The Ability of Soil Pore Network Metrics to Predict Redox Dynamics Is Scale Dependent

 , ,
, , 
Abstract
:1. Introduction
- (a)
- Oxygen diffusion within aggregate domains can be estimated based on porosity alone;
- (b)
- there is a single critical oxygen concentration at which heterotrophic respiration (the major energy yielding process in soil) ceases in all organisms; and
- (c)
- oxygen consumption is constant throughout “aerobic” aggregate domains.
- (a)
- constrain the size of the soil volume that is “seen” by the tip of a platinum probe; and
- (b)
- find quantitative, numerical indices of soil structure that can be used to test assumptions about causality regarding soil structure—redox state relationships.
- (1).
- Parameterization of soil structure using computed tomography. Diffusive domains and the surrounding spatial void pattern within a given soil volume (i.e., soil structure) are considered as quantifiable through X-ray computed tomography (XCT) (XCT, [29]). Nimmo and Perkins [30] hypothesized that as a pore network was increasingly disturbed, macroporosity would decrease. We assumed that, by manipulating saturation level and manipulating the geometry of the pore network while measuring concomitant changes in electromotive potential in multiple microenvironments, relationships between the XCT quantified pore network and the unique redox state contained within could be determined. In doing so, we aimed to contribute to the development of parameters, procedures, and concepts for the application of XCT to the investigation of structure—functionality relations in soil systems [31];
- (2).
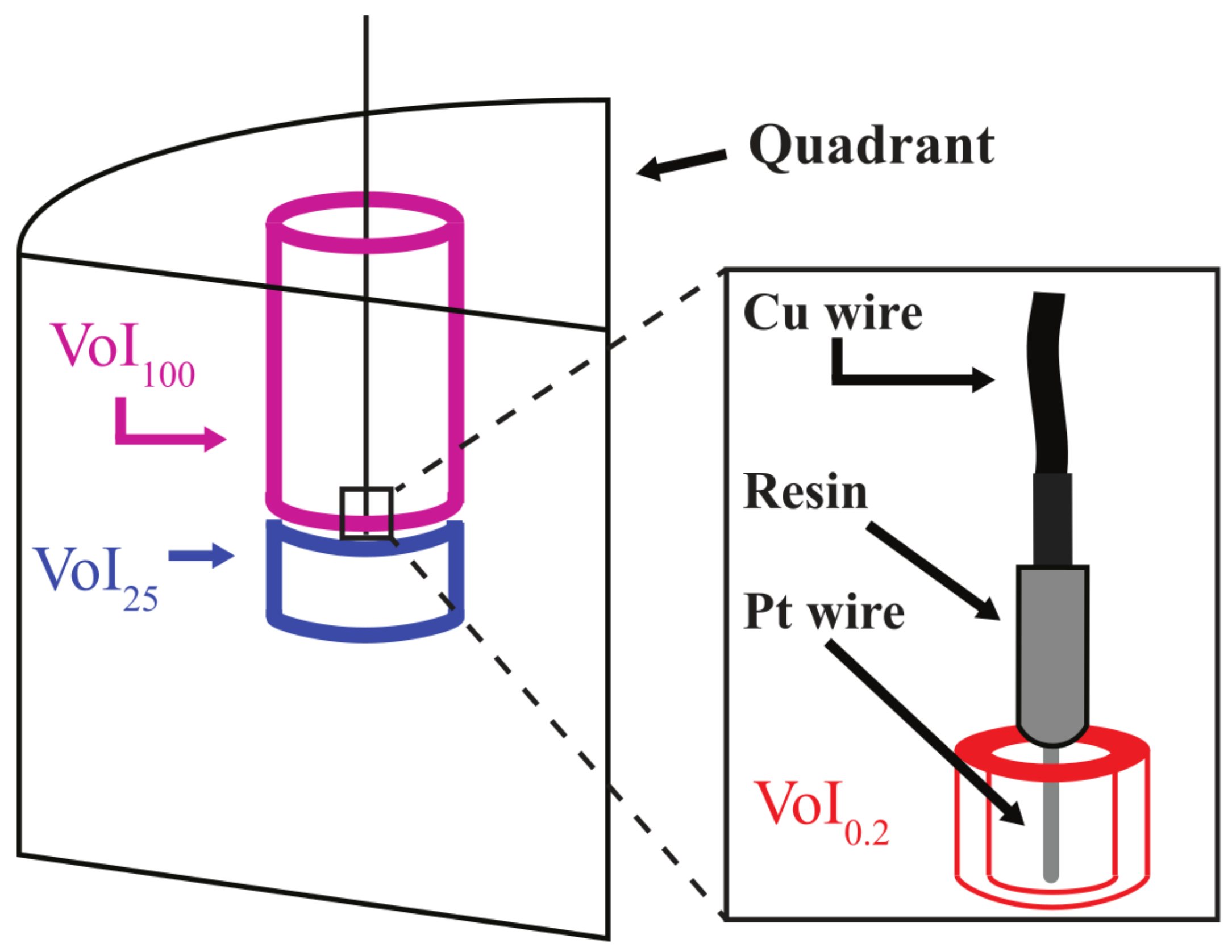
- variation of electromotive potentials in soil microenvironments. Our decision to use Pt-electrode potentials for the identification of biogeochemically distinct soil microsites was based on previous reports that Pt-electrodes are probing the redox state of very small individual volumes in the order of a few cubic millimeters [32,33,34]. To address uncertainties regarding the soil volume “seen” by the Pt-electrode tip, the relationships between virtual (i.e., defined by the settings of the analytical software) sub-sections of the pore network (Volumes of Interest, VoI) and measured electromotive potentials were examined; and
- (3).
- variation in moisture content. To elucidate the relationship(s) between wetting and drying events and the formation of anaerobic conditions we focus on short-term time brackets where moisture conditions change how the resulting variations in redox state are predicted by XCT derived pore network metrics.
2. Materials and Methods
2.1. Experimental Approach
2.1.1. Soil Description and Sample Collection
2.1.2. Set Up and Instrumentation
2.1.3. Experimental Conditions
2.2. Pore Network Quantification Using X-ray Computed Tomography
2.2.1. XCT Theory and Scan Conditions
2.2.2. Image Pre-Processing
- (a)
- The available energy sensed by the Pt-electrode tip represents the state of the soil solution in the pore system connecting the soil surface and the electrode tip. The resulting Volume of Interest (VoI100) was of a cylindrical shape centered around the electrode with a height of approximately 8 cm (minor variations between individual cylinders), a diameter of 4 cm, and an average volume of 100 mL.
- (b)
- The potential sensed represents a more constrained, but still sizable, region right below the electrode tip. This VoI had a diameter of 4 cm and extended 2 cm down from the bottom of the probe tip, resulting in a volume of approximately 25 mL (VoI25).
- (c)
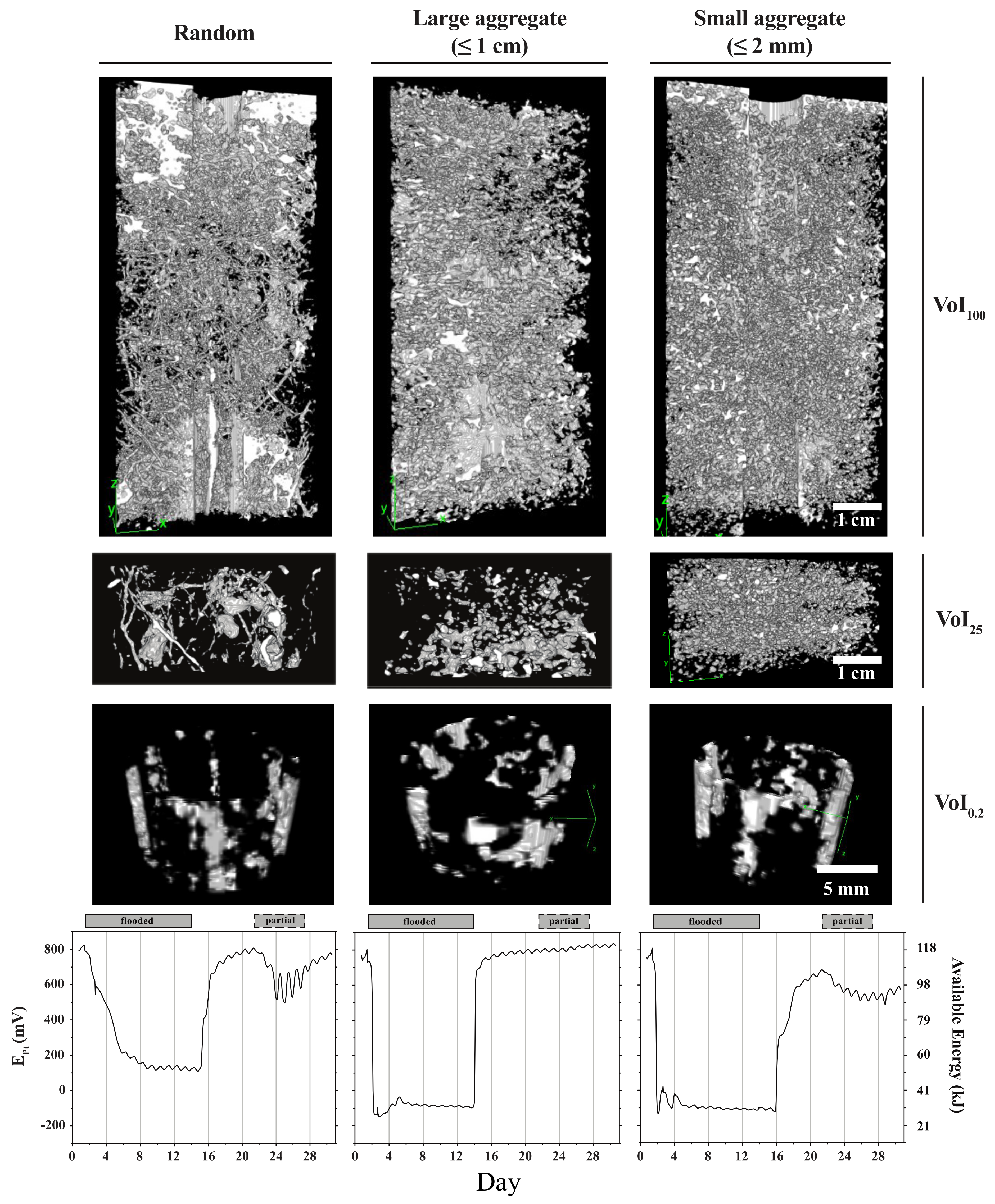
- Testing the suggestion of Fiedler [33] that Pt-electrodes are only sensitive to the conditions in a space of few cubic mm immediately surrounding and connected to the platinum tip, we finally selected a volume of interest surrounding the platinum wire in the fashion of a cylindrical sleeve with a height of 7 mm, an inner diameter of 5 mm, and a wall thickness of 0.84 mm, yielding a volume of 190 mm3 or approximately 0.2 mL (VoI0.2). The dimensions of the inner core were chosen to avoid image artifacts created by the metal of the probe tip. Figure 4 demonstrates how the respective images varied as a function of pore network structure. Representative curves are added to reiterate significant differences in available energy dynamics. For each sub-sampled VoI, the contrast was set using Fiji’s auto brightness/contrast setting. The binary threshold was then set manually by comparing pore edges in four different images to the same pore edges in the corresponding images from the 8-bit image stack prior to thresholding [45]. A 3D median filter of the dimensions, 5 × 5 × 5 pixels, was then applied to each binary stack, which reduced noise, but preserved pore edges [46].
2.2.3. Image Analyses
2.3. Statistics
3. Results and Discussion
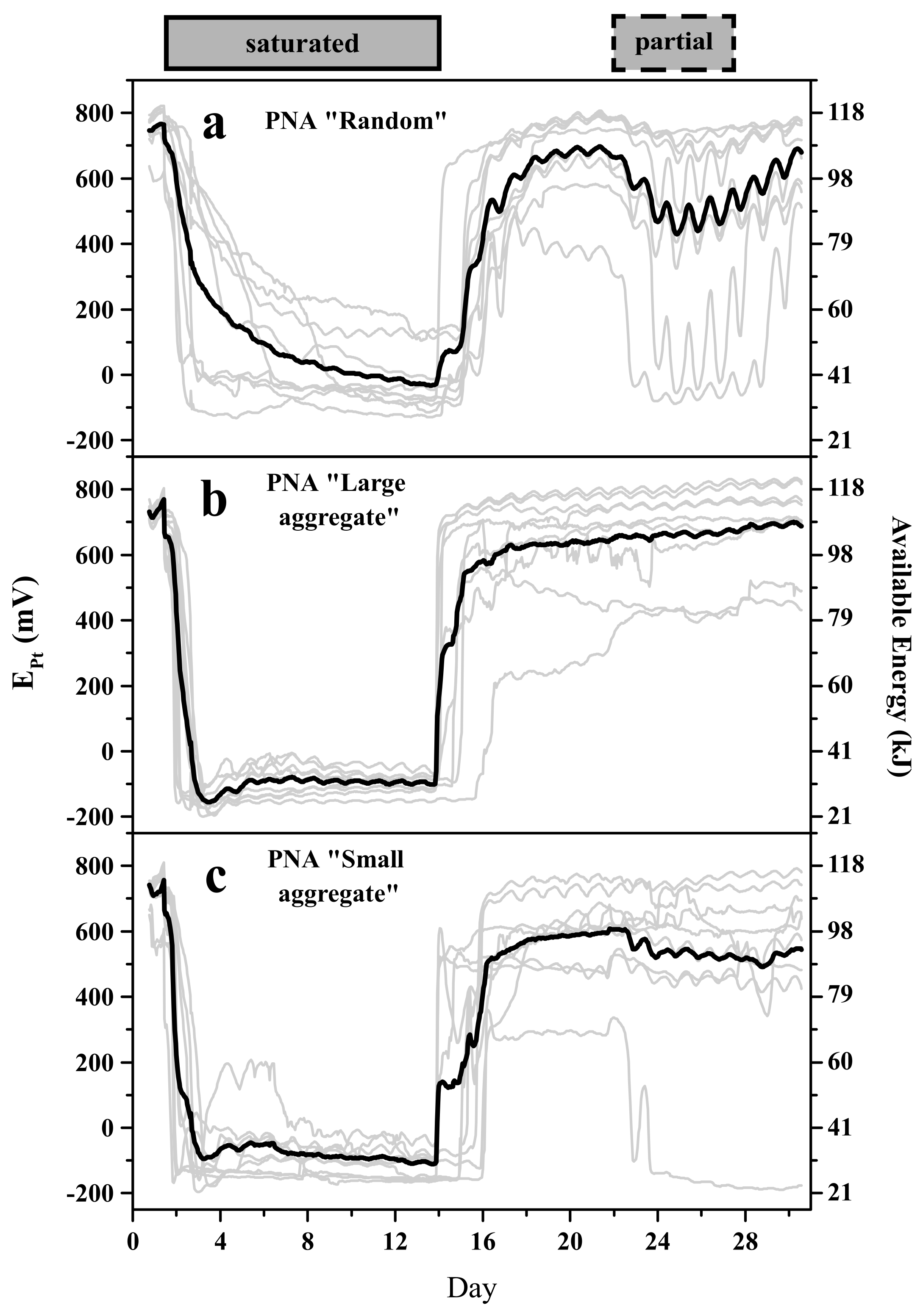
3.1. Pt-Electrodes Provide Robust and Reliable Information about Available Energy
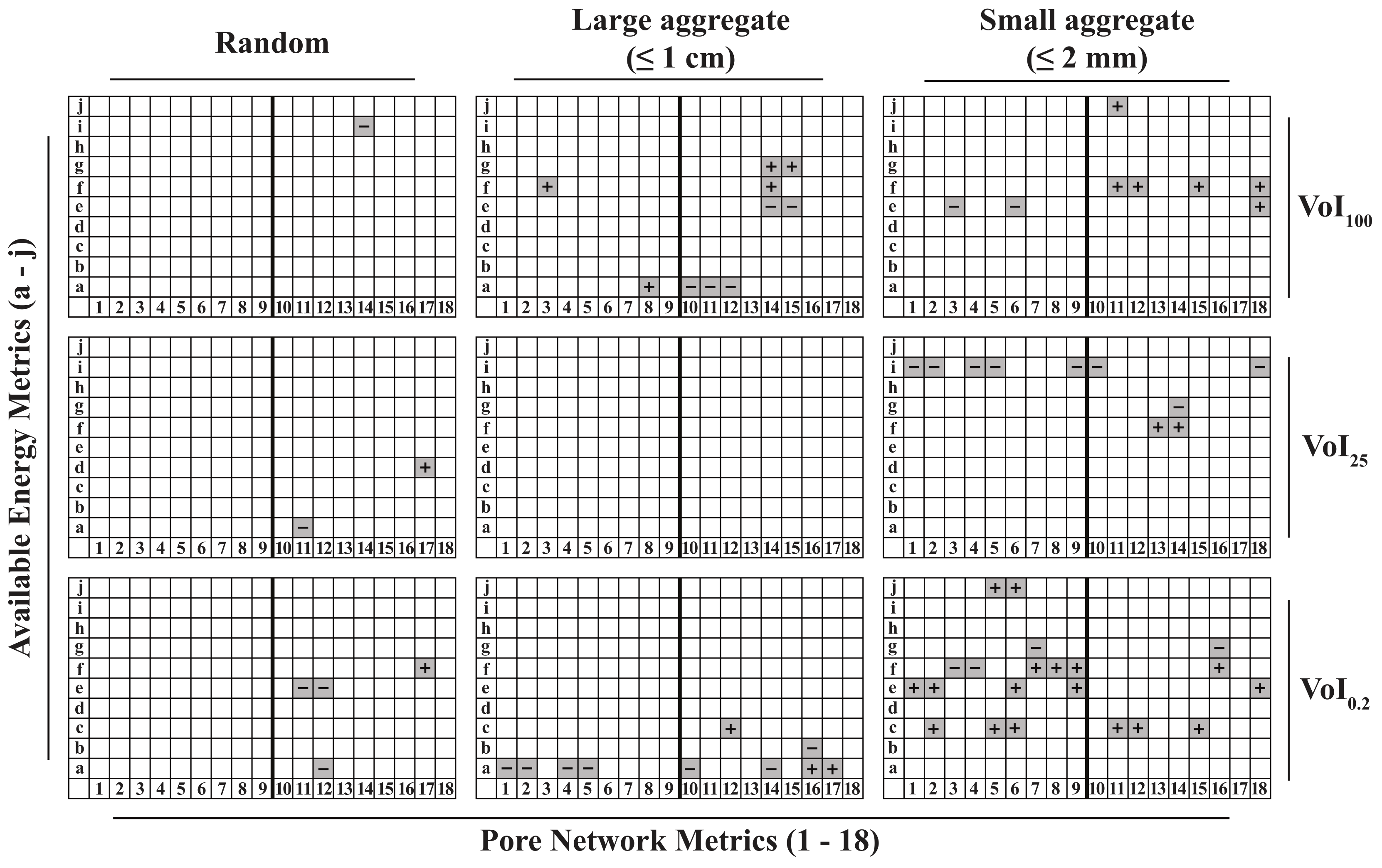
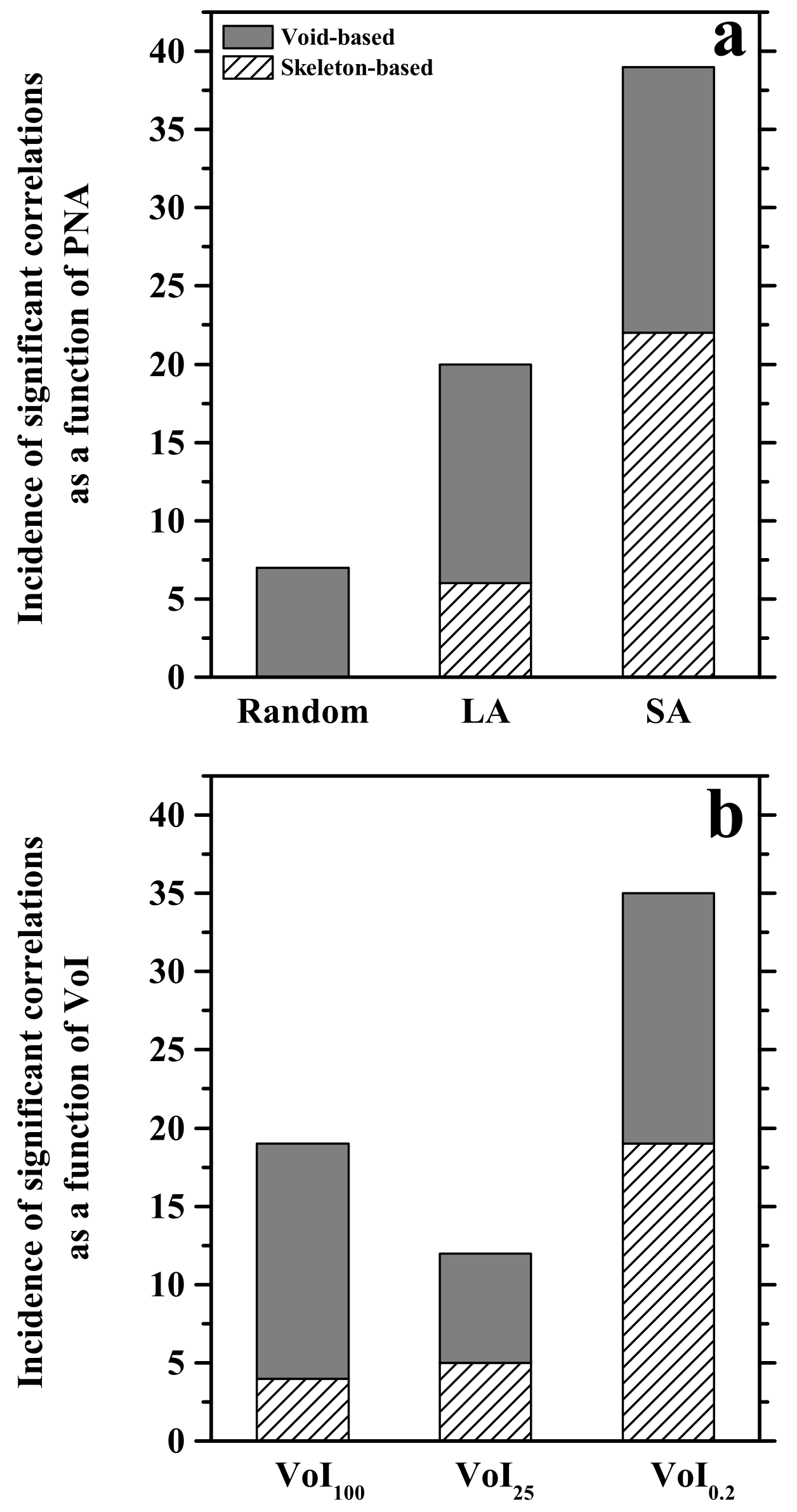
3.2. The Pore Network Metric—Pore Network Architecture Relationship Depends on the Observed Soil Volume
3.3. Pore Network Architecture Modifies Available Energy
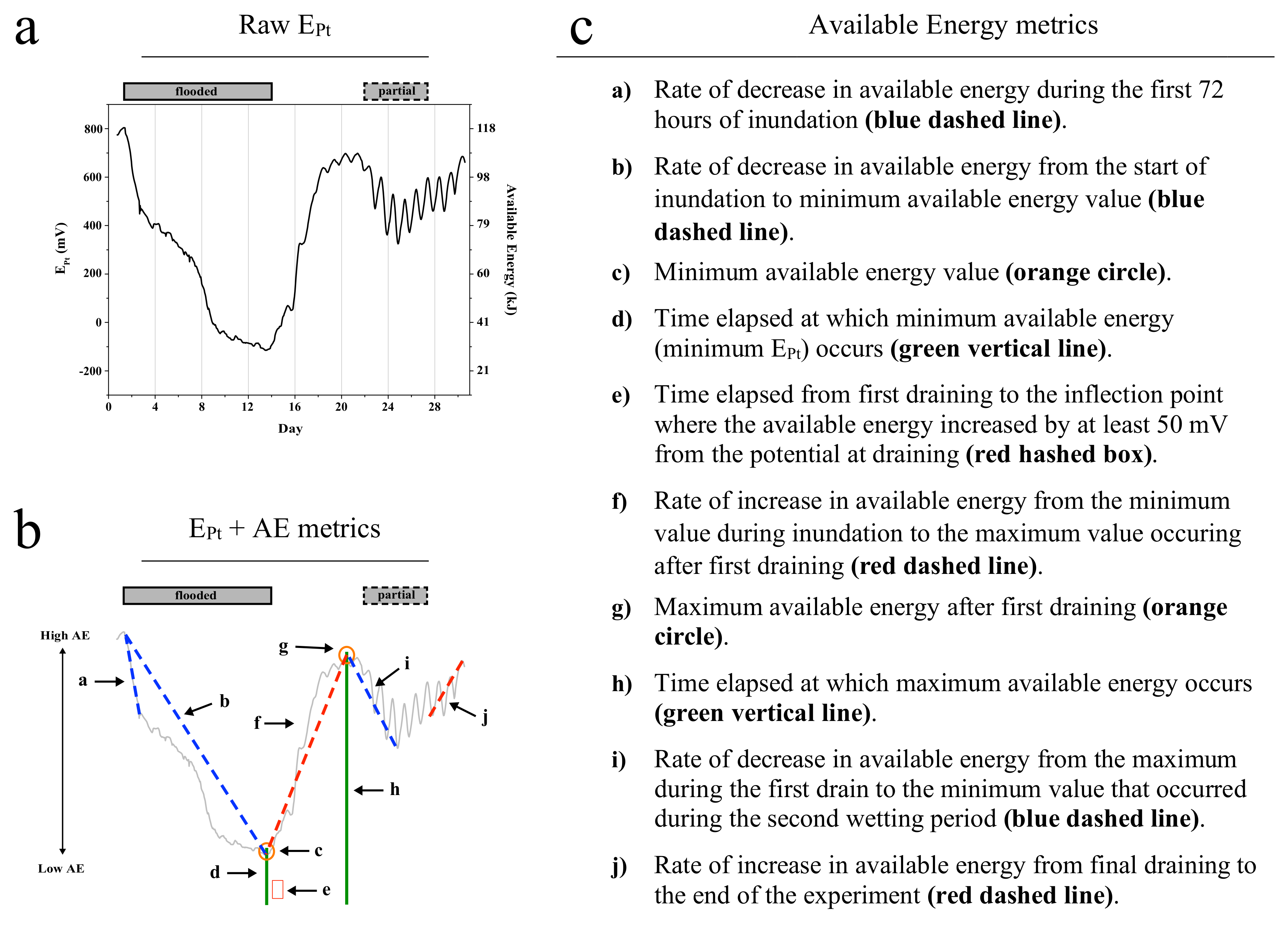
3.4. Pore Network Metrics Have Differential Power to Explain Available Energy Metrics
3.5. The Explanatory Power of PNMs Depends on Pore Network Architecture
3.6. Utility of Available Energy and Pore Network Metrics
3.7. The Explanatory Power of PNMs Is Greatest for a Small Soil Volume Immediately Surrounding the Electrode Tip
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wania, R.; Ross, I.; Prentice, I.C. Integrating peatlands and permafrost into a dynamic global vegetation model: 2. Evaluation and sensitivity of vegetation and carbon cycle processes. Glob. Biogeochem. Cycles 2009, 23. [Google Scholar] [CrossRef] [Green Version]
- Fendorf, S.; Michael, H.A.; van Geen, A. Spatial and temporal variations of groundwater arsenic in South and Southeast Asia. Science 2010, 328, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Borja, J.; Taleon, D.M.; Auresenia, J.; Gallardo, S. Polychlorinated biphenyls and their biodegradation. Process Biochem. 2005, 40, 1999–2013. [Google Scholar] [CrossRef]
- Abramowicz, D. Aerobic and Anaerobic Biodegredation of PCBs: A Review. Crit. Rev. Biotechnol. 1990, 10, 241–251. [Google Scholar] [CrossRef]
- Riley, W.J.; Maggi, F.; Kleber, M.; Torn, M.S.; Tang, J.Y.; Dwivedi, D.; Guerry, N. Long residence times of rapidly decomposable soil organic matter: Application of a multi-phase, multi-component, and vertically resolved model (BAMS1) to soil carbon dynamics. Geosci. Model Dev. 2014, 7, 1335–1355. [Google Scholar] [CrossRef]
- Cussler, E.L. Diffusion: Mass Transfer in Fluid Systems, 2nd ed.; Cambridge University Press: New York, NY, USA, 1997. [Google Scholar]
- Keiluweit, M.; Nico, P.S.; Kleber, M.; Fendorf, S. Are oxygen limitations under recognized regulators of organic carbon turnover in upland soils? Biogeochemistry 2016, 127, 157–171. [Google Scholar] [CrossRef]
- McClain, M.E.; Boyer, E.W.; Dent, C.L.; Gergel, S.E.; Grimm, N.B.; Groffman, P.M.; Hart, S.C.; Harvey, J.W.; Johnston, C.A.; Mayorga, E.; et al. Biogeochemical Hot Spots and Hot Moments at the Interface of Terrestrial and Aquatic Ecosystems. Ecosystems 2003, 6, 301–312. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Riley, W.J.; Subin, Z.M.; Lawrence, D.M.; Swenson, S.C.; Torn, M.S.; Meng, L.; Mahowald, N.M.; Hess, P. Barriers to predicting changes in global terrestrial methane fluxes: Analyses using CLM4Me, a methane biogeochemistry model integrated in CESM. Biogeosciences 2011, 8, 1925–1953. [Google Scholar] [CrossRef]
- Kuka, K.; Franko, U.; Rühlmann, J. Modelling the impact of pore space distribution on carbon turnover. Ecol. Model. 2007, 208, 295–306. [Google Scholar] [CrossRef]
- Davidson, E.A.; Samanta, S.; Caramori, S.S.; Savage, K. The Dual Arrhenius and Michaelis-Menten kinetics model for decomposition of soil organic matter at hourly to seasonal time scales. Glob. Chang. Biol. 2012, 18, 371–384. [Google Scholar] [CrossRef]
- Koven, C.D.; Riley, W.J.; Subin, Z.M.; Tang, J.Y.; Torn, M.S.; Collins, W.D.; Bonan, G.B.; Lawrence, D.M.; Swenson, S.C. The effect of vertically-resolved soil biogeochemistry and alternate soil C and N models on C dynamics of CLM4. Biogeosci. Discuss. 2013, 10, 7201–7256. [Google Scholar] [CrossRef]
- Keiluweit, M.; Wanzek, T.; Kleber, M.; Nico, P.S.; Fendorf, S. Anaerobic Microsites have an Unaccounted Role in Soil Carbon Stabilization. Nat. Commun. 2017, 8, 1771. [Google Scholar] [CrossRef] [PubMed]
- Negassa, W.C.; Guber, A.K.; Kravchenko, A.N.; Marsh, T.L.; Hildebrandt, B.; Rivers, M.L. Properties of soil pore space regulate pathways of plant residue decomposition and community structure of associated bacteria. PLoS ONE 2015, 10, e0123999. [Google Scholar] [CrossRef]
- Kravchenko, A.N.; Negassa, W.C.; Guber, A.K.; Hildebrandt, B.; Marsh, T.L.; Rivers, M.L. Intra-aggregate Pore Structure Influences Phylogenetic Composition of Bacterial Community in Macroaggregates. Soil Sci. Soc. Am. J. 2014, 78, 1924. [Google Scholar] [CrossRef]
- Ruamps, L.S.; Nunan, N.; Chenu, C. Microbial biogeography at the soil pore scale. Soil Biol. Biochem. 2011, 43, 280–286. [Google Scholar] [CrossRef]
- Wieder, W.R.; Bonan, G.B.; Allison, S.D. Global soil carbon projections are improved by modelling microbial processes. Nat. Clim. Chang. 2013, 3, 909–912. [Google Scholar] [CrossRef]
- Wieder, W.R.; Allison, S.D.; Davidson, E.A.; Georgiou, K.; Hararuk, O.; He, Y.J.; Hopkins, F.; Luo, Y.Q.; Smith, M.J.; Sulman, B.; et al. Explicitly representing soil microbial processes in Earth system models. Glob. Biogeochem. Cycles 2015, 29, 1782–1800. [Google Scholar] [CrossRef] [Green Version]
- Ruamps, L.S.; Nunan, N.; Pouteau, V.; Leloup, J.; Raynaud, X.; Roy, V.; Chenu, C. Regulation of soil organic C mineralisation at the pore scale. FEMS Microbiol. Ecol. 2013, 86, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Angle, J.C.; Morin, T.H.; Solden, L.M.; Narrowe, A.B.; Smith, G.J.; Borton, M.A.; Rey-Sanchez, C.; Daly, R.A.; Mirfenderesgi, G.; Hoyt, D.W.; et al. Methanogenesis in oxygenated soils is a substantial fraction of wetland methane emissions. Nat. Commun. 2017, 8, 1567. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimi, A.; Or, D. Dynamics of soil biogeochemical gas emissions shaped by remolded aggregate sizes and carbon configurations under hydration cycles. Glob. Chang. Biol. 2018, 24, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Baas Becking, L.G.; Kaplan, I.R.; Moore, D. Limits of the Natural Environment in terms of pH and Oxidation-Reduction potentials. J. Geol. 1960, 68, 243–288. [Google Scholar] [CrossRef]
- Wanzek, T.; Keiluweit, M.; Baham, J.; Dragila, M.I.; Fendorf, S.; Fiedler, S.; Nico, P.S.; Kleber, M. Quantifying biogeochemical heterogeneity in soil systems. Geoderma 2018, 324, 89–97. [Google Scholar] [CrossRef]
- Bohn, H.L. Redox Potentials. Soil Sci. 1971, 112, 39–45. [Google Scholar] [CrossRef]
- Trumbore, S.E. Potential responses of soil organic carbon to global environmental change. Proc. Natl. Acad. Sci. USA 1997, 94, 8284–8291. [Google Scholar] [CrossRef] [Green Version]
- Hockaday, W.C.; Masiello, C.A.; Randerson, J.T.; Smernik, R.J.; Baldock, J.A.; Chadwick, O.A.; Harden, J.W. Measurement of soil carbon oxidation state and oxidative ratio by13C nuclear magnetic resonance. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef] [Green Version]
- Dincer, I. The role of exergy in energy policy making. Energy Policy 2002, 137–149. [Google Scholar] [CrossRef]
- Luo, L.; Lin, H.; Halleck, P. Quantifying Soil Structure and Preferential Flow in Intact Soil Using X-ray Computed Tomography. Soil Sci. Soc. Am. J. 2008, 72, 1058. [Google Scholar] [CrossRef]
- Nimmo, J.R.; Perkins, K.S. Effect of soil disturbance on recharging fluxes: Case study on the Snake River Plain, Idaho National Laboratory, USA. Hydrogeol. J. 2008, 16, 829–844. [Google Scholar] [CrossRef]
- Rabot, E.; Wiesmeier, M.; Schlüter, S.; Vogel, H.J. Soil structure as an indicator of soil functions: A review. Geoderma 2018, 314, 122–137. [Google Scholar] [CrossRef]
- Cogger, C.G.; Kennedy, P.E.; Carlson, D. Seasonally Saturated Soils in the Puget Lowland II. Measuring and Interpreting Redox Potentials. Soil Sci. 1992, 154, 50–58. [Google Scholar] [CrossRef]
- Fiedler, S. In-situ longterm measurements of redox potential in hydromorphic soils, in Redox Fundamentals. In Redox Fundamentals, Processes and Measuring Techniques; Schüring, J., Schulz, H.D., Fischer, W.R., Böttcher, J., Duijnisveld, W.H.M., Eds.; Springer: New York, NY, USA, 1999; pp. 81–94. [Google Scholar]
- Fiedler, S.; Vepraskas, M.J.; Richardson, J.L. Soil Redox Potential: Importance, Field Measurements, and Observations. Adv. Agron. 2007, 94, 1–54. [Google Scholar] [CrossRef]
- Staff, S.S. Kellogg Soil Survey Laboratory Methods Manual; Soil Survey Investigations Report No. 42; Burt, R., Staff, S.S., Eds.; U.S. Department of Agriculture, Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Jones, R.H. Oxidation-Reduction Potential Measurement. ISA J. 1966, 13, 40. [Google Scholar]
- Austin, W.E.; Huddleston, J.H. Viability of permanently installed platinum redox electrodes. Soil Sci. Soc. Am. J. 1999, 63, 1757–1762. [Google Scholar] [CrossRef]
- Nordstrom, D.K.; Wilde, F.D. Field Measurements: Section 6.5 Reduction Oxidation Potential (Electrode Method). In National Field Manual for the Collection of Water Quality Data, 1.2 ed.; U.S. Geological Survey Office: Reston, VA, USA, 2005; Chapter A6. [Google Scholar]
- Taina, I.A.; Heck, R.J.; Elliot, T.R. Application of X-ray computed tomography to soil science: A literature review. Can. J. Soil Sci. 2008, 88, 1–20. [Google Scholar] [CrossRef]
- Tippkötter, R.; Eickhorst, T.; Taubner, H.; Gredner, B.; Rademaker, G. Detection of soil water in macropores of undisturbed soil using microfocus X-ray tube computerized tomography (μCT). Soil Tillage Res. 2009, 105, 12–20. [Google Scholar] [CrossRef]
- Pierret, A.; Capowiez, Y.; Belzunces, L.; Moran, C.J. 3D reconstruction and quantification of macropores using X-ray computed tomography and image analysis. Geoderma 2002, 106, 247–271. [Google Scholar] [CrossRef]
- Perret, J.; Prasher, S.O.; Kantzas, A.; Langford, C. Three-Dimensional Quantification of Macropore Networks in Undisturbed Soil Cores. Soil Sci. Soc. Am. J. 1999, 63, 1530. [Google Scholar] [CrossRef]
- Wildenschild, D.; Hopmans, J.W.; Vaz, C.M.P.; Rivers, M.L.; Rikard, D.; Christensen, B.S.B. Using X-ray computed tomography in hydrology: Systems, resolutions, and limitations. J. Hydrol. 2002, 267, 285–297. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Quin, P.R.; Cowie, A.L.; Flavel, R.J.; Keen, B.P.; Macdonald, L.M.; Morris, S.G.; Singh, B.P.; Young, I.M.; Van Zwieten, L. Oil mallee biochar improves soil structural properties—A study with x-ray micro-CT. Agric. Ecosyst. Environ. 2014, 191, 142–149. [Google Scholar] [CrossRef]
- Jassogne, L.; McNeill, A.; Chittleborough, D. 3D-visualization and analysis of macro- and meso-porosity of the upper horizons of a sodic, texture-contrast soil. Eur. J. Soil Sci. 2007, 58, 589–598. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.C.; Kashyap, R.L.; Chu, C.N. Building Skeleton Models Via 3-D Medial Surface Axis Thinning Algorithms. Cvgip Graph. Models Image Process. 1994, 56, 462–478. [Google Scholar] [CrossRef]
- Arganda-Carreras, I.; Fernandez-Gonzalez, R.; Munoz-Barrutia, A.; Ortiz-De-Solorzano, C. 3D reconstruction of histological sections: Application to mammary gland tissue. Microsc. Res. Tech. 2010, 73, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Doube, M.; Klosowski, M.M.; Arganda-Carreras, I.; Cordelieres, F.P.; Dougherty, R.P.; Jackson, J.S.; Schmid, B.; Hutchinson, J.R.; Shefelbine, S.J. BoneJ: Free and extensible bone image analysis in ImageJ. Bone 2010, 47, 1076–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.; Lin, H.; Li, S. Quantification of 3-D soil macropore networks in different soil types and land uses using computed tomography. J. Hydrol. 2010, 393, 53–64. [Google Scholar] [CrossRef]
- Lorensen, W.E.; Cline, H.E. Marching Cubes: A High Resolution 3D Surface Construction Algorithm. Comput. Graph. 1987, 21, 163–169. [Google Scholar]
- RStudio, T. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Bartlett, R.J. Characterizing soil redox behavior. In Soil Physical Chemistry; Sparks, D.L., Ed.; CRC Press: Boca Raton, FL, USA, 1998; pp. 371–397. [Google Scholar]
- Bartlett, R.J.; James, B.R. System for Categorizing Soil Redox Status by Chemical Field Testing. Geoderma 1995, 68, 211–218. [Google Scholar] [CrossRef]
- James, B.R.; Brose, D.A. Oxidation Reduciont Phenomena. In Handbook of Soil Sciences: Properties and Processes, 2nd ed.; Huang, Y.L., Malcolm, E.S., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 1–24. [Google Scholar]
- McKeague, J.A. Relationship od water table and Eh to Properties of three clay soils in the Ottawa Valley. Can. J. Soil Sci. 1965, 45, 49–62. [Google Scholar] [CrossRef]
- Köhne, J.M.; Schlüter, S.; Vogel, H.-J. Predicting Solute Transport in Structured Soil Using Pore Network Models. Vadose Zone J. 2011, 10, 1082. [Google Scholar] [CrossRef]
- Quispel, A. Measurement of the oxidation reduction potentials of normal and inundated soils. Soil Sci. 1947, 101, 265–275. [Google Scholar] [CrossRef]
- Grable, A.R.; Siemer, E.G. Effects of bulk density, aggregate size, and soil water suction on oxygen diffusiion, redox potentials, and elongatoin of corn roots. Soil Sci. Soc. Am. J. 1968, 32, 180–186. [Google Scholar] [CrossRef]
- Vepraskas, M.J.; Wilding, L.P. Aquic Moisture Regimes in Soils with and without Low Chroma Colors. Soil Sci. Soc. Am. J. 1983, 47, 280–285. [Google Scholar] [CrossRef]
- Zimmermann, I.; Fleige, H.; Horn, R. Soil structure amelioration with quicklime and irrigation experiments in earth graves. J. Soils Sediments 2016, 16, 2514–2522. [Google Scholar] [CrossRef]
- Or, D.; Smets, B.F.; Wraith, J.M.; Dechesne, A.; Friedman, S.P. Physical constraints affecting bacterial habitats and activity in unsaturated porous media—A review. Adv. Water Resour. 2007, 30, 1505–1527. [Google Scholar] [CrossRef]
- Luo, L.; Lin, H.; Schmidt, J. Quantitative Relationships between Soil Macropore Characteristics and Preferential Flow and Transport. Soil Sci. Soc. Am. J. 2010, 74, 1929. [Google Scholar] [CrossRef]
- Paradelo, M.; Katuwal, S.; Moldrup, P.; Norgaard, T.; Herath, L.; de Jonge, L.W. X-ray CT-Derived Soil Characteristics Explain Varying Air, Water, and Solute Transport Properties across a Loamy Field. Vadose Zone J. 2016, 15. [Google Scholar] [CrossRef]
- Rappoldt, C.; Crawford, J.W. The distribution of anoxic volume in a fractal model of soil. Geoderma 1999, 88, 329–347. [Google Scholar] [CrossRef]
- Naveed, M.; Moldrup, P.; Arthur, E.; Wildenschild, D.; Eden, M.; Lamandé, M.; Vogel, H.-J.; de Jonge, L.W. Revealing Soil Structure and Functional Macroporosity along a Clay Gradient Using X-ray Computed Tomography. Soil Sci. Soc. Am. J. 2013, 77, 403. [Google Scholar] [CrossRef]
- Katuwal, S.; Norgaard, T.; Moldrup, P.; Lamandé, M.; Wildenschild, D.; de Jonge, L.W. Linking air and water transport in intact soils to macropore characteristics inferred from X-ray computed tomography. Geoderma 2015, 237–238, 9–20. [Google Scholar] [CrossRef]
- Helliwell, J.R.; Miller, A.J.; Whalley, W.R.; Mooney, S.J.; Sturrock, C.J. Quantifying the impact of microbes on soil structural development and behaviour in wet soils. Soil Biol. Biochem. 2014, 74, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Kravchenko, A.N.; Guber, A.K. Soil pores and their contributions to soil carbon processes. Geoderma 2017, 287, 31–39. [Google Scholar] [CrossRef]
- Kravchenko, A.N.; Negassa, W.C.; Guber, A.K.; Rivers, M.L. Protection of soil carbon within macro-aggregates depends on intra-aggregate pore characteristics. Sci. Rep. 2015, 5, 16261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsbo, M.; Koestel, J.; Kätterer, T.; Jarvis, N. Preferential Transport in Macropores is Reduced by Soil Organic Carbon. Vadose Zone J. 2016, 15. [Google Scholar] [CrossRef] [Green Version]
- Rabbi, S.M.; Daniel, H.; Lockwood, P.V.; Macdonald, C.; Pereg, L.; Tighe, M.; Wilson, B.R.; Young, I.M. Physical soil architectural traits are functionally linked to carbon decomposition and bacterial diversity. Sci. Rep. 2016, 6, 33012. [Google Scholar] [CrossRef] [Green Version]
- Toosi, E.R.; Kravchenko, A.N.; Mao, J.; Quigley, M.Y.; Rivers, M.L. Effects of management and pore characteristics on organic matter composition of macroaggregates: Evidence from characterization of organic matter and imaging. Eur. J. Soil Sci. 2017, 68, 200–211. [Google Scholar] [CrossRef]
- Peth, S.; Horn, R.; Beckmann, F.; Donath, T.; Fischer, J.; Smucker, A.J.M. Three-Dimensional Quantification of Intra-Aggregate Pore-Space Features using Synchrotron-Radiation-Based Microtomography. Soil Sci. Soc. Am. J. 2008, 72, 897. [Google Scholar] [CrossRef]
- Sammartino, S.; Lissy, A.-S.; Bogner, C.; Van Den Bogaert, R.; Capowiez, Y.; Ruy, S.; Cornu, S. Identifying the Functional Macropore Network Related to Preferential Flow in Structured Soils. Vadose Zone J. 2015, 14. [Google Scholar] [CrossRef]
- Li, T.C.; Shao, M.A.; Jia, Y.H. Application of X-ray tomography to quantify macropore characteristics of loess soil under two perennial plants. Eur. J. Soil Sci. 2016, 67, 266–275. [Google Scholar] [CrossRef] [Green Version]
- Naveed, M.; Moldrup, P.; Schaap, M.G.; Tuller, M.; Kulkarni, R.; Vogel, H.-J.; Wollesen de Jonge, L. Prediction of biopore- and matrix-dominated flow from X-ray CT-derived macropore network characteristics. Hydrol. Earth Syst. Sci. 2016, 20, 4017–4030. [Google Scholar] [CrossRef]
- Jarvis, N.; Larsbo, M.; Koestel, J. Connectivity and percolation of structural pore networks in a cultivated silt loam soil quantified by X-ray tomography. Geoderma 2017, 287, 71–79. [Google Scholar] [CrossRef]
- Dong, H.; Blunt, M.J. Pore-network extraction from micro-computerized-tomography images. Phys. Rev. E Stat. Nonlinear Softw. Matter Phys. 2009, 80, 036307. [Google Scholar] [CrossRef] [PubMed]
- Arganda-Carreras, I. ImageJ Software Plug-in, Skeletonize, 3D 1.0.1, 2014.
- Katuwal, S.; Arthur, E.; Tuller, M.; Moldrup, P.; de Jonge, L.W. Quantification of Soil Pore Network Complexity with X-ray Computed Tomography and Gas Transport Measurements. Soil Sci. Soc. Am. J. 2015, 79, 1577. [Google Scholar] [CrossRef]
- Larsbo, M.; Koestel, J.; Jarvis, N. Relations between macropore network characteristics and the degree of preferential solute transport. Hydrol. Earth Syst. Sci. 2014, 18, 5255–5269. [Google Scholar] [CrossRef]
- Rabot, E.; Lacoste, M.; Hénault, C.; Cousin, I. Using X-ray Computed Tomography to Describe the Dynamics of Nitrous Oxide Emissions during Soil Drying. Vadose Zone J. 2015, 14. [Google Scholar] [CrossRef] [Green Version]
- Moldrup, P.; Olesen, T.; Komatsu, T.; Schønning, P.; Rolston, D.E. Tortuosity, Diffusivity, and Permeability in the Soil Liquid and Gaseous Phases. Soil Sci. Soc. Am. J. 2001, 65, 613–623. [Google Scholar] [CrossRef]
- Kawamoto, K.; Moldrup, P.; Schjønning, P.; Iversen, B.V.; Komatsu, T.; Rolston, D.E. Gas Transport Parameters in the Vadose Zone: Development and Tests of Power-Law Models for Air Permeability. Vadose Zone J. 2006, 5, 1205. [Google Scholar] [CrossRef]
- Tuli, A.; Hopmans, J.W. Effect of degree of fluid saturation on transport coefficients in disturbed soils. Eur. J. Soil Sci. 2004, 55, 147–164. [Google Scholar] [CrossRef]
- Tracy, S.R.; Daly, K.R.; Sturrock, C.J.; Crout, N.M.J.; Mooney, S.J.; Roose, T. Three-dimensional quantification of soil hydraulic properties using X-ray Computed Tomography and image-based modeling. Water Resour. Res. 2015, 51, 1006–1022. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, J.; Guggenberger, G.; Baumgartl, T.; Ellerbrock, R.H.; Urbanek, E.; Goebel, M.-O.; Kaiser, K.; Horn, R.; Fischer, W.R. Physical carbon-sequestration mechanisms under special consideration of soil wettability. J. Plant Nutr. Soil Sci. 2008, 171, 14–26. [Google Scholar] [CrossRef]
- San José Martínez, F.; Muñoz Ortega, F.J.; Caniego Monreal, F.J.; Kravchenko, A.N.; Wang, W. Soil aggregate geometry: Measurements and morphology. Geoderma 2015, 237–238, 36–48. [Google Scholar] [CrossRef]
- Ananyeva, K.; Wang, W.; Smucker, A.J.M.; Rivers, M.L.; Kravchenko, A.N. Can intra-aggregate pore structures affect the aggregate’s effectiveness in protecting carbon? Soil Biol. Biochem. 2013, 57, 868–875. [Google Scholar] [CrossRef]
- Almquist, V.; Brueck, C.; Clarke, S.; Wanzek, T.; Dragila, M.I. Bioavailable water in coarse soils: A fractal approach. Geoderma 2018, 323, 146–155. [Google Scholar] [CrossRef]
- Massol-Deya, A.A.; Whallon, J.; Hickey, R.F.; Tiedje, J.M. Channel structures in aerobic biofilms of fixed-film reactors treating contaminated groundwater. Appl. Environ. Microbiol. 1995, 61, 769–777. [Google Scholar] [PubMed]
- Vandevivere, P.; Baveye, P.C. Saturated hydraulic conductivity reduction caused by aerobic bacteria in sand columns. Soil Sci. Soc. Am. J. 1992, 56, 1–12. [Google Scholar] [CrossRef]
- Or, D.; Tuller, M. Flow in unsaturated fractured porous media: Hydraulic conductivity of rough surfaces. Water Resour. Res. 2000, 36, 1165–1177. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
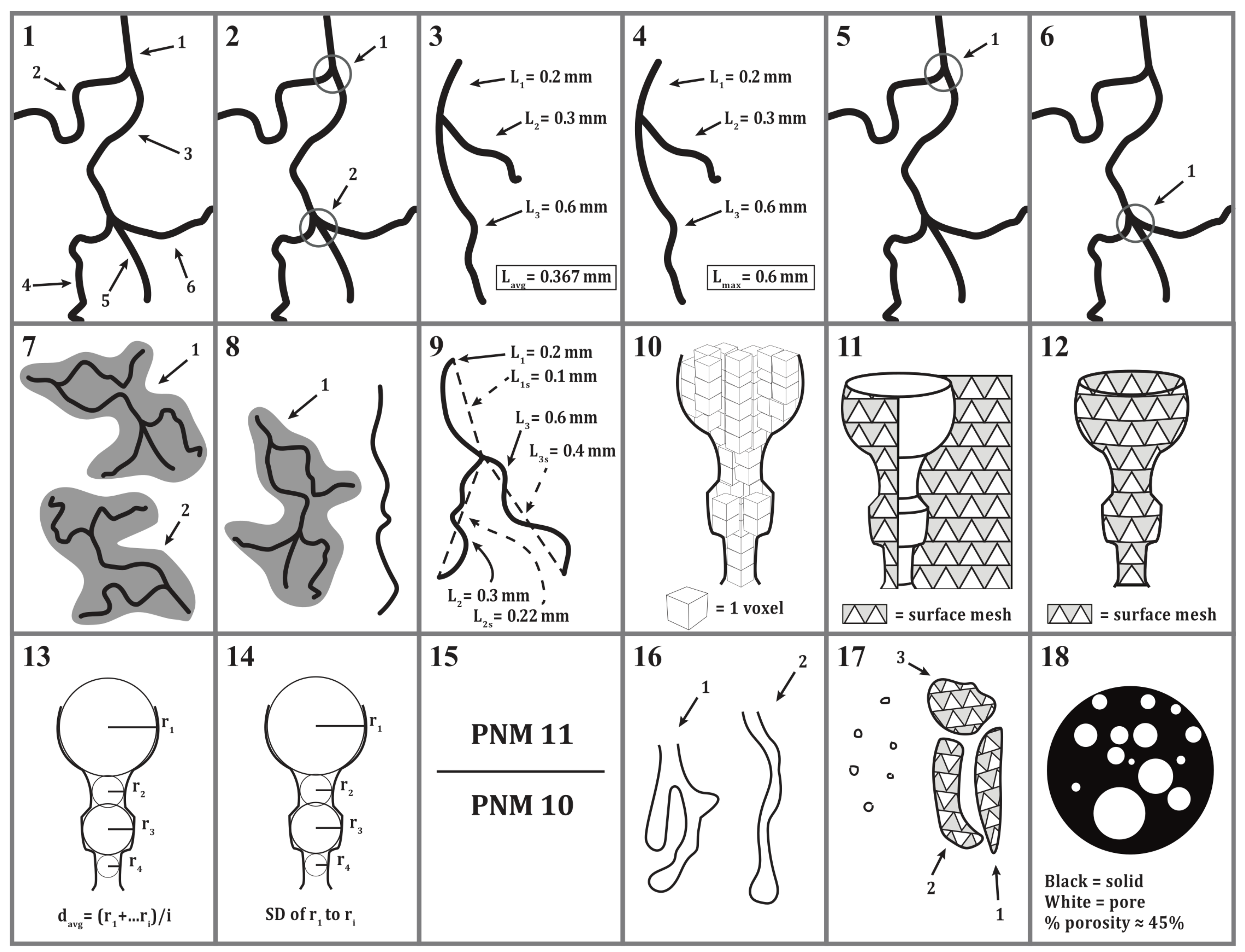
| Metric Number | Pore Network Metric (PNM) | Unit | Metric Description |
|---|---|---|---|
| 1 | Number of branches | Count | The number of slab segments (composites of slab voxels) in a VoI |
| 2 | Total number of junctions | Count | The total number of voxels in the VoI with more than two neighbor voxels |
| 3 | Mean branch length | mm | Average length of a branch in the VoI; calculated using all branches in the VoI |
| 4 | Maximum branch length | mm | Length of the longest branch in the VoI |
| 5 | Number of triple points | Count/mL | The number of junctions in the VoI with exactly three branches, expressed as a count per unit volume |
| 6 | Number of quadruple points | Count/mL | The number of junctions in the VoI with exactly three branches, expressed as a count per unit volume. |
| 7 | Total number of skeletons | Count | Number of individual (non-connected) skeleton (centerline) networks in the VoI |
| 8 | Number of skeletons with branches >1 | Count | The number of skeleton networks that contain at least one junction and branch |
| 9 | Mean tortuosity | n/a | Mean convolution of all pores in the VoI. Calculated as the sum of all total branch lengths in the sample divided by the sum of the straight-line distances of all branches in the VoI [50] |
| Metric Number | Pore Network Metric (PNM) | Unit | Metric Description |
|---|---|---|---|
| 10 | Image based void volume | mm3 | Volume occupied by an individual pore. Reported as average pore volume for each sample. Calculated by counting the number of voxels contained within a given void |
| 11 | Void surface area | mm2 | Calculated by fitting a triangular surface mesh (via marching cubes) to the interior of each individual void [51] |
| 12 | Enclosed void volume | mm3 | Volume of an individual void enclosed by triangular surface mesh (0 if no mesh could be fit) |
| 13 | Mean pore diameter | mm | Calculated at several points as the diameter of the greatest sphere that fits within the void and which contains the point |
| 14 | Standard deviation of mean pore diameter | mm | Standard deviation of sphere diameters used in mean pore diameter calculation |
| 15 | Surface area to volume ratio | mm−1 | Surface area divided by image based void volume |
| 16 | Total number of individual voids | Count | Number of individual voids identified in the VoI |
| 17 | Number of individual voids with enclosed volume > 0 | Count | The number of voids to which a triangular surface mesh was fit in the VoI |
| 18 | Image based porosity | % | Number of void voxels in the VoI divided by the total number of voxels in the VoI |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wanzek, T.; Keiluweit, M.; Varga, T.; Lindsley, A.; Nico, P.S.; Fendorf, S.; Kleber, M. The Ability of Soil Pore Network Metrics to Predict Redox Dynamics Is Scale Dependent. Soil Syst. 2018, 2, 66. https://doi.org/10.3390/soilsystems2040066
Wanzek T, Keiluweit M, Varga T, Lindsley A, Nico PS, Fendorf S, Kleber M. The Ability of Soil Pore Network Metrics to Predict Redox Dynamics Is Scale Dependent. Soil Systems. 2018; 2(4):66. https://doi.org/10.3390/soilsystems2040066
Chicago/Turabian StyleWanzek, Thomas, Marco Keiluweit, Tamas Varga, Adam Lindsley, Peter S. Nico, Scott Fendorf, and Markus Kleber. 2018. "The Ability of Soil Pore Network Metrics to Predict Redox Dynamics Is Scale Dependent" Soil Systems 2, no. 4: 66. https://doi.org/10.3390/soilsystems2040066
APA StyleWanzek, T., Keiluweit, M., Varga, T., Lindsley, A., Nico, P. S., Fendorf, S., & Kleber, M. (2018). The Ability of Soil Pore Network Metrics to Predict Redox Dynamics Is Scale Dependent. Soil Systems, 2(4), 66. https://doi.org/10.3390/soilsystems2040066