arid grassland restoration. J. Appl. Ecol. 2011, 48, 961–969. [Google Scholar] [CrossRef]
- Boyd, C.S.; Lemos, J.A. Evaluating winter/spring seeding of a native perennial bunchgrass in the sagebrush steppe. Rangel. Ecol. Manag. 2015, 68, 494–500. [Google Scholar] [CrossRef]
- Dyer, A.R.; Hardison, J.L.; Rice, K.J. Phenology constrains opportunistic growth response in Bromus tectorum L. Plant Ecol. 2012, 213, 103–112. [Google Scholar] [CrossRef]
- Compagnoni, A.; Adler, P.B. Warming, soil moisture, and loss of snow increase Bromus tectorum’s population growth rate. Elem. Sci. Anthr. 2014, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fusco, E.J.; Abatzoglou, J.T.; Balch, J.K.; Finn, J.T.; Bradley, B.A. Quantifying the human influence on fire ignition across the western USA. Ecol. Appl. 2016, 26, 2390–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, K.W.; Nafus, A.M. Exotic annual grass invasion alters fuel amounts, continuity, and moisture content. Int. J. Wildland Fire 2013, 22, 353–358. [Google Scholar] [CrossRef]
- Boyte, S.P.; Wylie, B.K.; Major, D.J. Mapping and monitoring cheatgrass dieoff in rangelands of the northern Great Basin, USA. Rangel. Ecol. Manag. 2015, 68, 18–28. [Google Scholar] [CrossRef]
- Akinsoji, A. Postfire vegetation dynamics in a sagebrush steppe in southeastern Idaho, USA. Vegetatio 1988, 78, 151–155. [Google Scholar] [CrossRef]
- Pyke, D.A.; Wirth, T.A.; Beyers, J.L. Does seeding after wildfires in rangelands reduce erosion or invasive species? Restor. Ecol. 2013, 21, 415–421. [Google Scholar] [CrossRef]
- West, N.E.; Yorks, T.P. Vegetation responses following wildfire on grazed and ungrazed sagebrush semi-desert. J. Range Manag. 2002, 55, 171–181. [Google Scholar] [CrossRef]
- Morris, L.R.; Leger, E.A. Secondary succession in the sagebrush semidesert 66 years after fire in the Great Basin, USA. Natl. Areas J. 2016, 36, 187–193. [Google Scholar] [CrossRef]
- Mangla, S.; Sheley, R.L.; James, J.J.; Radosevich, S.R. Intra and interspecific competition among invasive and native species during early stages of plant growth. Plant Ecol. 2011, 212, 531–542. [Google Scholar] [CrossRef]
- Ray-Mukherjee, J.; Jones, T.A.; Adler, P.B.; Monaco, T.A. Immature seedling growth of two North American native perennial bunchgrasses and the invasive grass Bromus tectorum. Rangel. Ecol. Manag. 2011, 64, 358–365. [Google Scholar] [CrossRef]
- Aguirre, L.; Johnson, D.A. Influence of temperature and cheatgrass competition on seedling development of two bunchgrasses. J. Range Manag. 1991, 44, 347–354. [Google Scholar] [CrossRef]
- Goergen, E.M.; Leger, E.A.; Espeland, E.K. Native perennial grasses show evolutionary response to Bromus tectorum (cheatgrass) invasion. PLoS ONE 2011, 6, e18145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.; Leger, E.A. Plastic responses of native plant root systems to the presence of an invasive annual grass. Am. J. Bot. 2015, 102, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Hull, A.C.; Klomp, G.J. Longevity of crested wheatgrass in the sagebrush-grass type in southern Idaho. J. Range Manag. 1966, 19, 5–11. [Google Scholar] [CrossRef]
- Arredondo, J.T.; Jones, T.A.; Johnson, D.A. Seedling growth of intermountain perennial and weedy annual grasses. J. Range Manag. 1998, 51, 584–589. [Google Scholar] [CrossRef] [Green Version]
- Bowman-Prideaux, C.; Newingham, B.A.; Strand, E.K. The effects of fire history, post-fire rehabilitation, and environmental factors on shrub steppe communities. In preparation.
- Hunter, M.E.; Omi, P.N.; Martinson, E.J.; Chong, G.W. Establishment of non-native plant species after wildfires: Effects of fuel treatments, abiotic and biotic factors, and post-fire grass seeding treatments. Int. J. Wildland Fire 2006, 15, 271–281. [Google Scholar] [CrossRef]
- Wirth, T.A.; Pyke, D.A. Effectiveness of Post-Fire Seeding at the Fitzner-Eberhardt Arid Land Ecology Reserve; Geological Survey Open-File Report 2011-1241; U.S. Geological Survey: Reston, VA, USA, 2011.
- Bruce, L.B.; Perryman, B.; Conley, K.; McAdoo, K. Case Study: Grazing Management on Seeded and Unseeded Post-Fire Public Rangelands. Prof. Anim. Sci. 2007, 23, 285–290. [Google Scholar] [CrossRef]
- Newhall, R.; Rasmussen, V.P.; Kitchen, B. Introducing big sagebrush into a crested wheatgrass monoculture. Natl. Resour. Environ. Issues 2011, 17, 26. Available online: http://digitalcommons.usu.edu/nrei/vol17/iss1/26 (accessed on 30 January 2012).
- Marlette, G.M.; Anderson, J.E. Seed banks and propagule dispersal in crested-wheatgrass stands. J. Appl. Ecol. 1986, 23, 161–175. [Google Scholar] [CrossRef]
- Gunnell, K.L. Seed Banks of Sagebrush Communities Seeded with Crested Wheatgrass. Master’s Thesis, Utah State University, Logan, UT, USA, 2009. Available online: http://digitalcommons.usu.edu/etd/250 (accessed on 6 April 2013).
- Pellant, M.; Lysne, C.R. Strategies to enhance plant structure and diversity in crested wheatgrass seedings. In Proceedings of the Sage-Grouse Habitat Restoration Symposium Proceedings, Boise, ID, USA, 4–7 June 2001; Shaw, N.L., Pellant, M., Monsen, S.B., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005. [Google Scholar]
- Nafus, A.M.; Svejcar, T.J.; Ganskopp, D.C.; Davies, K.W. Abundances of coplanted native bunchgrasses and crested wheatgrass after 13 Years. Rangel. Ecol. Manag. 2015, 68, 211–214. [Google Scholar] [CrossRef]
- Ganskopp, D.; Myers, B.; Lambert, S.; Cruz, R. Preferences and behavior of cattle grazing 8 varieties of grasses. J. Range Manag. 1997, 50, 578–586. [Google Scholar] [CrossRef]
- Jones, T.A.; Michael, H.R.; Dale, C.N. Cattle preference for 4 wheatgrass taxa. J. Range Manag. 1994, 47, 119–122. [Google Scholar] [CrossRef]
- Ganskopp, D.; Richman, L.; Johnson, D.; Angell, R.; Cruz, R. Preferences of Angora goats for eight selections of grasses used for reclamation of Great Basin rangelands. Small Rumin. Res. 1996, 19, 103–112. [Google Scholar] [CrossRef]
- Orrock, J.; Witter, M. Multiple drivers of apparent competition reduce re-establishment of a native plant in invaded habitats. Oikos 2010, 119, 101–108. [Google Scholar] [CrossRef]
- Roscher, C.; Gerighausen, U.; Schmid, B.; Schulze, E.D. Plant diversity and community history shift colonization success from early-to mid-successional species. J. Plant Ecol. 2015, 8, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Pilliod, D.S.; Welty, J.L.; Arkle, R.S. Refining the cheatgrass-fire cycle in the Great Basin: Precipitation timing and fine fuel composition predict wildfire trends. Ecol. Evol. 2017, 7, 8126–8151. [Google Scholar] [CrossRef] [PubMed]
- Strand, E.K.; Launchbaugh, K.L.; Limb, R.F.; Torell, L.A. Livestock grazing effects on fuel loads for wildland fire in sagebrush dominated ecosystems. J. Rangel. Appl. 2014, 1, 35–57. [Google Scholar]
- Davies, K.W.; Vavra, M.; Schultz, B.W.; Rimbey, N.R. Implications of longer term grazing rest in the sagebrush steppe. J. Rangel. Appl. 2014, 1, 14–34. [Google Scholar]
- Chambers, J.C.; Miller, R.F.; Board, D.I.; Pyke, D.A.; Roundy, B.A.; Grace, J.B.; Schupp, E.W.; Tausch, R.J. Resilience and resistance of sagebrush ecosystems: Implications for state and transition models and management treatments. Rangel. Ecol. Manag. 2014, 67, 440–454. [Google Scholar] [CrossRef] [Green Version]
- Davies, K.W.; Bates, J.D.; Svejcar, T.J.; Boyd, C.S. Effects of long-term livestock grazing on fuel characteristics in rangelands: An example from the sagebrush steppe. Rangel. Ecol. Manag. 2010, 63, 662–669. [Google Scholar] [CrossRef]
- Williams, J.R.; Morris, L.R.; Gunnell, K.L.; Johanson, J.K.; Monaco, T.A. Variation in sagebrush communities historically seeded with crested wheatgrass in the eastern Great Basin. Rangel. Ecol. Manag. 2017, 70, 683–690. [Google Scholar] [CrossRef]
- Jewell, S. An integrated rangeland fire management strategy. In Final Report to the Secretary of the Interior; Department of the Interior: Washington, DC, USA, 2015; p. 94. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fire Regime Variable | Category | Count (n) |
|---|---|---|
| Total Fire (n) | Unburned | 164 |
| 1 | 114 | |
| 2 | 154 | |
| 3 | 63 | |
| 4 | 42 | |
| 5 | 24 | |
| 6–7 | 12 | |
| Return Interval (years) | <5 | 6 |
| 5–10 | 143 | |
| 11–20 | 188 | |
| >20 | 236 | |
| Fire Frequency | 0.00 | 236 |
| 0.05 | 1880 | |
| 0.10 | 81 | |
| 0.15 | 43 | |
| ≥0.20 | 26 |
| Response Variable | Eval xR² | Model Fit xR² (±SE) | Avg Size | Predictor Variable | Sensitivity | Tolerance | |
|---|---|---|---|---|---|---|---|
| Fires (n) * | 0.8 | 0.83 | 31.2 | 1st fire year | 0.09 | 9.45 | (15%) |
| 1950–2014 | (±0.0) | Spring VPD min | 0.05 | 0.26 | (25%) | ||
| Treatment (n) | 0.05 | 1.5 | (25%) | ||||
| Pre-fire seeding treat † | - | - | - | ||||
| Fire Return Interval * | 0.78 | 0.81 | 43.32 | Aerial seeding (n) | 0.22 | 0.5 | (10%) |
| 1994–2014 | (±0.0) | 1st year treated | 0.15 | 10.35 | (15%) | ||
| Spring VPD min | 0.02 | 0.46 | (45%) | ||||
| Pre-fire seeding treat † | - | - | - | ||||
| Fire Frequency * | 0.76 | 0.77 | 33.29 | Drill seeding (n) | 0.07 | 0.75 | (25%) |
| 1994–2014 | (±0.0) | Spring precipitation | 0.05 | 6.28 | (25%) | ||
| Aerial seeding (n) | 0.03 | 0.25 | (5%) | ||||
| Pre-fire seeding treat † | - | - | - | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowman-Prideaux, C.; Newingham, B.A.; Strand, E.K. The Effect of Seeding Treatments and Climate on Fire Regimes in Wyoming Sagebrush Steppe. Fire 2021, 4, 16. https://doi.org/10.3390/fire4020016
Bowman-Prideaux C, Newingham BA, Strand EK. The Effect of Seeding Treatments and Climate on Fire Regimes in Wyoming Sagebrush Steppe. Fire. 2021; 4(2):16. https://doi.org/10.3390/fire4020016
Chicago/Turabian StyleBowman-Prideaux, Chris, Beth A. Newingham, and Eva K. Strand. 2021. "The Effect of Seeding Treatments and Climate on Fire Regimes in Wyoming Sagebrush Steppe" Fire 4, no. 2: 16. https://doi.org/10.3390/fire4020016
APA StyleBowman-Prideaux, C., Newingham, B. A., & Strand, E. K. (2021). The Effect of Seeding Treatments and Climate on Fire Regimes in Wyoming Sagebrush Steppe. Fire, 4(2), 16. https://doi.org/10.3390/fire4020016