
Observations on Earthworm Communities and Soils in Various Natural and Man-Affected Ecosystems

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Sorting of Earthworm Samples
2.3. Soil Properties Analyses
2.4. Statistical Analyses
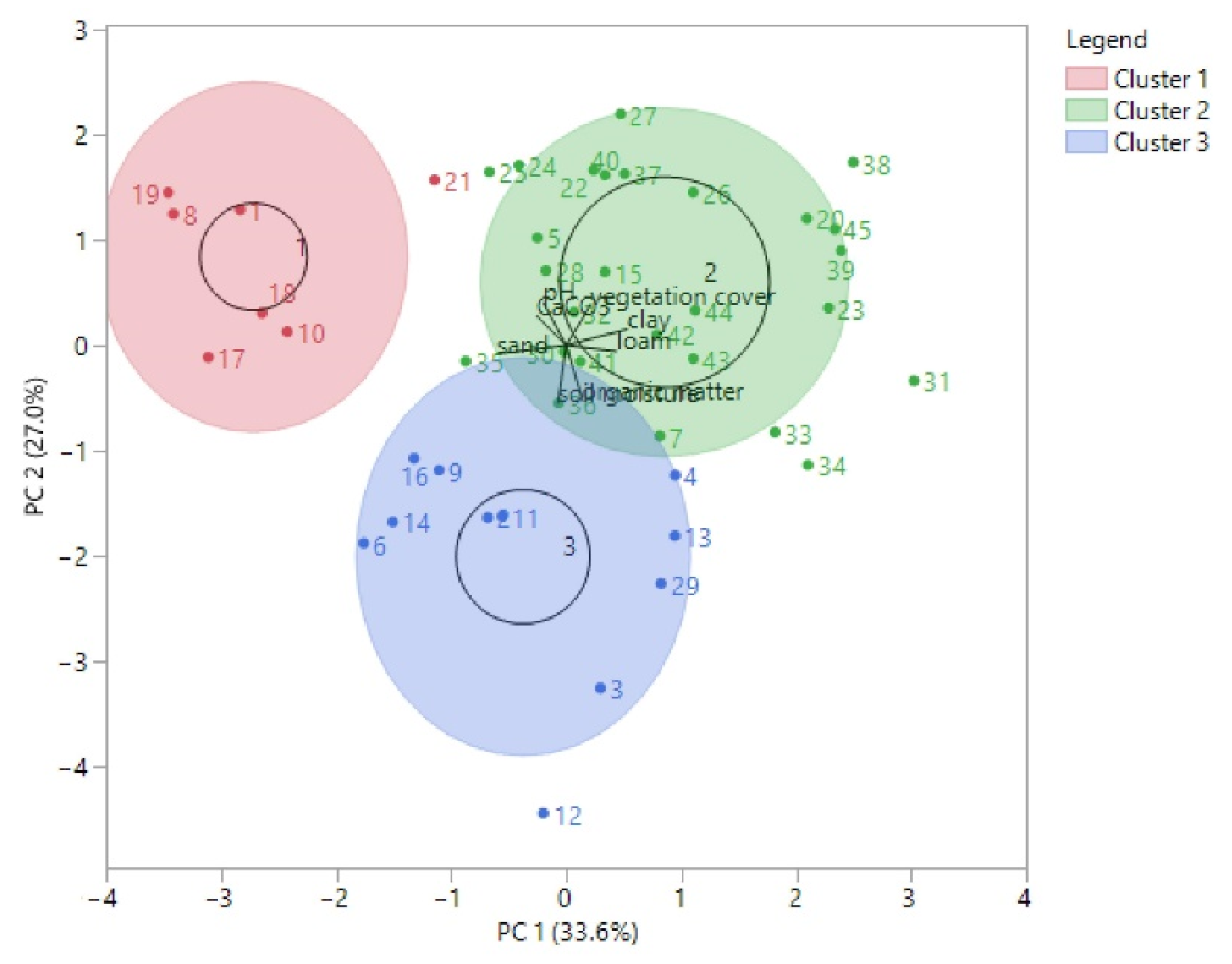
3. Results
3.1. Earthworm Species and Abundance
3.2. Soil Properties
3.3. Vegetation Differences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, C.A.; Bohlen, P.J. Biology and Ecology of Earthworms, 3rd ed.; Chapman & Hall: London, UK, 1998. [Google Scholar]
- Didden, W.A.M. Earthworm communities in grasslands and horticultural soils. Biol. Fertil. Soils 2001, 33, 111–117. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J. Spatial patterns of surface soil properties and vegetation in a Mediterranean semi-arid steppe. Plant Soil 2002, 241, 279–291. [Google Scholar] [CrossRef]
- Wills, A.; Abbott, I. Landscape-scale species richness of earthworms in the Porongurup Range, Western Australia: Influence of aspect, soil fertility, and vegetation type. Biol. Fertil. Soils 2003, 39, 94–102. [Google Scholar] [CrossRef]
- Whalen, J.K. Spatial and temporal distribution of earthworm patches in corn field, hayfield and forest systems of southwestern Quebec, Canada. Appl. Soil Ecol. 2004, 27, 143–151. [Google Scholar] [CrossRef]
- Pavlíček, T.; Csuzdi, C.; Smooha, G.; Beiles, A.; Nevo, E. Biodiversity and microhabitat distribution of earthworms at “Evolution Canyon” a Mediterranean microsite, mount Carmel, Israel. Isr. J. Zool. 1996, 42, 449–454. [Google Scholar] [CrossRef]
- Szlavecz, K.; Placella, S.A.; Pouyat, R.V.; Groffman, P.; Csuzdi, C.; Yesilonis, I. Invasive earthworm species and nitrogen cycling in remnant forest patches. Appl. Soil Ecol. 2006, 32, 54–62. [Google Scholar] [CrossRef]
- Huerta, E.; Rodriguez-Olan, J.; Evia-Castillo, I.; Montejo-Meneses, E.; de la Cruz-Mondragon, M.; Garcia-Hernandez, R.; Uribe-Lopez, S. Earthworms and soil properties in Tabasco, Mexico. Eur. J. Soil Biol. 2007, 43, S190–S195. [Google Scholar] [CrossRef]
- Cole, L.J.; Pollock, M.; Robertson, D.; Holland, J.P.; McCracken, D.; Harrison, W. The influence of fine-scale habitat heterogeneity on invertebrate assemblage structure in upland semi-natural grassland. Agric. Ecosyst. Environ. 2010, 136, 69–80. [Google Scholar] [CrossRef]
- Ermakov, A.I.; Golovanova, E. Species composition and abundance of earthworms in the tundra biocenoses of Denezhkin Kamen’ Mountain (Northern Urals). Contemp. Probl. Ecol. 2010, 3, 10–14. [Google Scholar] [CrossRef]
- Salomé, C.; Guenat, C.; Bullinger-Weber, G.; Gobat, J.-M.; Le Bayon, C. Earthworm communities in alluvial forests: Influence of altitude, vegetation stages and soil parameters. Pedobiologia 2011, 54, S89–S98. [Google Scholar] [CrossRef]
- Curry, J. Factors Affecting the Abundance of Earthworms in Soils. In Earthworm Ecology, 2nd ed.; Routledge: London, UK, 2004; pp. 91–113. [Google Scholar] [CrossRef]
- Phillips, H.R.P.; Bach, E.M.; Bartz, M.L.C.; Bennett, J.M.; Beugnon, R.; Briones, M.J.I.; Brown, G.G.; Ferlian, O.; Gongalsky, K.B.; Guerra, C.A.; et al. Global data on earthworm abundance, biomass, diversity and corresponding environmental properties. Sci. Data 2021, 8, 136. [Google Scholar] [CrossRef] [PubMed]
- Hoeffner, K.; Santonja, M.; Monard, C.; Barbe, L.; LE Moing, M.; Cluzeau, D. Soil properties, grassland management, and landscape diversity drive the assembly of earthworm communities in temperate grasslands. Pedosphere 2021, 31, 375–383. [Google Scholar] [CrossRef]
- Prendergast-Miller, M.T.; Jones, D.T.; Berdeni, D.; Bird, S.; Chapman, P.J.; Firbank, L.; Grayson, R.; Helgason, T.; Holden, J.; Lappage, M.; et al. Arable fields as potential reservoirs of biodiversity: Earthworm populations increase in new leys. Sci. Total Environ. 2021, 789, 147880. [Google Scholar] [CrossRef]
- Vršič, S.; Breznik, M.; Pulko, B.; Rodrigo-Comino, J. Earthworm Abundance Changes Depending on Soil Management Practices in Slovenian Vineyards. Agronomy 2021, 11, 1241. [Google Scholar] [CrossRef]
- Tzelepis, N.D. Contribution to the Study of the Oligochaeta of Greece. Ph.D. Thesis, Kapodistrian University of Athens, Athens, Greece, 1943. [Google Scholar]
- Zicsi, A.; Michalis, K. Übersicht der regenwurm-fauna Griechenlands (Oligochaeta: Lumbricidae). Acta Zool. Hung. 1981, XXVII, 239–264. [Google Scholar]
- Zicsi, A.; Michalis, K. Zwei Neue Dendrobaena—Arten aus Griechenland (Oligochaeta, Lumbricidae). Acta Zool. Hung 1993, 39, 301–310. [Google Scholar]
- Szederjesi, T.; Csuzdi, C. New and little known earthworm species from Greece (Oligochaeta: Lumbricidae, Acanthodrilidae). Zootaxa 2012, 3304, 25–42. [Google Scholar] [CrossRef]
- Szederjesi, T.; Angyal, D.; Balázs, G.; Dányi, L. Remarks on the earthworm genus Helodrilus Hoffmeister, 1845 with new epigean and subterranean records (Oligochaeta: Lumbricidae). Opusc. Zool. Bp. 2014, 45, 181–188. [Google Scholar]
- Szederjesi, T.; Vavoulidou, E.; Chalkia, C.; Dányi, L.; Csuzdi, C. An annotated checklist of earthworms of Greece (Clitellata: Megadrili). Zootaxa 2017, 4272, 57–82. [Google Scholar] [CrossRef]
- Michalis, K.; Fragoulis, A.N.; Panidis, S. Eathworms (oligochaeta: Lumbricidae) from the mountainous mas of Pella province southwest region, and Imathias province, northwest region a systematical and ecological study. Ecol. Mediterr. 1985, 11, 37–42. [Google Scholar] [CrossRef]
- Michalis, K.B. Conrtibution to the systematic and ecological study of the earthworms (Oligochaeta: Lumbricidae) of the Western Thrace. Arq. Mus. Bocage Nova Ser. 1987, 1, 59–69. [Google Scholar]
- Stojanović, M.; Sekulić, J.; Trakić, T. Distribution and threat status of the endemic earthworm Allolobophora dofleini (Ologochaeta: Lumbricidae) on the Balkan Peninsula. North-West J. Zool. 2017, 13, 136–143. [Google Scholar]
- Solomou, A.D.; Sfougaris, A.I.; Vavoulidou, E.M.; Csuzdi, C. The effects of farming practices on earthworm dynamics in olive groves of central Greece. Zool. Middle East 2012, 58, 119–126. [Google Scholar] [CrossRef]
- Solomou, A.D.; Sfougaris, A.I.; Vavoulidou, E.M.; Csuzdi, C. Species Richness and Density of Earthworms in Relation to Soil Factors in Olive Orchard Production Systems in Central Greece. Commun. Soil Sci. Plant Anal. 2013, 44, 301–311. [Google Scholar] [CrossRef]
- Solomou, A.D.; Sfougaris, A. Contribution of Agro-Environmental Factors to Yield and Plant Diversity of Olive Grove Ecosystems (Olea europaea L.) in the Mediterranean Landscape. Agronomy 2021, 11, 161. [Google Scholar] [CrossRef]
- Vavoulidou, E.; Benckiser, G.; Kavvadias, V.A. Soil Fauna Activities in Agricultural Greek Landscapes. In Soil and Recycling Management in the Anthropocene Era; Benckiser, G., Ed.; Springer Nature: Cham, Switzerland, 2021; pp. 87–113. [Google Scholar]
- Vavoulidou, E.; Bilalis, D.; Chalkia, C. Chapter 7. The earthworms. In The Fauna of Greece. Biology & Management of the Wild Fauna; Pafilis, P., Ed.; Broken Hill Publishers Ltd.: Nicosia, Cyprus, 2020; pp. 199–226. [Google Scholar]
- Michalis, K. Criodrilus lacuum HOFFMEISTER, 1845 (Oligochaeta: Criodrilidae)—Earthworm species new to Greece. Acta Zool. Cracov. 1993, 36, 15–21. [Google Scholar]
- Csuzdi, C.; Zicsi, A. Earthworms of Hungary (Annelida: Oligochaeta, Lumbricidae); Hungarian Natural History Museum & Sys-tematic Zoology Research Group of the Hungarian Academy of Sciences: Budapestt, Hungary, 2003; p. 271. [Google Scholar]
- Sherlock, E. Key to the Earthworms of the UK and Ireland; FSC Publications: London, UK, 2012; p. 44. [Google Scholar]
- Szederjesi, T.; Pop, V.V.; Pavlíček, T.; Márton, O.; Krízsik, V.; Csuzdi, C. Integrated taxonomy reveals multiple species in the Dendrobaena byblica (Rosa, 1893) complex (Oligochaeta: Lumbricidae). Zool. J. Linn. Soc. 2018, 182, 500–516. [Google Scholar] [CrossRef]
- Available online: www.gbif.org/species/2307609 (accessed on 10 September 2021).
- Gasparatos, D.; Kabasilis, S.; Giannakopoulou, F. Exercises of General Soil Science; A.U.A. Press: Athens, Greece, 2011. [Google Scholar]
- Benton, J., Jr. Laboratory Guide for Conducting Soil Tests and Plant Analysis; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- 4SAS Institute. JMP Version 10.0.0; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 October 2021).
- Baldivieso-Freitas, P.; Blanco-Moreno, J.M.; López, M.G.; Peigné, J.; Pérez-Ferrer, A.; Trigo-Aza, D.; Sans, F.X. Earthworm abundance response to conservation agriculture practices in organic arable farming under Mediterranean climate. Pedobiologia 2018, 66, 58–64. [Google Scholar] [CrossRef]
- Schon, N.S.; Mackay, A.D.; Gray, R.A. Changes in the abundance and diversity of earthworms in hill soils under different long-term fertiliser and sheep stocking regimes. N. Z. J. Agric. Res. 2021, 64, 162–177. [Google Scholar] [CrossRef]
- Feijoo, A.; Carvajal, A.F.; Zúñiga, M.C.; Quintero, H.; Fragoso, C. Diversity and abundance of earthworms in land use systems in central-western Colombia. Pedobiologia 2011, 54, S69–S75. [Google Scholar] [CrossRef]
- Kherbouche, D.; Bernhard-Reversat, F.; Moali, A.; Lavelle, P. The effect of crops and farming practices on earthworm communities in Soummam valley, Algeria. Eur. J. Soil Biol. 2012, 48, 17–23. [Google Scholar] [CrossRef]
- Tsiafouli, M.A.; Thébault, E.; Sgardelis, S.P.; De Ruiter, P.C.; van der Putten, W.; Birkhofer, K.; Hemerik, L.; de Vries, F.; Bardgett, R.D.; Brady, M.V.; et al. Intensive agriculture reduces soil biodiversity across Europe. Glob. Chang. Biol. 2014, 21, 973–985. [Google Scholar] [CrossRef]
- Domínguez, A.; Bedano, J.C. Earthworm and Enchytraeid Co-occurrence Pattern in Organic and Conventional Farming. Soil Sci. 2016, 181, 148–156. [Google Scholar] [CrossRef]
- Lindahl, A.M.L.; Dubus, I.G.; Jarvis, N.J. Site Classification to Predict the Abundance of the Deep-Borrowing Earthworm Lumbricus terrestris L. Vadoze Zone J. 2009, 8, 911–915. [Google Scholar] [CrossRef]
- Pfiffner, L.; Luka, H. Earthworm populations in two low-input cereal farming systems. Appl. Soil Ecol. 2007, 37, 184–191. [Google Scholar] [CrossRef]
- Castro, J.; Barreal, M.E.; Briones, M.J.; Gallego, P.P. Earthworm communities in conventional and organic fruit orchards under two different climates. Appl. Soil Ecol. 2019, 144, 83–91. [Google Scholar] [CrossRef]
- Meissner, G.; Athmann, M.E.; Fritz, J.; Kauer, R.; Stoll, M.; Schultz, H.R. Conversion to organic and biodynamic viticultural practices: Impact on soil, grapevine development and grape quality. OENO One 2019, 53, 639–659. [Google Scholar] [CrossRef]
- Bauer, R.; Küpper, K.; Müller, H.W. 1998. Characterization of the Lumbricid Fauna in Alluvial Soils in the Danube River Floodplain Area East of Vienna. Linzer. Boil. Beitr. 1998, 30, 11–20. Available online: https://www.zobodat.at/pdf/LBB_0030_1_0011-0020.pdf (accessed on 7 December 2021).
- Leon, Y.S.-D.; Zou, X.; Borges, S.; Ruan, H. Recovery of Native Earthworms in Abandoned Tropical Pastures. Conserv. Biol. 2003, 17, 999–1006. [Google Scholar] [CrossRef]
- Chandran, M.S.S.; Sujatha, S.; Mohan, M.; Julka, J.M.; Ramasamy, E.V. Earthworm diversity at Nilgiri biosphere reserve, Western Ghats, India. Biodivers. Conserv. 2012, 21, 3343–3353. [Google Scholar] [CrossRef]
- Robinson, C.H.; Piearce, T.G.; Ineson, P.; Dickson, D.A.; Nys, C. Earthworm communities of limed coniferous soils: Field observations and implications for forest management. For. Ecol. Manag. 1992, 55, 117–134. [Google Scholar] [CrossRef]
- Plum, N.M.; Filser, J. Floods and drought: Response of earthworms and potworms (Oligochaeta: Lumbricidae, Enchytraeidae) to hydrological extremes in wet grassland. Pedobiologia 2005, 49, 443–453. [Google Scholar] [CrossRef]
- Ivask, M.; Meriste, M.; Kuu, A.; Kutti, S.; Sizov, E. Effect of flooding by fresh and brackish water on earthworm communities along Matsalu Bay and the Kasari River. Eur. J. Soil Biol. 2012, 53, 11–15. [Google Scholar] [CrossRef]
- Pop, V.V. Earthworm-vegetation-soil relationships in the Romanian Carpathians. Soil Biol. Biochem. 1997, 29, 223–229. [Google Scholar] [CrossRef]
- Decaëns, T.; Margerie, P.; Aubert, M.; Hedde, M.; Bureau, F. Assembly rules within earthworm communities in North-Western France—A regional analysis. Appl. Soil Ecol. 2008, 39, 321–335. [Google Scholar] [CrossRef]
- Mathieu, J.; Grimaldi, M.; Jouquet, P.; Rouland, C.; Lavelle, P.; Desjardins, T.; Rossi, J.-P. Spatial patterns of grasses influence soil macrofauna biodiversity in Amazonian pastures. Soil Biol. Biochem. 2009, 41, 586–593. [Google Scholar] [CrossRef]
- Korboulewsky, N.; Perez, G.; Chauvat, M. How tree diversity affects soil fauna diversity: A review. Soil Biol. Biochem. 2016, 94, 94–106. [Google Scholar] [CrossRef]
- Homan, C.; Beier, C.; McCay, T.; Lawrence, G. Application of lime (CaCO3) to promote forest recovery from severe acidification increases potential for earthworm invasion. For. Ecol. Manag. 2016, 368, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Angst, Š.; Mueller, C.W.; Cajthaml, T.; Angst, G.; Lhotáková, Z.; Bartuška, M.; Špaldoňová, A.; Frouz, J. Stabilization of soil organic matter by earthworms is connected with physical protection rather than with chemical changes of organic matter. Geoderma 2017, 289, 29–35. [Google Scholar] [CrossRef]
- Hendrix, P.; Mueller, B.; Bruce, R.; Langdale, G.; Parmelee, R. Abundance and distribution of earthworms in relation to landscape factors on the Georgia Piedmont, USA. Soil Biol. Biochem. 1992, 24, 1357–1361. [Google Scholar] [CrossRef]
- Hughes, M.S.; Bull, C.M.; Doube, B.M. The use of resource patches by earthworms. Biol. Fertil. Soils 1994, 18, 241–244. [Google Scholar] [CrossRef]
- Ashwood, F.; Butt, K.R.; Doick, K.J.; Vanguelova, E.I. Investigating tree foliar preference by the earthworms Aporrectodea longa and Allolobophora chlorotica in reclaimed and loam soil. Appl. Soil Ecol. 2017, 110, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Ammer, S.; Weber, K.; Abs, C.; Ammer, C.; Prietzel, J. Factors influencing the distribution and abundance of earthworm communities in pure and converted Scots pine stands. Appl. Soil Ecol. 2006, 33, 10–21. [Google Scholar] [CrossRef]
- 6Jacob, M.; Weland, N.; Platner, C.; Schaefer, M.; Leuschner, C.; Thomas, F.M. Nutrient release from decomposing leaf litter of temperate deciduous forest trees along a gradient of increasing tree species diversity. Soil Biol. Biochem. 2009, 41, 2122–2130. [Google Scholar] [CrossRef]
- Crumsey, J.M.; Le Moine, J.M.; Vogel, C.S.; Nadelhoffer, K. Historical patterns of exotic earthworm distributions inform contemporary associations with soil physical and chemical factors across a northern temperate forest. Soil Biol. Biochem. 2014, 68, 503–514. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a/a | Site (Sampling Date) | Type | Species | Total | ||||
|---|---|---|---|---|---|---|---|---|
| Terrestrial Water Bodies | ||||||||
| 1 | Menidi river, W Attiki (2–8–2015) | river bank | Ap. c. caliginosa 44.8 ± 25.5 | B. r.-subrubicundus 6.4 ± 6.4 | Eis. fetida 9.6 ± 6.4 | Dendr. veneta 35.2 ± 11.8 | 96 ± 46.1 | |
| 2 | Karakanta Penteli Penteli mtn (21–8–2015) | mountain spring | Ap. c. caliginosa 0.8 ± 0.8 | Ap. c.–trapezoides 3.2 ± 2 | O. complanatus 4 ± 2.5 | Eis. tetraedra 5.6 ± 3.5 | Dend. b.-byblica 16 ± 9.6 | 31.2 ± 15 |
| unidentified 1.6 ± 1.6 | ||||||||
| 3 | Mesiano Nero Parnitha mtn (23–8–2015) | mountain spring | Ap. rosea 4 ± 3.1 | Eis. tetraedra 4 ± 3.1 | Dendr.b.-byblica 12 ± 5.9 | 20 ± 5.9 | ||
| 4 | Panaktos –Oinoi North Attiki (1–9–2015) | mountain spring | Dendr. b.-byblica 3.2 ± 3.2 | unidentified 3.2 ± 3.2 | 6,4 ± 3.9 | |||
| 5 | Filis spring, N. Attiki (13–9–2015) | mountain spring | Ap. c.- trapezoides 86.4 ± 36 | Eis. tetraedra 3.2 ± 2 | Dendr. b.-byblica 84.8 ± 45.9 | 177.2 ± 79.3 | ||
| 6 | Kryfo Nero, Parnitha mtn (28–10–2015) | mountain spring | Ap. rosea 12 ± 5.9 | O. complanatus 4.8 ± 2.3 | Eis. tetraedra 3.2 ± 2 | B. r.-rubidus 1.6 ± 1 | Dendr. b.-byblica 4 ± 3.1 | 27.2 ± 7.9 |
| unidentified 1.6 ± 1.6 | ||||||||
| 7 | Paliochori., Parnitha mtn (14–11–2015) | mountain spring | Ap. rosea 2.4 ± 2.4 | O. complanatus 11.2 ± 3.2 | Eis. fetida 0.8 ± 0.8 | L. rubellus 14.4 ± 4.8 | Eis. tetraedra 18.4 ± 16.5 | 47.2 ± 13.1 |
| 8 | Pykrodafnis stream Central Attiki (3–4–2016) | river bank | Eis. tetraedra 191.2 ± 129.6 | Β. r.-subrubicundus 100.8 ± 79.1 | 292 ± 208.4 | |||
| 9 | Agia Triada Parnitha mtn (7–5–2016) | mountain spring | Ap. c. caliginosa 6.4 ± 3 | Ap. c.- trapezoides 5.6 ± 4.7 | O. complanatus 2.4 ± 1.6 | Eis. fetida 0.8 ± 0.8 | All. chlorotica 4.8 ± 3.9 | 44.8 ± 19.2 |
| Oct. lacteum 2.4 ± 2.4 | Micr. phosphoreus 11.2 ± 6.8 | Euk. saltensis 6.4 ± 4.1 | ||||||
| 10 | Pachi stream., W. Attiki (4–6–2016) | stream bank | Ap. rosea 8 ± 4.4 | Ap. c.- trapezoides 22.4 ± 13.1 | Eis. fetida 57.6 ± 36.8 | Eis. tetraedra 47.2 ± 21.4 | Β. r.-subrubicundus 3.2 ± 2 | 244.8 ± 87.6 |
| Dendr. veneta 3.2 ± 1.5 | Euk. saltensis 78.4 ± 31.7 | Ocn. occidentalis 2.4 ± 1.6 | unidentified 20 ± 19 | |||||
| 11 | Asopos, Corinth (20–6–2016) | river bank | Ap. rosea 2.4 ± 1.6 | Eis. tetraedra 3.2 ± 2.3 | D. byblica-byblica 9.6 ± 5.5 | Cr. lacuum 14.4 ± 13.4 | 29.6 ± 15.2 | |
| 12 | Doxa lake, Corinth (13–8–2016 | lake | Eis. tetraedra 9.6 ± 7.8 | Dendr. b.-byblica 7.2 ± 7.2 | Cr. lacuum √ | 16.8 ± 14.9 | ||
| 13 | Koromilia, Parnitha mtn (4–9–2016) | mountain spring | Ap. rosea 7.2 ± 7.2 | Ap. c.- trapezoides 0.8 ± 0.8 | 11.2 ± 7.5 | |||
| 14 | Kyra, Parnitha mtn (2–10–2016) | mountain spring | Dendr. b.-byblica 12 ± 4.2 | 12 ± 4.2 | ||||
| 15 | Boeotic Kifissos, Boeotia (5–10–2016) | estuary | Ap. rosea 32.8 ± 30.8 | 32.8 ± 30.8 | ||||
| 16 | Viliani, Parnitha mtn (21–7–2017) | mountain spring | O. complanatus 6.4 ± 2.4 | Eis. fetida 0.8 ± 0.8 | 7.2 ± 2.9 | |||
| 17 | Kourna lake, Crete (30–8–2017) | lake | 0 | |||||
| 18 | Mpeletsi, Parnitha mtn (5–11–2017) | artificial lake | Ap. c.- trapezoides 4 ± 2.5 | O. complanatus 3.2 ± 3.2 | L. rubellus 0.8 ± 0.8 | Ap. rosea 0.8 ± 0.8 | 8.8 ± 5.4 | |
| 19 | Spata stream, N. Attiki (2–8–18) | river | Ap. c.- trapezoides 0.8 ± 0.8 | Eis. fetida 0.8 ± 0.8 | Eis. tetraedra 16.8 ± 16.8 | Dend. b.-byblica 4.8 ± 4.8 | 23.2 ± 23.2 | |
| Undisturbed Sites | Type | Species | Total | |||||
| 20 | Menidi road,.W. Attiki (9–8–2015) | edge of street (H2O-logged) | Ap. rosea 19.2 ± 9.3 | Ap. c. caliginosa 6.4 ± 6.4 | Ap. c.- trapezoides 34 ± 26.7 | O. complanatus 6.4 ± 3.9 | 66 ± 36.5 | |
| 21 | Nea Makri, N. Attiki (27–9–2015) | field | Ap. rosea 0.8 ± 0.8 | Ap. c.- trapezoides 5.6 ± 2.5 | O. complanatus 12.8 ± 8.7 | 19.2 ± 9.8 | ||
| 22 | Eucalyptus plantation outskirts of Athens (25–10–2015) | forest plantation | Ap. rosea 12.8 ± 7.9 | Ap. c. caliginosa 4 ± 3.1 | Ocn. occidentalis 0.8 ± 0.8 | 17.6 ± 7.4 | ||
| 23 | Faskomilia, S. Athens (2–11–2015) | maquis | Ap. rosea 3.2 ± 3.2 | All. dofleini √ | 3.2 ± 3.2 | |||
| 24 | Dionysos, N. Attiki (21–11–2015) | coniferous & maquis | O. complanatus 2.4 ± 1.6 | L. rubellus 3.2 ± 2.3 | Dendr. olympiaca 1.6 ± 1 | 7.2 ± 4.5 | ||
| 25 | Ano Glyfada, S. Athens (6–12–2015) | maquis | Ap. rosea 5.6 ± 3.7 | 5.6 ± 3.7 | ||||
| 26 | Parnitha maquis (26–12–2015) | maquis | O. complanatus 0.8 ± 0.8 | unidentified 2.4 ± 2.4 | 3.2 ± 2.3 | |||
| 27 | Parnitha pineforest (27–12–2015) | coniferous forest | O. complanatus 1.6 ± 1 | Dendr. attemsi | 1.6 ± 1 | |||
| 28 | Veikou grove, Athens (1–1–2016) | grove | Ap. rosea 92.8 ± 54.3 | Ap. c. caliginosa 0.8 ± 0.8 | Ap. c.- trapezoides 2.4 ± 2.4 | O. complanatus 2.4 ± 1 | Micr.dubius 5.6 ± 5.6 | 104 ± 62.5 |
| 29 | Kamena Parnitha (9–1–2016) | burned forest | Ap. rosea 1.6 ± 1.6 | O. complanatus 3.2 ± 0.8 | 3.2 ± 0.8 | |||
| 30 | Kavouri, S. Athens (17–1–2016) | pineforest & sea bank | Ap. rosea 109.6 ± 106.6 | 109.6±106.6 | ||||
| 31 | Tatoi, North Attiki (14–2–2016 | coniferous & maquis | Ap. rosea 8 ± 8 | Ap. c. caliginosa 2.4 ± 2.4 | O. complanatus 8.8 ± 4.1 | unidentified 0.8 ± 0.8 | 20 ± 8 | |
| 32 | Mola, Parnitha mtn (23–3–2016) | alpine meadow | O. complanatus 18.4 ± 8.8 | Mur. minuscula 1.6 ± 1.6 | 20 ± 8 | |||
| 33 | Mola-Parnitha (13–4–2017) | alpine meadow | Ap. rosea 41.6 ± 24.7 | O. complanatus 4 ± 2.2 | Micr.phosphoreus | 45.6 ± 24 | ||
| 34 | Stymfalia road, Corinth (7–8–2016) | road alley | Ap. rosea 27.2 ± 17 | O. complanatus 8 ± 3.3 | 35.2 ± 18.9 | |||
| 35 | Poros island, pineforest (3–12–2016) | coniferous forest | 0 | |||||
| Cultivated Fields | Type | Species | Total | |||||
| 36 | Syngrou field, N. Athens (4–10–2015) | garden | Ap. c. caliginosa 0.8 ± 0.8 | O. complanatus 9.6 ± 6.8 | L. rubellus 2.4 ± 2.4 | 12.8 ± 9.9 | ||
| 37 | Lawn in AUA West Athens (30–6–2016) | public garden | Ap. rosea 367.2 ± 51.5 | Ap. c. caliginosa 80.8 ± 31.9 | Ap. c.- trapezoides 80.8 ± 31.9 | O. complanatus 0.8 ± 0.8 | Eis. fetida 0.8 ± 0.8 | 673.6 ± 158.7 |
| Euk. saltensis 2.4 ± 2.4 | A. chlorotica 53.6 ± 47.8 | Am. gracilis 2.4 ± 2.4 | Unidentified sp.1 134.4 ± 56.9 | Unidentified sp.2 8.8 ± 8.8 | ||||
| 38 | Vineyard, Corinth (24–9–2016) | vineyard | Ap. rosea 7.2 ± 7.2 | Apporectodea spp. 3.2 ± 3.2 | Dendr. olympiaca 0.8 ± 0.8 | 11.2 ± 10.2 | ||
| 39 | Corinthia highland Corinth (30–10–2016) | fallow field | Ap. rosea 24 ± 13.1 | Ap. c. caliginosa 4.8 ± 4.8 | O. complanatus 5.6 ± 3 | L. rubellus 3.2 ± 2.3 | Dendr. olympiaca 0.8 ± 0.8 | 48.8 ± 22.2 |
| Micr.phosphoreus | Micr. dubius 4.8 ± 2.9 | Ocn. occidentalis 5.6 ± 5.6 | ||||||
| 40 | Marousi, N. Athens (13–11–2016) | garden | Ap. rosea 2.4 ± 1.6 | Ap. c.- trapezoides 0.8 ± 0.8 | O. complanatus 18.4 ± 6.4 | L. rubellus 1.6 ± 1.6 | 23.2 ± 7.9 | |
| 41 | Poros island, lowlands (3–12–2016) | plains | Ap. rosea 8.8 ± 8.8 | Ap. c. caliginosa 1.6 ± 1.6 | Ap. c.- trapezoides 2.4 ± 1.6 | O. complanatus 28.8 ± 12 | L. rubellus 4 ± 2.5 | 45.6 ± 23.4 |
| 42 | Troizina citrus East Peloponnese (4–12–2016) | citrus orchard | Ap. rosea 3.2 ± 2 | Ap. c. caliginosa 0.8 ± 0.8 | Ap. c.- trapezoides 6.4 ± 3.7 | O. complanatus 3.2 ± 2 | 13.6 ± 7.1 | |
| 43 | Troizina olives East Peloponnese (4–12–2016) | olive orchard | Ap. rosea 0.8 ± 0.8 | Ap. c. caliginosa 1.6 ± 1.6 | Ap. c.- trapezoides 2.4 ± 1.6 | O. complanatus 12 ± 11 | 16.8 ± 12.2 | |
| 44 | Marathon, N. Attiki (22–1–2017) | organic vegetables | Ap. rosea 9.6 ± 6 | Ap. c. caliginosa 30.4 ± 22.7 | Ap. c.- trapezoides 15.2 ± 11.6 | O. complanatus 2.4 ± 1.1 | Eis. tetraedra 1.6 ± 1 | 119.2 ± 24.3 |
| Micr. phosphoreus 31.2 ± 11.6 | Micr. dubius 25.6 ± 15.4 | unidentified 3.2 ± 2.3 | ||||||
| 45 | Corinthia highland (22–10–2017) | bushfield | Ap. caliginosa 1.6 ± 1.6 | O. complanatus 2.4 ± 2.4 | L. rubellus 0.8 ± 0.8 | Ocn. occidentalis 1.6 ± 1 | unidentified 1.6 ± 1 | 8 ± 6.2 |
| Parameter | Water Bodies | Undisturbed Sites | Cultivated Fields |
|---|---|---|---|
| soil moisture (%) | (29 ± 3) α | (17 ± 1) β | (14 ± 1) β |
| sand (%) | (64 ± 3) α | (48 ±3) β | (43 ± 3) β |
| loam (%) | (22.5 ± 2) α | (29 ± 2) β | (28 ± 2) α,β |
| clay (%) | (13 ± 2) α | (24 ± 2) β | (29 ± 3) β |
| pH | (7.4 ± 0.1) α | (7.4 ± 0.1) α | (7.4 ± 0.1) α |
| CaCO3 (%) | (24.2 ± 4.5) α | (22.0 ± 4.9) α | (14.3 ± 6.2) α |
| organic matter (%) | (6.0 ± 0.8) α | (5.4 ± 0.7) α | (4.8 ± 0.9) α |
| vegetation cover (%) | (29 ± 8) α | (61 ± 8) β | (61 ± 10) β |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chalkia, C.; Vavoulidou, E.; Csuzdi, C.; Emmanouil, C.; Dritsoulas, A.; Katsileros, A. Observations on Earthworm Communities and Soils in Various Natural and Man-Affected Ecosystems. Soil Syst. 2021, 5, 71. https://doi.org/10.3390/soilsystems5040071
Chalkia C, Vavoulidou E, Csuzdi C, Emmanouil C, Dritsoulas A, Katsileros A. Observations on Earthworm Communities and Soils in Various Natural and Man-Affected Ecosystems. Soil Systems. 2021; 5(4):71. https://doi.org/10.3390/soilsystems5040071
Chicago/Turabian StyleChalkia, Christina, Evangelia Vavoulidou, Csaba Csuzdi, Christina Emmanouil, Alexandros Dritsoulas, and Anastasios Katsileros. 2021. "Observations on Earthworm Communities and Soils in Various Natural and Man-Affected Ecosystems" Soil Systems 5, no. 4: 71. https://doi.org/10.3390/soilsystems5040071
APA StyleChalkia, C., Vavoulidou, E., Csuzdi, C., Emmanouil, C., Dritsoulas, A., & Katsileros, A. (2021). Observations on Earthworm Communities and Soils in Various Natural and Man-Affected Ecosystems. Soil Systems, 5(4), 71. https://doi.org/10.3390/soilsystems5040071