
Review of Phosphite as a Plant Nutrient and Fungicide


Abstract
:1. Introduction
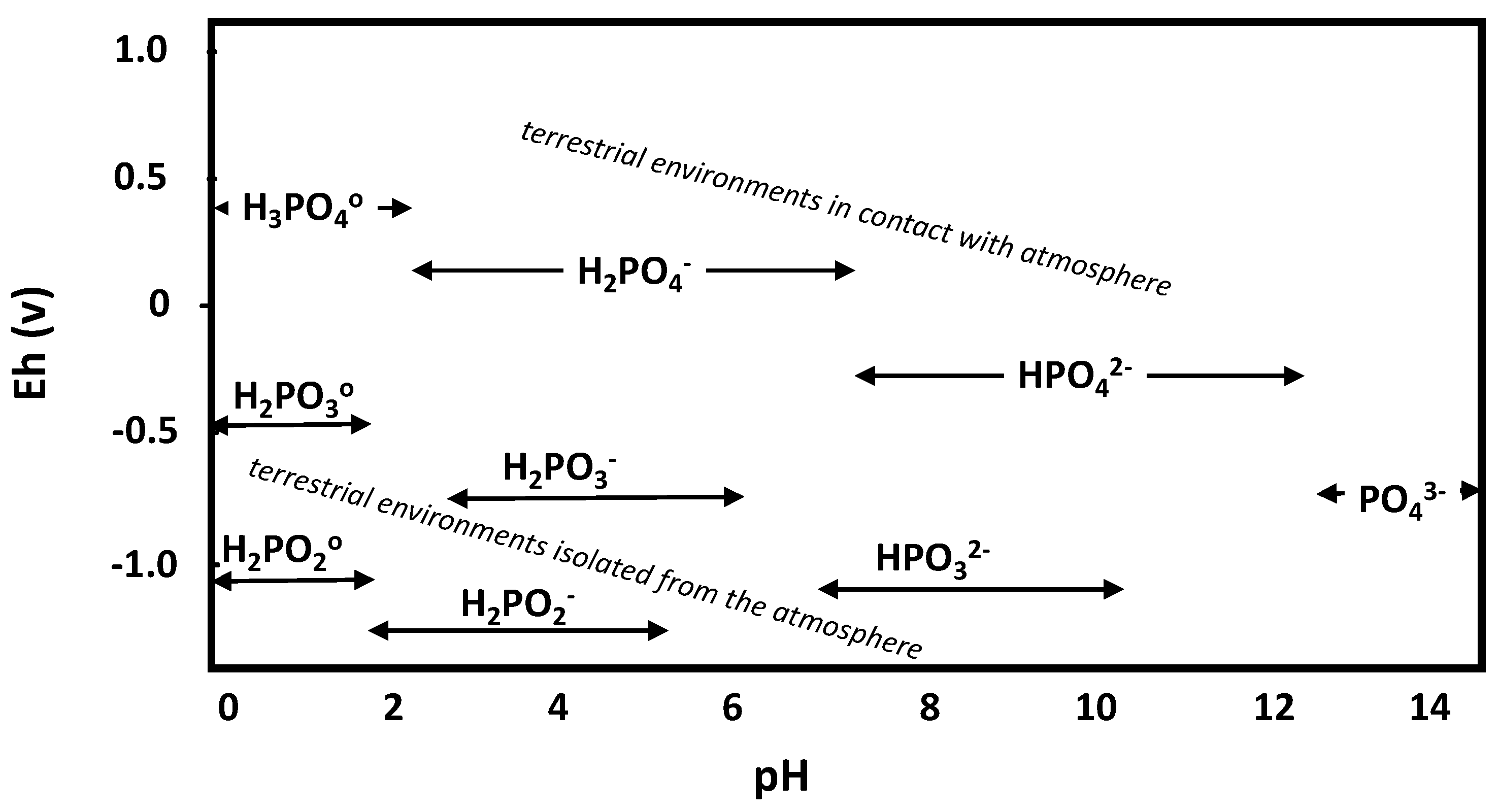
2. Reduced Phosphorus Chemistry in Soil
3. Microbial Oxidation of Phi to Pi
4. Phi Uptake, Translocation, and Utilization in Plants
5. Phi Use as a Plant Nutrient Source
- With any study conducted in hydroponic nutrient solutions, oxidation of Phi to Pi will be limited, although maintaining Phi throughout the study is critical to evaluating plant response to Phi compared to Pi.
- Most studies do not include an assessment of fungal infections or their control with Phi treatment.
- In studies conducted in soil or other potting media, Phi oxidation to Pi is not generally assessed. More importantly, the residual availability of soil applied Phi is not commonly quantified.
- Although few have documented the potential for Phi oxidation in plant cells, most studies do not assess Phi to Pi transformation in the plant, critical to assessing Phi oxidation potential in the plant.
- Results suggesting increased P nutrition by measuring total P (%) need to be moderated with the nutrient concentrating effects of reductions in biomass yield.
6. Phi Use as a Plant Fungicide
- Use of Phi products may result in accumulation of presence of Phi residues in horticultural products. For example, the European Union established a 2 ppm maximum residue level (MRL) for Phi in horticultural products [125]. It takes relatively small foliar or soil Phi application rates to result in Phi residues in marketable products [115].
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- MacIntire, W.; Winterberg, S.; Hardin, L.; Sterges, A.; Clements, L. Fertilizer evaluation.of certain phosphorus, phosphorous, and phosphoric materials by means of pot cultures. Agron. J. 1950, 42, 543–549. [Google Scholar] [CrossRef]
- Jackman, R.; Lambert, J.; Rothbaum, H. Red phosphorus as a fertiliser for grass-clover pasture. New Zealand J. Agric. Res. 1970, 13, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Guest, D.; Grant, B. The complex action of phosphonates as antifungal agents. Biol. Rev. 1991, 66, 159–187. [Google Scholar] [CrossRef]
- Lovatt, C.; Mikkelsen, R. Phosphite fertilizers: What are they? Can you use them? What can they do? Better Crops 2006, 90, 11–13. [Google Scholar]
- Hanrahan, G.; Salmassi, T.; Khachikian, C.; Foster, K. Reduced inorganic phosphorus in the natural environment: Significance, speciation and determination. Talanta 2005, 66, 435–444. [Google Scholar] [CrossRef]
- Pasek, M.; Harnmeijer, J.; Buick, R.; Maheen, G.; Atlas, Z. Evidence for reactive reduced phosphorus species in the early Archean ocean. Proc. Natl. Acad. Sci. USA 2013, 110, 10089–10094. [Google Scholar] [CrossRef] [Green Version]
- Glindemann, D.; Bergmann, A.; Stottmeister, U.; Gassmann, G. Phosphine in the lower terrestrial troposphere. Naturwissenschaften 1996, 83, 131–133. [Google Scholar] [CrossRef]
- Pasek, M. Rethinking early Earth phosphorus geochemistry. Proc. Natl. Acad. Sci. USA 2008, 105, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Morton, S.; Edwards, M. Reduced Phosphorus Compounds in the Environment. Crit. Rev. Environ. Sci. Technol. 2005, 35, 333–364. [Google Scholar] [CrossRef]
- Lindsay, W.L. Chemical Equilibria in Soils; John Wiley & Sons: New York, NY, USA, 1979. [Google Scholar]
- McDonald, A.; Grant, B.; Plaxton, W. Phosphite (phosphorous acid): Its relevance in the environment and agriculture and influence on plant phosphate starvation response. J. Plant. Nutr. 2001, 24, 1505–1519. [Google Scholar] [CrossRef]
- Schink, B.; Friedrich, M. Bacterial metabolism: Phosphite oxidation by sulphate reduction. Nature 2000, 406, 37. [Google Scholar] [CrossRef] [PubMed]
- Morton, S.; Glindemann, D.; Edwards, M. Phosphates, Phosphites, and Phosphides in Environmental Samples. Environ. Sci. Technol. 2003, 37, 1169–1174. [Google Scholar] [CrossRef] [PubMed]
- Pasek, M.; Sampson, J.; Atlas, Z. Redox chemistry in the phosphorus biogeochemical cycle. Proc. Natl. Acad. Sci. USA 2014, 111, 15468–15473. [Google Scholar] [CrossRef] [Green Version]
- Benitez-Nelson, C. The biogeochemical cycling of phosphorus in marine systems. Earth Sci. Rev. 2000, 51, 109–135. [Google Scholar] [CrossRef]
- Huang, L.; Jia, X.; Zhang, G.; Shao, M. Soil organic phosphorus transformation during ecosystem development: A review. Plant. Soil. 2017, 417, 17–42. [Google Scholar] [CrossRef]
- Cade-Menun, B.; Berch, S.; Preston, C.; Lavkulich, L. Phosphorus forms and related soil chemistry of Podzolic soils on northern Vancouver Island I A comparison of two forest types. Can. J. For. Res. 2000, 30, 1714–1725. [Google Scholar] [CrossRef]
- Hedley, M.; Stewart, J.; Chauhan, B. Changes in the inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubation. Soil Sci. Soc. Am. J. 1982, 46, 970–976. [Google Scholar] [CrossRef]
- Condron, L.M.; Newman, S. Revisiting the fundamentals of phosphorous fractionation of sediments and soil. J. Soils Sediments 2011, 11, 830–840. [Google Scholar] [CrossRef]
- Havlin, J.L.; Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. Soil Fertility and Nutrient Management: An Introduction to Nutrient Management, 8th ed.; Pearson: Upper Saddle River, NJ, USA, 2014; p. 516. [Google Scholar]
- Adams, F.; Conrad, J. Transition of Phosphite to Phosphate in Soils. Soil Sci. 1953, 75, 361–371. [Google Scholar] [CrossRef]
- Casida, L. Microbial oxidation and utilization of orthophosphite during growth. J. Bacteriol. 1960, 80, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Malacinski, G.; Konetzka, W. Bacterial oxidation of orthophosphite. J. Bacteriol. 1966, 91, 578–582. [Google Scholar] [CrossRef] [Green Version]
- Loera-Quezada, M.; Leyva-Gonzalez, M.; Lopez-Arredondo, D.; Herrara-Estrella, L. Phosphite cannot be used as a phosphorus source but is non-toxic for microalgae. Plant. Sci. 2015, 231, 124–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezuidenhout, J.; Darvas, J.; Kotze, J. The dynamics and distribution of phosphite in avocado trees treated with phosetyl-Al. S. Afr. Avocado Grow. Assoc. Yearb. 1987, 10, 101–103. [Google Scholar]
- Ohtake, H.; Wu, H.; Imazu, K.; Anbe, Y.; Kato, J.; Kuroda, A. Bacterial phosphonate degradation, phosphite oxidation and polyphosphate accumulation. Resour. Conserv. Recycl. 1996, 18, 125–134. [Google Scholar] [CrossRef]
- Imazu, K.; Tanaka, S.; Kuroda, A.; Anbe, Y.; Kato, J.; Ohtake, H. Enhanced Utilization of Phosphonate and Phosphite by Klebsiella aerogenes. Appl. Environ. Microbiol. 1998, 64, 3754–3758. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, W.; Wolfe, R. Molecular Genetic Analysis of Phosphite and Hypophosphite Oxidation by Pseudomonas stutzeri WM88. J. Bacteriol. 1998, 180, 5547–5558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, S.; Glindemann, D.; Wang, X.; Niu, X.; Edwards, M. Analysis of Reduced Phosphorus in Samples of Environmental Interest. Environ. Sci. Technol. 2005, 39, 4369–4376. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.; Winans, L.; Helms, S. Anaerobic utilization of phosphite and hypophosphite by Bacillus sp. Appl. Environ. Microbiol. 1978, 35, 937–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pao, S.; Paulsen, I.; Saier, M. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Costas, A.; White, A.; Metcalf, W. Purification and Characterization of a Novel Phosphorus-oxidizing Enzyme from Pseudomonas stutzeri WM88. J. Biol. Chem. 2001, 276, 17429–17436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, J.; Fogle, E.; Christman, H.; Johannes, T.; Zhao, H.; Metcalf, W.; van der Donk, W. Investigation of the Role of Arg301 Identified in the X-ray Structure of Phosphite Dehydrogenase. Biochemistry 2012, 51, 4254–4262. [Google Scholar] [CrossRef] [PubMed]
- Schink, B.; Thiemann, V.; Laue, H.; Friedrich, M. Desulfotignum phosphitoxidans sp. nov., a new marine sulfate reducer that oxidizes phosphite to phosphate. Arch. Microbiol. 2002, 177, 381–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeonova, D.; Wilson, M.; Metcalf, W.; Schink, B. Identification and Heterologous Expression of Genes Involved in Anaerobic Dissimilatory Phosphite Oxidation by Desulfotignum phosphitoxidans. J. Bacteriol. 2010, 192, 5237–5244. [Google Scholar] [CrossRef] [Green Version]
- Howard, K.; Dell, B.; Hardy, G. Phosphite and mycorrhizal formation in seedlings of three Australian Myrtaceae. Aust. J. Botany. 2001, 48, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, W.; Wanner, B. Involvement of the Escherichia coli phn (psiD) gene cluster in assimilation of phosphorus in the form of phosphonates, phosphite, Pi esters, and Pi. J. Bacteriol. 1991, 173, 587–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodyer, R.; van der Donk, W.; Zhao, H. Relaxing the nicotinamide cofactor specificity of phosphite dehydrogenase by rational design. Biochemistry 2003, 42, 11604–11614. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Metcalf, W. A new activity for an old enzyme: Escherichia coli bacterial alkaline phosphatase is a phosphite-dependent hydrogenase. Proc. Natl. Acad. Sci. USA 2004, 101, 7919–7924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, A.; Metcalf, W. Two C-P lyase operons in Pseudomonas stutzeri and their roles in the oxidation of phosphonates, phosphite, and hypophosphite. J. Bacteriol. 2004, 186, 4730–4739. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, I.; Coates, J. Microbial Phosphite Oxidation and Its Potential Role in the Global Phosphorus and Carbon Cycles. Adv. Appl. Microbiol. 2017, 98, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S. Phosphate-Solubilizing Microorganisms: Mechanism and Their Role in Phosphate Solubilization and Uptake. J. Soil Sci. Plant. Nutr. 2020, 21, 49–68. [Google Scholar] [CrossRef]
- Tian, J.; Ge, F.; Zhang, D.; Deng, S.; Liu, X. Roles of Phosphate Solubilizing Microorganisms from Managing Soil Phosphorus Deficiency to Mediating Biogeochemical P Cycle. Biology 2021, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotech. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Rafi, M.; Krishnaveni, M.; Charyulu, P. Phosphate-Solubilizing Microorganisms and their Emerging Role in Sustainable Agriculture. In Recent Developments in Applied Microbiology and Biochemistry; Buddolla, S., Ed.; Elsevier: London, UK, 2019; pp. 223–233. [Google Scholar]
- Raymond, N.; Gomez-Munoz, B.; van der Bom, F.; Nybroe, O.; Jensen, L.; Muller-Stover, D.; Oberson, A.; Richardson, A. Phosphate-solubilising microorganisms for improved crop productivity: A critical assessment. New Phytol. 2021, 229, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- White, A.; Metcalf, W. Microbial Metabolism of Reduced Phosphorus Compounds. Ann. Rev. Microbiol. 2007, 61, 379–400. [Google Scholar] [CrossRef] [PubMed]
- Fontana, M.; Bragazza, L.; Guillaime, T.; Santonja, M.; Buttler, A.; Elfouki, S.; Sinaj, S. Valorization of calcium phosphite waste as phosphorus fertilizer: Effects on green manure productivity and soil properties. J. Environ. Manag. 2021, 285, 112061. [Google Scholar] [CrossRef] [PubMed]
- Kehler, A.; Haygarth, P.; Tamburini, F.; Blackwell, M. Cycling of reduced phosphorus compounds in soil and potential impacts of climate change. Eur. J. Soil Sci. 2021, 2021, 1–21. [Google Scholar]
- Griffith, J.; Coffey, M.; Grant, B. Phosphonate inhibition as a function of phosphate concentration in isolates of Phytophthora palmivova. J. Gen. Microbiol. 1993, 139, 2109–2116. [Google Scholar] [CrossRef] [Green Version]
- Ratjen, A.; Gerendás, J. A critical assessment of the suitability of phosphite as a source of phosphorus. J. Plant. Nutr. Soil Sci. 2009, 172, 821–828. [Google Scholar] [CrossRef]
- Thao, H.; Yamakawa, T.; Myint, A.; Sarr, P. Effects of phosphite, a reduced form of phosphate, on the growth and phosphorus nutrition of spinach (Spinacia oleracea L.). Soil Sci. Plant. Nutr. 2008, 54, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Orbovic, V.; Syvertsen, J.; Bright, D.; van Clief, D.; Graham, J. Citrus seedling growth and susceptibility to root rot as affected by phosphite and phosphate. J. Plant. Nutr. 2008, 31, 774–787. [Google Scholar] [CrossRef]
- Schroetter, S.; Angeles-Wedler, D.; Kreuzig, R.; Schnug, E. Effects of phosphite on phosphorus supply and growth of corn (Zea mays). Landbauforsch. Völkenrode 2006, 56, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Danova-Alt, R.; Dijema, C.; Dewaard, P.; Kock, M. Transport and compartmentation of phosphite in higher plant cells–kinetic and 31P nuclear magnetic resonance studies. Plant. Cell Environ. 2008, 31, 1510–1521. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Tsai, P.; Shya, Y.; Sheu, F. The effects of phosphite on phosphate starvation responses of Ulva lactuca (Ulvales, Chlorophyta). J. Phycol. 2005, 41, 975–982. [Google Scholar] [CrossRef]
- Carswell, M.; Grant, B.; Plaxton, W. Disruption of the phosphate-starvation response of oilseed rape suspension cells by the fungicide phosphonate. Planta 1997, 203, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Carswell, M.; Grant, B.; Theodorou, M.; Harris, J.; Niere, J.; Plaxton, W. The fungicide phosphonate disrupts the phosphate-starvation response in Brassica nigra seedlings. Plant Physiol. 1996, 110, 105–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Förster, H.; Adaskaveg, J.; Kim, D.; Stanghellini, M. Effect of Phosphite on Tomato and Pepper Plants and on Susceptibility of Pepper to Phytophthora Root and Crown Rot in Hydroponic Culture. Plant. Dis. 1998, 82, 1165–1170. [Google Scholar] [CrossRef] [Green Version]
- Varadarajan, D.; Karthikeyan, A.; Matilda, P.; Raghothama, K. Phosphite, an Analog of Phosphate, Suppresses the Coordinated Expression of Genes under Phosphate Starvation. Plant. Physiol. 2002, 129, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Thao, H.; Yamakawa, T. Phosphite (phosphorous acid): Fungicide, fertilizer or bio-stimulator? Soil Sci. Plant. Nutr. 2009, 55, 228–234. [Google Scholar] [CrossRef]
- Ticconi, C.; Delatorre, C.; Abel, S. Attenuation of Phosphate Starvation Responses by Phosphite in Arabidopsis. Plant. Physiol. 2001, 127, 963–974. [Google Scholar] [CrossRef]
- Ticconi, C.; Abel, S. Short on phosphate: Plant surveillance and countermeasures. Trends Plant. Sci. 2004, 9, 548–555. [Google Scholar] [CrossRef]
- Singh, V.; Wood, S.; Knowles, V.; Plaxton, W. Phosphite accelerates programmed cell death in phosphate starved oilseed rape (Brassica napus) suspension cell cultures. Planta 2003, 218, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Merino, F.; Trejo-Tellez, L. Conventional and novel uses of phosphite in horticulture: Potentialities and challenges. Italus Hortes 2016, 23, 1–13. [Google Scholar]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant. Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera-Estrella, L.; Lopez-Arredondo, D.; Heriberto, A. 2012. Fungi Adapted to Metabolize Phosphite as a Source of Phosphorus. U.S. Patent Application 2012/0285210 A1, 15 November 2012. [Google Scholar]
- Lopez-Arredondo, D.; Herrera-Estrella, L. Engineering phosphorus metabolism in plants to produce a dual fertilization and weed control system. Nat. Biotech. 2012, 30, 889–893. [Google Scholar] [CrossRef]
- Manna, M.; Archary, V.; Islam, T.; Agrawal, P.; Reddy, M. The development of a phosphite mediated fertilization and weed control system for rice. Sci. Rep. 2016, 6, 24941. [Google Scholar] [CrossRef]
- Ram, B.; Fartyal, D.; Sheri, V.; Varakumar, P.; Borphukan, B.; James, D.; Yadav, R.; Bhatt, A.; Achary, P.A.V.; Reddy, M. Characterization of phoA, a bacterial alkaline phosphatase for Phi use efficiency in rice plant. Front. Plant. Sci. 2019, 10, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Pandeya, D.; Lopez-Arrendondo, D.; Janga, M.; Campbell, L.; Estrella-Hernandez, P.; Bagavathiannan, M.; Herrera-Estrella, L.; Rathore, K. Selective fertilization with phosphite allows unhindered growth of cotton plants expressing the ptxD gene while suppressing weeds. Proc. Natl. Acad. Sci. USA 2018, 115, E6946–E6955. [Google Scholar] [CrossRef] [Green Version]
- Achary, V.; Ram, B.; Manna, M.; Datta, D.; Bhatt, A.; Reddy, M.; Agrawal, P. Phosphite: A novel P fertilizer for weed management and pathogen control. Plant Biotech. J. 2017, 15, 1493–1508. [Google Scholar] [CrossRef] [Green Version]
- Lucas, L. Development and management of summer decline of bentgrass. PACE Insights 1995, 1, 2–3. [Google Scholar]
- Dorer, S. Nutritional Effects of a Fungicide Combination on Summer Bentgrass Decline. Master’s Thesis, North Carolina State University, Raleigh, NC, USA, 1996. [Google Scholar]
- Thao, H.; Yamakawa, T. Growth of celery (Apium graveolens var. dulce) as influenced by phosphite. J. Fac. Agric. Kyushu Univ. 2008, 53, 375–378. [Google Scholar] [CrossRef]
- Lovatt, C. A definitive test to determine whether phosphite fertilization can replace phosphate fertilization to supply P in the metabolism of “Hass” on Duke 7. Calif. Avoc. Soc.Yearb. 1990, 74, 61–64. [Google Scholar]
- Lovatt, C. Timing citrus and avocado foliar nutrient applications to increase fruit set and size. HortTechnology 1999, 9, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Lovatt, C. Properly timing foliar applied fertilizers increases efficacy: A review and update on timing foliar nutrient applications to citrus and avocado. Hort. Technol. 2013, 23, 536–541. [Google Scholar] [CrossRef] [Green Version]
- Zambrosi, F.; Mattos, D.; Syvertsen, J. Plant growth, leaf photosynthesis, and nutrient-use efficiency of citrus rootstocks decrease with phosphite supply. J. Plant. Nutr. Soil Sci. 2011, 174, 487–495. [Google Scholar] [CrossRef]
- Ávila, F.; Faquin, V.; Silva, D.; Bastos, C.; Oliveira, N.; Soares, D. Phosphite as Phosphorus Source to Grain Yield of Common Bean Plants Grown in Soils Under Low or Adequate Phosphate Availability. Ciênc. Agrotec. 2012, 36, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Ávila, F.; Faquin, V.; Lobato, A.; Ávila, P.; Marques, D.; Guedes, E.; Tan, D. Effect of phosphite supply in nutrient solution on yield, phosphorus nutrition and enzymatic behavior in common bean (Phaseolus vulgaris L.) plants. Aust. J. Crop. Sci. 2013, 7, 713–722. [Google Scholar]
- Ávila, F.; Faquin, V.; Araújo, J.; Marques, D.; Ribeiro, P.; Lobato, A.; Ramos, S.; Baliza, D. Phosphite supply affects phosphorus nutrition and biochemical responses in maize plants Austr. J. Crop. Sci. 2011, 5, 646–653. [Google Scholar]
- Sutradhar, A.; Arnall, D.; Dunn, B.; Raun, W. Does phosphite, a reduced form of phosphate contribute to phosphorus nutrition in corn (Zea mays L.)? J. Plant. Nutr. 2019, 42, 982–989. [Google Scholar] [CrossRef]
- Constán-Aguilar, C.; Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.; Camacho, M.; Romero, L.; Ruiz, J.M.; Blasco, B. Physiological and Nutritional Evaluation of the Application of Phosphite as a Phosphorus Source in Cucumber Plants. Commun. Soil Sci. Plant. Anal. 2014, 45, 204–222. [Google Scholar] [CrossRef]
- Thao, H.; Yamakawa, T. Phosphate absorption of intact komatsuna plants as influenced by phosphite. Soil Sci. Plant. Nutr. 2010, 56, 133–139. [Google Scholar] [CrossRef]
- Estrada-Ortiz, E.; Trejo-Téllez, L.; Gómez-Merino, F.; Núñez-Escobar, R.; Sandoval-Villa, M. The effects of phosphite on strawberry yield and fruit quality. J. Soil Sci. Plant. Nutr. 2013, 13, 612–620. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Ortiz, E.; Trejo-Téllez, L.; Gómez-Merino, F.; Silva-Rojas, H.; Castillo-González, A.; Avitia-García, E. Physiological Responses of Chard and Lettuce to Phosphite Supply in Nutrient Solution. J. Agr. Sci. Tech. 2016, 18, 1079–1090. [Google Scholar]
- Glinicki, R.; Sas-Paszt, L.; Jadczuk-Tobjasz, E. The Effect of Plant Stimulant/Fertilizer “Resistim” on growth and Development of Strawberry Plants. J. Fruit Ornament. Plant. Res. 2010, 18, 111–124. [Google Scholar]
- Hirosse, E.; Creste, J.; Custódio, C.; Barbosa, N.; Machado-Neto, N. In vitro growth of sweet potato fed with potassium phosphite. Acta Sci. Agron. 2012, 34, 85–91. [Google Scholar] [CrossRef]
- Vinas, M.; Mendez, J.; Jiménez, V. Effect of foliar applications of phosphites on growth, nutritional status and defense responses in tomato plants. Sci. Hortic. 2020, 265, 109200. [Google Scholar] [CrossRef]
- Ali, M.; Sutradhar, A.; Edano, M.; Edwards, J.; Girma, K. Response of Winter Wheat Grain Yield and Phosphorus Uptake to Foliar Phosphite Fertilization. Int. J. Agron. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Albrigo, L. Effects of foliar applications of urea or Nutriphite on flowering and yields of Valencia orange trees. Proc. Florida State Hortic. Soc. 1999, 112, 1–4. [Google Scholar]
- Rickard, D. Review of Phosphorus Acid and Its Salts as Fertilizer Materials. J. Plant. Nutr. 2000, 23, 161–180. [Google Scholar] [CrossRef]
- Jost, R.; Pharmawati, M.; Lapis-Gaza, H.; Rossig, C.; Berkowitz, O.; Lambers, H.; Finnegan, P. Differentiating phosphate-dependent and phosphate-independent systemic phosphate-starvation response networks in Arabidopsis thaliana through the application of phosphite. J. Exp. Bot. 2016, 66, 2501–2514. [Google Scholar] [CrossRef] [Green Version]
- Panigrahy, M.; Rao, D.; Sarla, N. Molecular mechanisms in response to phosphate starvation in rice. Biotech. Adv. 2009, 27, 389–397. [Google Scholar] [CrossRef]
- Abel, S.; Ticconi, C.; Delatorre, C. Phosphate sensing in higher plants. Physiol. Plant. 2002, 115, 1–8. [Google Scholar] [CrossRef]
- Raghothama, K. Phosphate transport and signaling. Curr. Opin. Plant. Biol. 2000, 3, 182–187. [Google Scholar] [CrossRef]
- Marschner, H. Mineral. Nutrition in Plants, 2nd ed.; Academic Press: San Diego, CA, USA, 1995. [Google Scholar]
- Deliopoulus, T.; Kettlewell, P.; Hare, M. Fungal disease suppression by inorganic salts: A review. Crop. Prot. 2010, 29, 1059–1075. [Google Scholar] [CrossRef]
- Gomez-Merino, F.; Trejo-Tellez, L. Biostimulant activity of phosphite in horticulture. Sci. Hort. 2015, 196, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Hardy, G.; Barrett, S.; Shearer, B. The future of phosphite as a fungicide to control the soilborne plant pathogen Phytophthora cinnamomi in natural ecosystems. Australas. Plant Pathol. 2001, 30, 133–139. [Google Scholar] [CrossRef]
- Silva, O.; Santos, H.; Pria, M.D.; Mio, L.M. Potassium phosphite for control of downy mildew of soybean. Crop. Prot. 2011, 30, 598–604. [Google Scholar] [CrossRef]
- Shearer, B.; Fairman, R. Application of phosphite in a high-volume foliar spray delays and reduces the rate of mortality of four Banksia species infected with Phytophthora cinnamomi. Australas. Plant. Path. 2007, 36, 358–368. [Google Scholar] [CrossRef]
- Oka, Y.; Tkachi, N.; Mor, M. Phosphite Inhibits Development of the Nematodes Heterodera avenae and Meloidogyne marylandi in Cereals. Phytopath 2007, 97, 396–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, J.H. Phosphite for control of phytophthora diseases in citrus: Model for management of phytophthora species on forest tress? New Zealand J. For. Sci. 2011, 41, S49–S56. [Google Scholar]
- Reuveni, M.; Sheglov, D.; Cohen, Y. Control of moldy-core decay in apple fruits by β-aminobutyric acids and potassium phosphites. Plant Dis. 2003, 87, 933–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervera, M.; Cautin, R.; Jeria, G. Evaluation of calcium phosphite; magnesium phosphite and potassium phosphite in the control of phytophthora cinnamomic in Hass avocado trees (Persea americana Mill) grown in container. In Proceedings of the VI World Avocado Congress (Actas VI Congreso Mundial del Aguacate), Viña Del Mar, Chile, 12–16 November 2007; ISBN 978-956-17-0413-8. [Google Scholar]
- Barrett, S.; Shearer, B.; Hardy, G. The efficacy of phosphite applied after inoculation on the colonisation of Banksia brownii stems by Phytophthora cinnamomic. Australas. Plant Pathol. 2003, 32, 1–7. [Google Scholar] [CrossRef]
- Abbasi, P.; Lazarovits, G. Effect of soil application of AG3 phosphonate on the severity of clubroot of bok choy and cabbage caused by Plasmodiophora brassicae. Plant Dis. 2006, 90, 1517–1522. [Google Scholar] [CrossRef] [Green Version]
- Gentile, S.; Valentino, D.; Tamietti, G. Control of Ink Disease by truck Injection of Potassium Phosphite. J. Plant. Path. 2009, 91, 565–571. [Google Scholar]
- Abbasi, P.; Lazarovits, G. Seed treatment with phosphonate (AG3) suppresses Pythium damping-off of cucumber seedlings. Plant Dis. 2006, 90, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Speiser, B.; Berner, A.; Haseli, A.; Tamm, L. Control of downy mildew of grapevine with potassium phosphonate: Effectivity and phosphonate residues in wine. Biol. Agric. Hortic. 2000, 17, 305–312. [Google Scholar] [CrossRef]
- Smillie, R.; Grant, B.; Guest, D. The Mode of Action of Phosphite: Evidence for Both Direct and Indirect Modes of Action on Three Phytophthora sp. in Plants. Phytopath 1989, 79, 921–926. [Google Scholar] [CrossRef] [Green Version]
- Panicker, S.; Gangadharan, K. Controlling downy mildew of maize caused by Peronosclerospora sorghi by foliar sprays of phosphoric acid compounds. Crop Prot. 1999, 18, 115–118. [Google Scholar] [CrossRef]
- Bock, C.; Brenneman, T.; Hotchkiss, M.; Wood, B. Evaluation of phosphite fungicide to control pecan scab in the southeastern USA. Crop. Prot. 2012, 36, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Liljeroth, E.; Lankinen, A.; Wiik, L.; Burra, D.; Alexandersson, E.; Andreasson, E. Potassium phosphite combined with reduced doses of fungicides provides efficient protection against potato late blight in large-scale field trails. Crop. Prot. 2016, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, M.; Han, X.; Zhang, Z.; Xi, Y.; Boorboori, M.; Wang-Pruski, G. Phosphite Application Alleviates Pythophthora infestans by Modulation of Photosynthetic and Physio-Biochemical Metabolites in Potato Leaves. Pathogens 2020, 9, 170. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; Inglis, D.; Miller, J. Control of potato tuber rots caused by oomycetes with foliar applications of phosphorous acid. Plant. Dis. 2004, 88, 1153–1159. [Google Scholar] [CrossRef] [Green Version]
- Lobato, M.; Machinandiarena, M.; Tambascio, C.; Dosio, G.; Caldiz, D.; Daleo, G.; Andreu, A.; Olivieri, F. Effect of foliar applications of phosphite on post-harvest potato tubers. Eur. J. Plant. Path. 2011, 130, 155–163. [Google Scholar] [CrossRef]
- Rebollar-Alviter, A.; Madden, L.; Ellis, M. Pre- and post-infection activity of azoxystrobin, pyraclostrobin, mefenoxam, and phosphite against leather rot of strawberry, caused by Phytophthora cactorum. Plant Dis. 2007, 91, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Yogev, E.; Sadowsky, A.; Solel, Z.; Oren, Y.; Orbach, Y. The performance of potassium phosphite for controlling Alternaria brown spot of citrus fruit. J. Plant Dis. Prot. 2006, 113, 207–213. [Google Scholar] [CrossRef]
- Fenn, M.; Coffey, M. Quantification of Phosphonate and Ethyl Phosphonate in Tobacco and Tomato Tissues and its Significance for the Mode of Action of Two Phosphonate Fungicides. Phytopathology 1989, 79, 76–82. [Google Scholar] [CrossRef]
- Ouimette, D.; Coffey, M. Phosphonate levels in avocado (persea americana) seedlings and soil following treatment with fosetyl-Al or potassium phosphonate. Plant. Dis. 1989, 73, 212–215. [Google Scholar] [CrossRef]
- Jackson, T.; Burgess, T.; Colquhoun, I.; Hardy, G. Action of the Fungicide Phosphite on Eucalyptus marginata Inoculated with Phytophthora cinnamomi. Plant. Path. 2000, 49, 147–154. [Google Scholar] [CrossRef]
- Trinchera, A.; Parisi, B.; Baratella, V.; Roccuzzo, G.; Soave, I.; Bazzocchi, C.; Fichera, D.; Finotti, M.; Riva, F.; Mocciaro, G.; et al. Assessing the origin of phosphonic acid residues in organic vegetable and fruit crops: The Biofosf project multi-factor approach. Agronomy 2020, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Sala, F.; Costa, C.; Echer, M.; Martins, M.; Blat, S. Phosphite effect on hot and sweet pepper reaction to Phytophthora capsici. Sci. Agric. 2004, 61, 492–495. [Google Scholar] [CrossRef]
- Mayton, H.; Myers, K.; Fry, W. Potato late blight-The role of foliar Phosphonates applications in suppressing pre-harvest tuber infections. Crop. Protect. 2008, 27, 943–950. [Google Scholar] [CrossRef]
- Cooke, L.; Little, G. The effect of foliar application of phosphonate formulations on the susceptibility of potato tubers to late blight. Pest. Manag. Sci. 2002, 58, 17–25. [Google Scholar] [CrossRef]
- Han, X.; Xi, Y.; Zhang, Z.; Mohammadi, M.; Joshi, J.; Borza, T.; Wang-Pruski, G. Effects of phosphite as a plant biostimulant on metabolism and stress response for better plant performance in Solanum tuberosum. Ecotox. Environ. Safety 2021, 210, 111873. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Zhang, Z.; Xi, Y.; Han, H.; Lan, F.; Zhang, B.; Wang-Pruski, G. Effects of potassium phosphite on biochemical contents and enzymatic activities of Chinese potatoes inoculated by phytophthora infestans. Appl. Ecol. Environ. Res. 2019, 17, 4499–4514. [Google Scholar] [CrossRef]
- Vincelli, P.; Dixon, E. Performance of selected phosphite fungicides on greens. Golf. Course Manag. 2005, 73, 77–81. [Google Scholar]
- Landschoot, P.; Cook, J. Sorting out the Phosphonate Products. Golf. Course Manag. 2005, 73, 73–77. [Google Scholar]
- Cook, P. Inhibition of Pythium ssp. and Suppression of Pythium Blight and Anthracnose with Phosphonate Fungicides. Master’s Thesis, Pennsylvania State University, State College, PA, USA, 2009. [Google Scholar]
- Ervin, E.; McCall, D.; Horvath, B. Efficacy of phosphite fungicides and fertilizers for control of pythium blight on a perennial ryegrass fairway in Virginia. Appl. Turfgrass Sci. 2009. [Google Scholar] [CrossRef]
- McComb, J.; O’Brien, P.; Calver, M.; Staskowski, P.; Jardine, N.; Eshraghi, L.; Ellery, J.; Gilovitz, J.; Scott, P.; O’Brien, J.; et al. Research into Natural and Induced Resistance in Australian Native Vegetation of Phytophthora cinnamomi and Innovative Methods to Contain and/or Eradicate within Localised Incursions in Areas of High Biodiversity in Australia; No. 19/2005DEH Sub Project 19.2.2; Centre for Phytophthora Science and Management for the Australian Government Department of the Environment, Water, Heritage and the Arts: Perth, Australia, 2008. [Google Scholar]
- Lambers, H.; Ahmedi, I.; Berkowitz, O.; Dunne, C.; Finnegan, P.; Hardy, G.; Jost, R.; Laliberté, E.; Pearse, S.; Teste, F. Phosphorus nutrition of phosphorus-sensitive Australian native plants: Threats to plant communities in a global biodiversity hotspot. Cons. Physiol. 2013, 1, 1–21. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
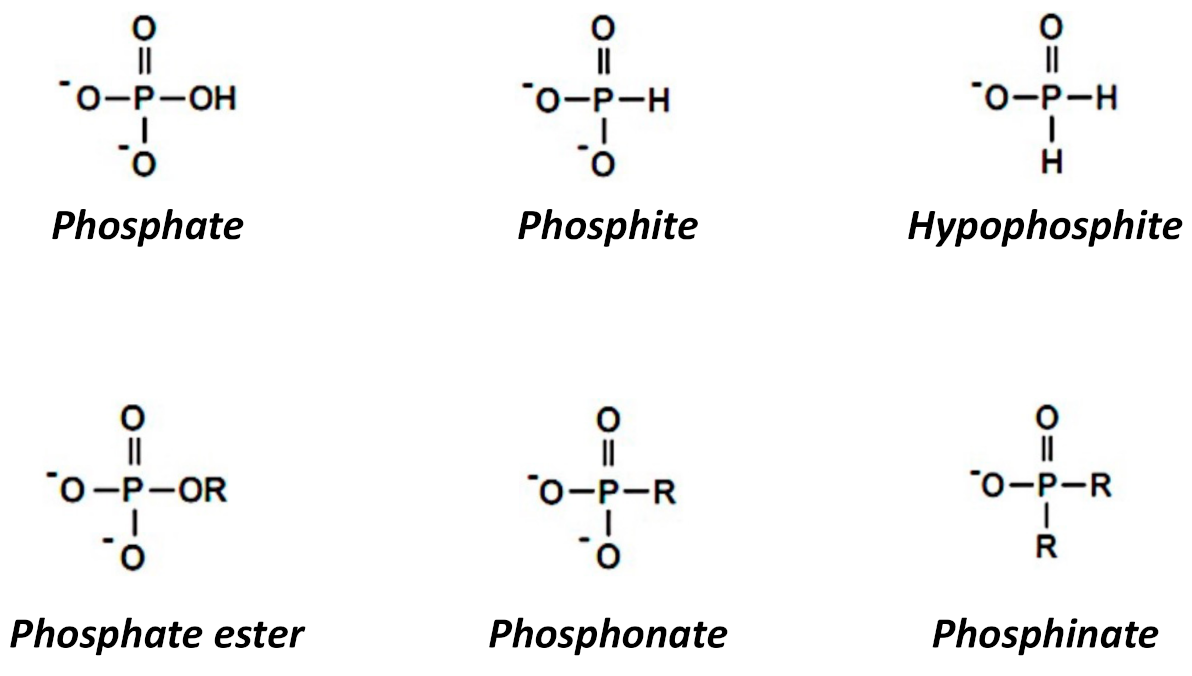
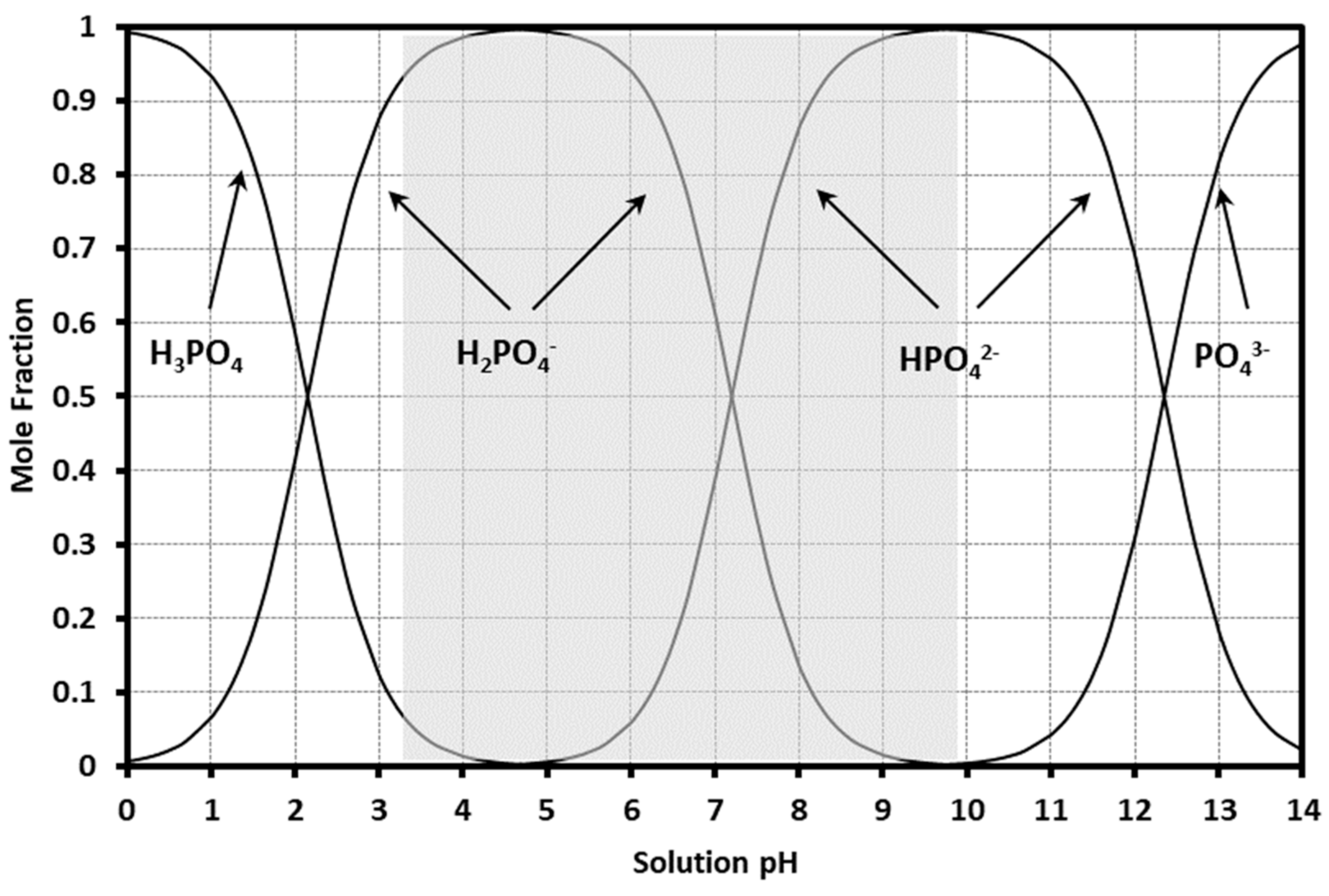
| Phosphorus Form 1 | Chemical Formula | Redox State 2 | Dissociation Reaction (in H2O) | Ka 3 |
|---|---|---|---|---|
| phosphoric acid | H3PO4 | +5 | H3PO40 ⇄ H2PO4− + H+ | 10−2.15 |
| phosphate | H2PO4−, HPO42− | H2PO4− ⇄ HPO42− + H+ | 10−7.2 | |
| HPO42− ⇄ PO43− + H+ | 10−12.35 | |||
| phosphorous acid (phosphonic acid) | H3PO3 | +3 | H3PO30 ⇄ H2PO3− + H+ | 10−1.5 |
| phosphite (phosphonate) | H2PO3−, HPO32− | H2PO3− ⇄ HPO32− + H+ | 10−6.79 | |
| hypophosphorus acid | H3PO2 | +1 | H3PO20 ⇄ H2PO2− + H+ | 10−1.1 |
| hypophosphite (phosphinate) | H2PO2− | |||
| phosphine | H3P | −3 | ||
| phosphonium | H4P+ | |||
| Plant | Application Method | Plant Response * | Reference |
|---|---|---|---|
| Arabidopsis | Hydroponic | Negative | [62] |
| Bentgrass | Foliar | Yes | [73,74] |
| Celery, spinach | Hydroponic | Negative 1 | [75] |
| Citrus, avacado | Foliar | Yes | [76,77,78] |
| Citrus | Foliar | Negative | [79] |
| Common bean | Soil, foliar | Negative 1 | [80,81] |
| Corn | Foliar | Negative 1 | [54] |
| Corn | Hydroponic | Negative | [82] |
| Corn | Foliar | No | [83] |
| Cotton | Foliar | Yes 2 | [71] |
| Cucumber | Foliar | Negative 1 | [84] |
| Komatsuna | Hydroponic | Negative | [85] |
| Oat, mustard, pea, (lupin) | Soil | No (Negative) | [48] |
| Onion | Hydroponic | Negative | [58] |
| Red clover, ryegrass | Soil | No | [1] |
| Strawberry, lettuce, chard | Hydroponic | No | [86,87] |
| Strawberry | Hydroponic | No | [88] |
| Sweet potato | Tissue culture | No | [89] |
| Tomato | Hydroponic | Negative | [60] |
| Tomato, pepper | Hydroponic | Negative 1 | [59] |
| Tomato | Foliar | No | [90] |
| Winter wheat | Foliar | No | [91] |
| Zucchini | Soil, foliar | Negative | [51] |
| Plant | Disease | Causal Agent | Reference |
|---|---|---|---|
| Apple | Mouldy core | Alternaria alternate | [106] |
| Avacado | Dieback | Phytophthora cinnamomi | [107] |
| Banksia | Dieback | Phytophthora cinnamomi | [108] |
| Bentgrass | Summer decline | Pythium | [73] |
| Cabbage | Clubroot | Plasmodiophora brassicae | [109] |
| Chestnut | Ink disease | Phytophthora cambivora | [110] |
| Cucumber | Damping-off | Pythium ultimum | [111] |
| Grape | Downy mildew | Plasmopara viticola | [112] |
| Lupin | Dieback | Phytophthora cinnamomi | [113] |
| Maize | Downy mildew | Peronosclerospora sorghi | [114] |
| Orange | Brown rot | Phytophthora citrophthora | [53] |
| Papaya | Fruit rot | Phytophthora palmivora | [113] |
| Pecan | Scab | Fusicladium effusum | [115] |
| Pepper | Crown and root rot | Phytophthora capsici | [59] |
| Potato | Late blight | Phytophthora infestans | [116] |
| Potato | Late blight | Phytophthora infestans | [117] |
| Potato | Pink rot | Phytophthora erythroseptica | [118] |
| Potato | Bacterial soft rot | Erwinia carotovora | [119] |
| Soybean | Downey mildew | Peronospora manshurica | [102] |
| Strawberry | Leather rot | Phytophthora cactorum | [120] |
| Tangelo | Brown spot | Alternaria alternata | [121] |
| Tobacco | Black shank | Phytophthora nicotianae | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havlin, J.L.; Schlegel, A.J. Review of Phosphite as a Plant Nutrient and Fungicide. Soil Syst. 2021, 5, 52. https://doi.org/10.3390/soilsystems5030052
Havlin JL, Schlegel AJ. Review of Phosphite as a Plant Nutrient and Fungicide. Soil Systems. 2021; 5(3):52. https://doi.org/10.3390/soilsystems5030052
Chicago/Turabian StyleHavlin, John L., and Alan J. Schlegel. 2021. "Review of Phosphite as a Plant Nutrient and Fungicide" Soil Systems 5, no. 3: 52. https://doi.org/10.3390/soilsystems5030052
APA StyleHavlin, J. L., & Schlegel, A. J. (2021). Review of Phosphite as a Plant Nutrient and Fungicide. Soil Systems, 5(3), 52. https://doi.org/10.3390/soilsystems5030052