Conversion of a Semiarid Nevada Soil to Irrigated Agriculture Preferentially Removes Labile Carbon

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Particle Size Fractionation
2.4. Density Fractionation
2.5. Soil C, N, δ13C and δ15N
2.6. Mid-Infrared Spectroscopy
2.7. Microbial Respiration
2.8. Statistical Analyses
3. Results
3.1. .Particle Size and Density Fraction Mass
3.2. Carbon and Nitrogen
3.3. Stable Isotope Composition
3.4. Mid-Infrared Spectroscopy
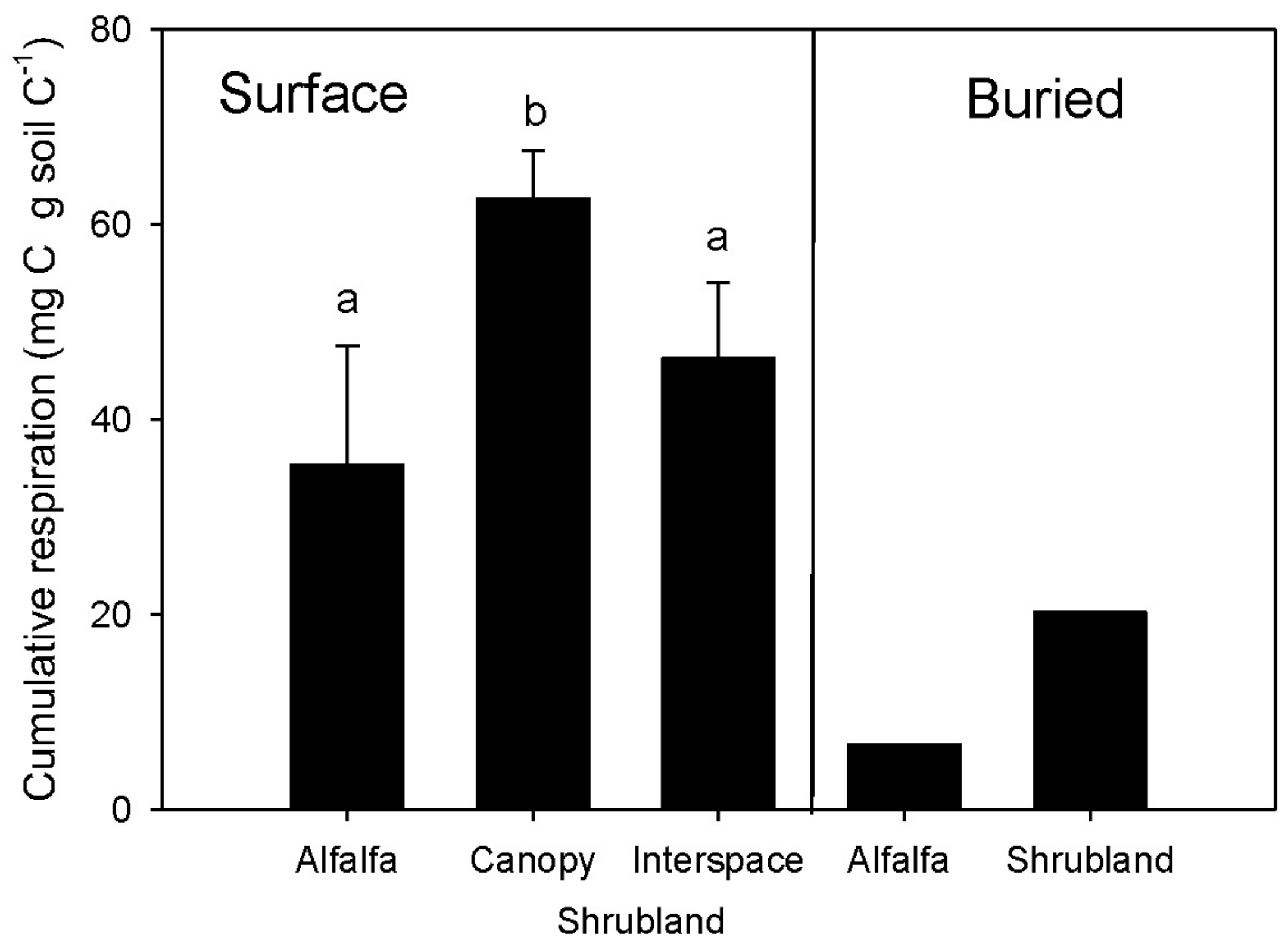
3.5. Microbial Respiration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Emmerich, W.E. Carbon dioxide fluxes in a semiarid environment with high carbonate soils. Agric. For. Meteorol. 2003, 116, 91–102. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Scharlemann, J.P.W.; Tanner, E.V.J.; Hiederer, R.; Kapos, V. Global soil carbon: Understanding and managing the largest terrestrial carbon pool. Carbon Manag. 2014, 5, 81–91. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon dynamics in cropland and rangeland. Environ. Pollut. 2002, 116, 353–362. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Wang, J. Impact of land use change on profile distributions of soil organic carbon fractions in the Yanqi Basin. Catena 2014, 115, 79–84. [Google Scholar] [CrossRef]
- Entry, J.A.; Sojka, R.E.; Shewmaker, G.E. Management of irrigated agriculture to increase organic carbon storage in soils. Soil Sci. Soc. Am. J. 2002, 66, 1957–1964. [Google Scholar] [CrossRef]
- Zach, A.; Tiessen, H.; Noellemeyer, E. Carbon turnover and carbon-13 natural abundance under land use change in semiarid savanna soils of La Pampa, Argentina. Soil Sci. Soc. Am. J. 2006, 70, 1541–1546. [Google Scholar] [CrossRef]
- Miller, R.E.; Huenneke, L.F. Size decline in Larrea tridentata (Creosotebush). Southwest Nat. 1996, 41, 248–250. [Google Scholar]
- Rundel, P.W.; Gibson, A.C. Ecological Communities and Processes in a Mojave Desert Ecosystem: Rock Valley, Nevada; Cambridge University Press: New York, NY, USA, 1996. [Google Scholar]
- Schlesinger, W.H.; Bernhardt, E.S. Biogeochemistry: An Analysis of Global Change, 3rd ed.; Academic Press: Oxford, UK, 2013. [Google Scholar]
- Lapenis, A.G.; Lawrence, G.B.; Bailey, S.W.; Aparin, B.F.; Shiklomanov, A.I.; Speranskaya, N.A.; Torn, M.S.; Calef, M. Climatically driven loss of calcium in steppe soil as a sink for atmospheric carbon. Glob. Biogeochem. Cycle 2008, 22. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, B.A.; Verburg, P.S.J. Effluxed CO2-13C from sterilized and unsterilized treatments of a calcareous soil. Soil Biol. Biochem. 2006, 38, 1727–1733. [Google Scholar] [CrossRef]
- Xie, J.; Li, Y.; Zhai, C.; Li, C.; Lan, Z. CO2 absorption by alkaline soils and its implication to the global carbon cycle. Environ. Geol. 2009, 56, 953–961. [Google Scholar] [CrossRef]
- Li, X.R.; Wang, X.P.; Li, T.; Zhang, J.G. Microbiotic soil crust and its effect on vegetation and habitat on artificially stabilized desert dunes in Tengger Desert, North China. Biol. Fertil. Soils 2002, 35, 147–154. [Google Scholar]
- Li, X.R.; Zhang, P.; Su, Y.G.; Jia, R.L. Carbon fixation by biological soil crusts following revegetation of sand dunes in arid desert regions of China: A four-year field study. Catena 2012, 97, 119–126. [Google Scholar] [CrossRef]
- Su, Y.Z.; Wang, X.F.; Yang, R.; Lee, J. Effects of sandy desertified land rehabilitation on soil carbon sequestration and aggregation in an arid region in China. J. Environ. Manag. 2010, 91, 2109–2116. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Lin, W.; Zhang, Y. Characteristics of carbon flux in two biologically crusted soils in the Gurbantunggut Desert, Northwestern China. Catena 2012, 96, 41–48. [Google Scholar] [CrossRef]
- Prǎvǎlie, R. Drylands extent and environmental issues. A global approach. Earth-Sci. Rev. 2016, 161, 259–278. [Google Scholar] [CrossRef]
- Eswaran, H.; Reich, P.F.; Kimble, J.M.; Beinroth, F.H.; Padmanabhan, E.; Moncharoen, P. Global carbon stocks. In Global Climate Change and Pedogenic Carbonates; Lal, R., Kimble, J.M., Eswaran, H., Stewart, B.A., Eds.; CRC/Lewis: Boca Raton, FL, USA, 2000. [Google Scholar]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 2014, 65, 4–21. [Google Scholar] [CrossRef]
- Jasoni, R.L.; Smith, S.D.; Arnone, J.A., III. Net ecosystem CO2 exchange in Mojave Desert shrublands during the eighth year of exposure to elevated CO2. Glob. Chang. Biol. 2005, 11, 749–756. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Fenstermaker, L.F.; Arnone, J.A., III. Large annual net ecosystem CO2 uptake of a Mojave Desert ecosystem. Glob. Chang. Biol. 2008, 14, 1475–1487. [Google Scholar] [CrossRef]
- Reid, W.V.; Mooney, H.A.; Cropper, A.; Capistrano, D.; Carpenter, S.R.; Chopra, K.; Dasgupta, P.; Dietz, T.; Duraiappah, A.K.; Hassan, R.; et al. Ecosystems and Human Well-Being: Synthesis, Millenium Ecosystem Assessment; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Wu, L.; Wood, Y.; Jiang, P.; Li, L.; Pan, G.; Lu, J.; Chang, A.C.; Enloe, H.A. Carbon sequestration and dynamics of two irrigated agricultural soils in California. Soil Sci. Soc. Am. J. 2008, 72, 808–814. [Google Scholar] [CrossRef]
- Albaladejo, J.; Ortiz, R.; Garcia-Franco, N.; Navarro, A.R.; Almagro, M.; Pintado, J.G.; Martínez-Mena, M. Land use and climate change impacts on soil organic carbon stocks in semi-arid Spain. J. Soils Sediments 2013, 13, 265–277. [Google Scholar] [CrossRef]
- Cochran, R.L.; Collins, H.P.; Kennedy, A.; Bezdicek, D.F. Soil carbon pools and fluxes after land conversion in a semiarid shrub-steppe ecosystem. Biol. Fertil. Soils 2007, 43, 479–489. [Google Scholar] [CrossRef]
- Tiessen, H.; Feller, C.; Sampaio, E.V.S.B.; Garin, P. Carbon sequestration and turnover in semiarid savannas and dry forest. Clim. Chang. 1998. [Google Scholar] [CrossRef]
- Urioste, A.M.; Hevia, G.G.; Hepper, E.N.; Anton, L.E.; Bono, A.A.; Buschiazzo, D.E. Cultivation effects on the distribution of organic carbon, total nitrogen and phosphorus in soils of the semiarid region of Argentinian Pampas. Geoderma 2006, 136, 621–630. [Google Scholar] [CrossRef]
- Wu, H.; Guo, Z.; Gao, Q.; Peng, C. Distribution of soil inorganic carbon storage and its changes due to agricultural land use activity in China. Agric. Ecosyst. Environ. 2009, 129, 413–421. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2015, 6, 166–172. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Smith, D.M.S.; Lambin, E.F.; Turner, B.L., II; Mortimore, M.; Batterbury, S.P.J.; Downing, T.E.; Dowlatabadi, H.; Fernández, R.J.; Herrick, J.E.; et al. Global desertification: building a science for dryland development. Science 2007, 316, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Galbally, I.; Meyer, C.P.; Wang, Y.-P.; Kirstine, W. Soil-atmosphere exchange of CH4, CO, N2O and NOx and the effects of land-use change in the semiarid Mallee system in Southeastern Australia. Glob. Chang. Biol. 2010, 16, 2407–2419. [Google Scholar]
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Lemenih, M.; Itanna, F. Soil carbon stocks and turnovers in various vegetation types and arable lands along an elevation gradient in southern Ethiopia. Geoderma 2004, 123, 177–188. [Google Scholar] [CrossRef]
- Saha, D.; Kukal, S.S.; Sharma, S. Landuse impacts on SOC fractions and aggregate stability in typic ustochrepts of Northwest India. Plant Soil 2011, 339, 457–470. [Google Scholar] [CrossRef]
- Bonino, E.E. Changes in carbon pools associated with a land-use gradient in the Dry Chaco, Argentina. For. Ecol. Manag. 2006, 223, 183–189. [Google Scholar] [CrossRef]
- Qiu, L.; Wei, X.; Zhang, X.; Cheng, J.; Gale, W.; Guo, C.; Long, T. Soil organic carbon losses due to land use change in a semiarid grassland. Plant Soil 2012, 355, 299–309. [Google Scholar] [CrossRef]
- Zhao, W.Z.; Xiao, H.L.; Liu, Z.M.; Li, J. Soil degradation and restoration as affected by land use change in the semiarid Bashang area, northern China. Catena 2005, 59, 173–186. [Google Scholar] [CrossRef]
- Zhou, Z.; Sun, O.J.; Huang, J.; Li, L.; Liu, P.; Han, X. Soil carbon and nitrogen stores and storage potential as affected by land-use in an agro-pastoral ecotone of northern China. Biogeochemistry 2007, 82, 127–138. [Google Scholar] [CrossRef]
- Cates, A.M.; Ruark, M.D.; Hedtcke, J.L.; Posner, J.L. Long-term tillage, rotation and perennialization effects on particulate and aggregate soil organic matter. Soil Tillage Res. 2016, 155, 371–380. [Google Scholar] [CrossRef]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and land-use change: processes and potential. Glob. Chang. Biol. 2000, 6, 317–328. [Google Scholar] [CrossRef]
- Scanlon, B.R.; Reedy, R.C.; Bronson, K.F. Impacts of land use change on nitrogen cycling archived in semiarid unsaturated zone nitrate profiles, Southern High Plains, Texas. Environ. Sci. Technol. 2008, 42, 7566–7572. [Google Scholar] [CrossRef] [PubMed]
- Smith, P. Land use change and soil organic carbon dynamics. Nutr. Cycl. Agroecosyst. 2008, 81, 169–178. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Belnap, J.; Marion, G. On carbon sequestration in desert ecosystems. Glob. Chang. Biol. 2009, 15, 1488–1490. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Wu, H.; Guo, Z.; Peng, C. Land use induced changes of organic carbon storage in soils of China. Glob. Chang. Biol. 2003, 9, 305–315. [Google Scholar] [CrossRef]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Gillabel, J.; Denef, K.; Brenner, J.; Merckx, R.; Paustian, K. Carbon sequestration and soil aggregation in center-pivot irrigated and dryland cultivated farming systems. Soil Sci. Soc. Am. J. 2007, 71, 1020–1028. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Zhang, Z.; Wan, S. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob. Chang. Biol. 2009, 15, 184–195. [Google Scholar] [CrossRef]
- Stockmann, U.; Adams, M.A.; Crawford, J.W.; Field, D.J.; Henakaarchchi, N.; Jenkins, M.; Minasny, B.; McBratney, A.B.; de Courcelles, V.; Singh, K.; et al. The knowns, known unknowns and unknowns of sequestration of soil organic carbon. Agric. Ecosyst. Environ. 2013, 164, 80–99. [Google Scholar] [CrossRef]
- Yan, M.; Zhou, G.; Zhang, X. Effects of irrigation on the soil CO2 efflux from different poplar clone plantations in arid northwest China. Plant Soil 2013, 375, 89–97. [Google Scholar] [CrossRef]
- Austin, A.T.; Yahdjian, L.; Stark, J.M.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.A.; Schaeffer, S.M. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Hastings, S.J.; Oechel, W.C.; Muhlia-Melo, A. Diurnal, seasonal and annual variation in the net ecosystem CO2 exchange of a desert shrub community (Sarcocaulescent) in Baja California, Mexico. Glob. Chang. Biol. 2005, 11, 927–939. [Google Scholar] [CrossRef]
- Huxman, T.E.; Snyder, K.A.; Tissue, D.; Leffler, A.J.; Ogle, K.; Pockman, W.T.; Sandquist, D.R.; Potts, D.L.; Schwinning, S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystem. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Burke, I.C.; Reiners, W.A.; Schimel, D.S. Organic matter turnover in a sagebrush steppe landscape. Biogeochemistry 1989, 7, 11–31. [Google Scholar] [CrossRef]
- Schaeffer, S.M.; Evans, R.D. Pulse additions of soil carbon and nitrogen affect soil nitrogen dynamics in an arid Colorado Plateau shrubland. Oecologia 2005, 145, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Sponseller, R.A. Precipitation pulses and soil CO2 flux in a Sonoran Desert ecosystem. Glob. Chang. Biol. 2007, 13, 426–436. [Google Scholar] [CrossRef]
- DeMarco, J.; Filley, T.; Throop, H.L. Patterns of woody plant-derived soil carbon losses and persistence after brush management in a semi-arid grassland. Plant Soil 2016, 406, 277–293. [Google Scholar] [CrossRef]
- Hooker, T.D.; Stark, J.M. Carbon flow from plant detritus and soil organic matter to microbes-Linking carbon and nitrogen cycling in semiarid soils. Soil Sci. Soc. Am. J. 2012, 76, 903–914. [Google Scholar] [CrossRef]
- Throop, H.L.; Lajtha, K.; Kramer, M. Density fractionation and 13C reveal changes in soil carbon following woody encroachment in a desert ecosystem. Biogeochemistry 2013, 112, 409–422. [Google Scholar] [CrossRef]
- Plant Guide for Yellow-Flowered Alfalfa (Medicago sativa subsp. falcata (L.) Arcang. Available online: https://www.nrcs.usda.gov/Internet/FSE_PLANTMATERIALS/publications/ndpmcpg12773.pdf (accessed on 3 August 2018).
- Plant Guide for Rubber Rabbitbrush (Ericameria nauseosa). Available online: https://plants.usda.gov/factsheet/pdf/fs_erna10.pdf (accessed on 3 August 2018).
- Plant Guide for Yellow Rabbitbrush (Chrysothamnus viscidiflorus). Available online: https://plants.usda.gov/plantguide/pdf/pg_chvi8.pdf (accessed on 3 August 2018).
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-NRCS: Washington, DC, USA, 2014.
- Kettler, T.A.; Doran, J.W.; Gilbert, T.L. Simplified method for soil particle-size determination to accompany soil-quality analyses. Soil Sci. Soc. Am. J. 2001, 65, 849–852. [Google Scholar] [CrossRef]
- Sollins, P.; Glassman, C.; Paul, E.A.; Swanston, C.; Lajtha, K.; Heil, J.W.; Elliott, E.T. Soil carbon and nitrogen: Pools and fractions. In Standard Soil Methods for Long-Term Ecological Research; Robertson, G.P., Coleman, D.C., Bledsoe, C.S., Sollins, P., Eds.; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Harris, D.; Horwáth, W.R.; van Kessel, C. Acid fumigation of soils to remove carbonates prior to total organic carbon or carbon-13 isotopic analysis. Soil Sci. Soc. Am. J. 2001, 65, 1853–1856. [Google Scholar] [CrossRef]
- Werner, R.A.; Bruch, B.A.; Brand, W.A. ConFlo III—An interface for high precision δ13C and δ15N analysis with an extended dynamic range. Rapid Commun. Mass Spectrom. 1999, 13, 1237–1241. [Google Scholar] [CrossRef]
- Parikh, S.J.; Goyne, K.W.; Margenot, A.J.; Mukome, F.N.; Calderón, F.J. Soil chemical insights provided through vibrational spectroscopy. Adv. Agron. 2014, 126, 1–148. [Google Scholar]
- Haberhauer, G.; Feigl, B.; Gerzabek, M.H.; Cerri, C. FT-IR spectroscopy of organic matter in tropical soils: Changes induced through deforestation. J. Appl. Spectrosc. 2000, 54, 221–224. [Google Scholar] [CrossRef]
- Calderón, F.J.; Reeves, J., III; Collins, H.P.; Paul, E.A. Chemical differences in soil organic matter fractions determined by diffuse-reflectance mid-infrared spectroscopy. Soil Sci. Soc. Am. J. 2011, 75, 568–579. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Ding, G.; Novak, J.M.; Amarasiriwardena, D.; Hunt, P.G.; Xing, B. Soil organic matter characteristics as affected by tillage management. Soil Sci. Soc. Am. J. 2002, 66, 421–429. [Google Scholar] [CrossRef]
- Haberhauer, G.; Gerzabek, M.H. Drift and transmission FT-IR spectroscopy of forest soils: An approach to determine decomposition processes of forest litter. Vib. Spectrosc. 1999, 19, 413–417. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Wang, T. Fourier transform Raman spectroscopic characterization of humic substances. Vib. Spectrosc. 1997, 14, 105–112. [Google Scholar] [CrossRef]
- Matamala, R.; Jastrow, J.; Hofmann, S.; Mishra, U.; Michaelson, G.; Calderon, F.; Ping, C.-L.; Zhaosheng, F. Influence of site and soil properties on the DRIFT spectra of northern cold-region soils. Geoderma 2017, 305, 80–91. [Google Scholar] [CrossRef]
- Matejkova, S.; Simon, T. Application of FTIR spectroscopy for evaluation of hydrophobic/hydrophilic organic components in arable soil. Plant Soil Environ. 2012, 58, 192–195. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K.; Elliott, E.T.; Combrink, C. Soil structure and organic matter: I. Distribution of aggregate-size classes and aggregate-associated carbon. Soil Sci. Soc. Am. J. 2000, 64, 681–689. [Google Scholar] [CrossRef]
- Calderón, F.J.; Jackson, L.E. Rototillage, disking, and subsequent irrigation: effects on soil nitrogen dynamics, microbial biomass, and carbon dioxide efflux. J. Environ. Qual. 2002, 31, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Oades, J.M. The retention of organic matter in soils. Biogeochemistry 1988, 5, 35–70. [Google Scholar] [CrossRef]
- De Oliveira, S.P.; De Lacerda, N.B.; Blum, S.C.; Escobar, M.E.O.; De Oliveira, T.S. Organic carbon and nitrogen stocks in soils of northeastern Brazil converted to irrigated agriculture. Land Degrad. Dev. 2015, 26, 9–21. [Google Scholar] [CrossRef]
- Kocyigit, R.; Demirci, S. Long-term changes of aggregate-associated and labile soil organic carbon and nitrogen after conversion from forest to grassland and cropland in northern Turkey. Land Degrad. Dev. 2012, 23, 475–482. [Google Scholar] [CrossRef]
- Fallahzade, J.; Hajabbasi, M.A. The effects of irrigation and cultivation on the quality of desert soil in central Iran. Land Degrad. Dev. 2012, 23, 53–61. [Google Scholar] [CrossRef]
- Amundson, R.; Austin, A.T.; Schuur, E.A.G.; Yoo, K.; Matzek, V.; Kendall, C.; Uebersax, A.; Brenner, D.; Baisden, W.T. Global patterns of the isotopic composition of soil and plant nitrogen. Glob. Biogeochem. Cycle 2003, 17, 1031. [Google Scholar] [CrossRef]
- Ariz, I.; Cruz, C.; Neves, T.; Irigoyen, J.J.; Garcia-Olaverri, C.; Nogués, S.; Aparicio-Tejo, P.M.; Aranjuelo, I. Leaf δ15N as a physiological indicator of the responsiveness of N2-fixing alfalfa plants to elevated [CO2], temperature and low water availability. Front. Plant Sci. 2015, 6, 574. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, P.; Ishizu, A.; Doucett, R.; Hart, S.C.; Schwartz, E.; Menyailo, O.V.; Hungate, B.A. 13C and 15N natural abundance of the soil microbial biomass. Soil Biol. Biochem. 2006, 38, 3257–3266. [Google Scholar] [CrossRef]
- Högberg, P.; Johannisson, C. 15N abundance of forests is correlated with losses of nitrogen. Plant Soil. 1993, 157, 147–150. [Google Scholar] [CrossRef]
- Kramer, M.G.; Sollins, P.; Sletten, R.S.; Swart, P.K. N isotope fractionation and measures of organic matter alteration during decomposition. Ecology 2003, 84, 2021–2025. [Google Scholar] [CrossRef]
- Werth, M.; Kuzyakov, Y. 13C fractionation at the root-microorganisms-soil interface: A review and outlook for partitioning studies. Soil Biol. Biochem. 2010, 42, 1372–1384. [Google Scholar] [CrossRef]
- Kozlovskii, F.I.; Ruhlmann, J.; Travnikova, L.S.; Kuzyakov, Y.V. Clay differentiation in initially homogeneous substrates upon long-term field experiments. Eurasian Soil Sci. 2001, 34, 130–138. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surface | Buried | ||||
|---|---|---|---|---|---|
| Alfalfa | Shrub Canopy | Shrub Interspace | Alfalfa | Shrub | |
| Sand + POM | 11.8 (0.2) a | 14.6 (0.3) b | 14.8 (0.3) b | 10.3 (0.9) | 17.5 (0.7) |
| Silt | 9.4 (0.1) a | 11.4 (0.2) a | 11.3 (0.1) a | 14.9 (2.6) | 15.5 (1.1) |
| Clay | 8.4 (0.2) a | 10.9 (0.7) b | 11.0 (0.4) b | 13.0 (0.1) | 14.8 (0.3) |
| Free floating | 14.0 (0.3) a | 16.9 (0.9) b | 16.2 (0.3) ab | 19.9 (0.9) | 26.7 (2.2) |
| Heavy | 9.8 (0.2) a | 11.1 (0.4) a | 11.1 (0.5) a | 12.9 (1.8) | 13.5 (0.2) |
| Aboveground | 8.7 (0.5) a | 65.5 (1.7) b | 69.4 (3.4) b | ||
| Belowground | 13.4 (0.3) a | 41.7 (1.1) b | 47.7 (1.9) c | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trimble, B.R.; Calderon, F.J.; Poulson, S.R.; Verburg, P.S.J. Conversion of a Semiarid Nevada Soil to Irrigated Agriculture Preferentially Removes Labile Carbon. Soil Syst. 2018, 2, 38. https://doi.org/10.3390/soilsystems2030038
Trimble BR, Calderon FJ, Poulson SR, Verburg PSJ. Conversion of a Semiarid Nevada Soil to Irrigated Agriculture Preferentially Removes Labile Carbon. Soil Systems. 2018; 2(3):38. https://doi.org/10.3390/soilsystems2030038
Chicago/Turabian StyleTrimble, Brittany R., Francisco J. Calderon, Simon R. Poulson, and Paul S. J. Verburg. 2018. "Conversion of a Semiarid Nevada Soil to Irrigated Agriculture Preferentially Removes Labile Carbon" Soil Systems 2, no. 3: 38. https://doi.org/10.3390/soilsystems2030038
APA StyleTrimble, B. R., Calderon, F. J., Poulson, S. R., & Verburg, P. S. J. (2018). Conversion of a Semiarid Nevada Soil to Irrigated Agriculture Preferentially Removes Labile Carbon. Soil Systems, 2(3), 38. https://doi.org/10.3390/soilsystems2030038