
Home-Field Advantage of Litter Decomposition Faded 8 Years after Spruce Forest Clearcutting in Western Germany


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Litter Decomposition Experiments
2.3. Data Statistics
3. Results
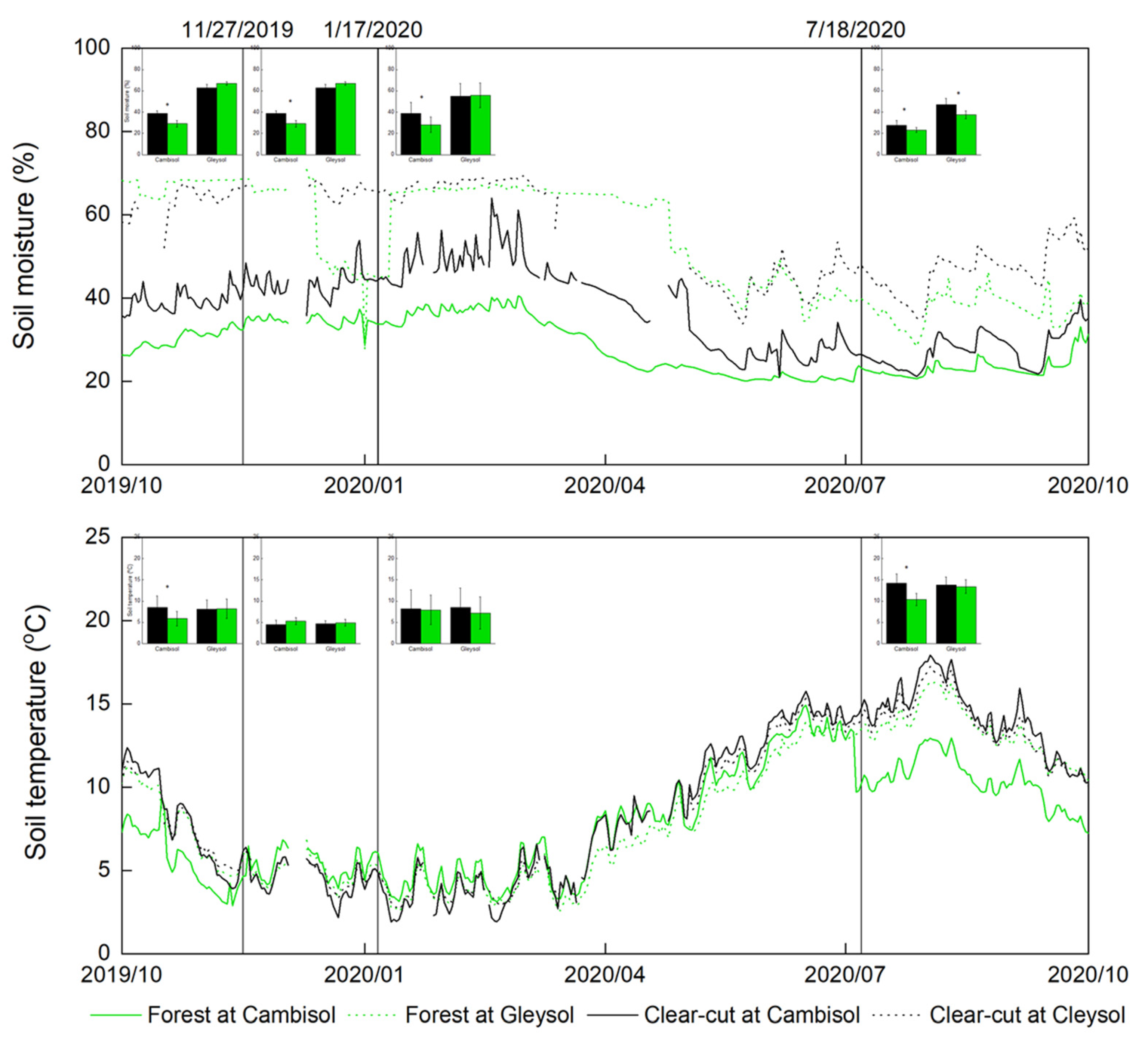
3.1. Environmental Difference between Stands and Soil
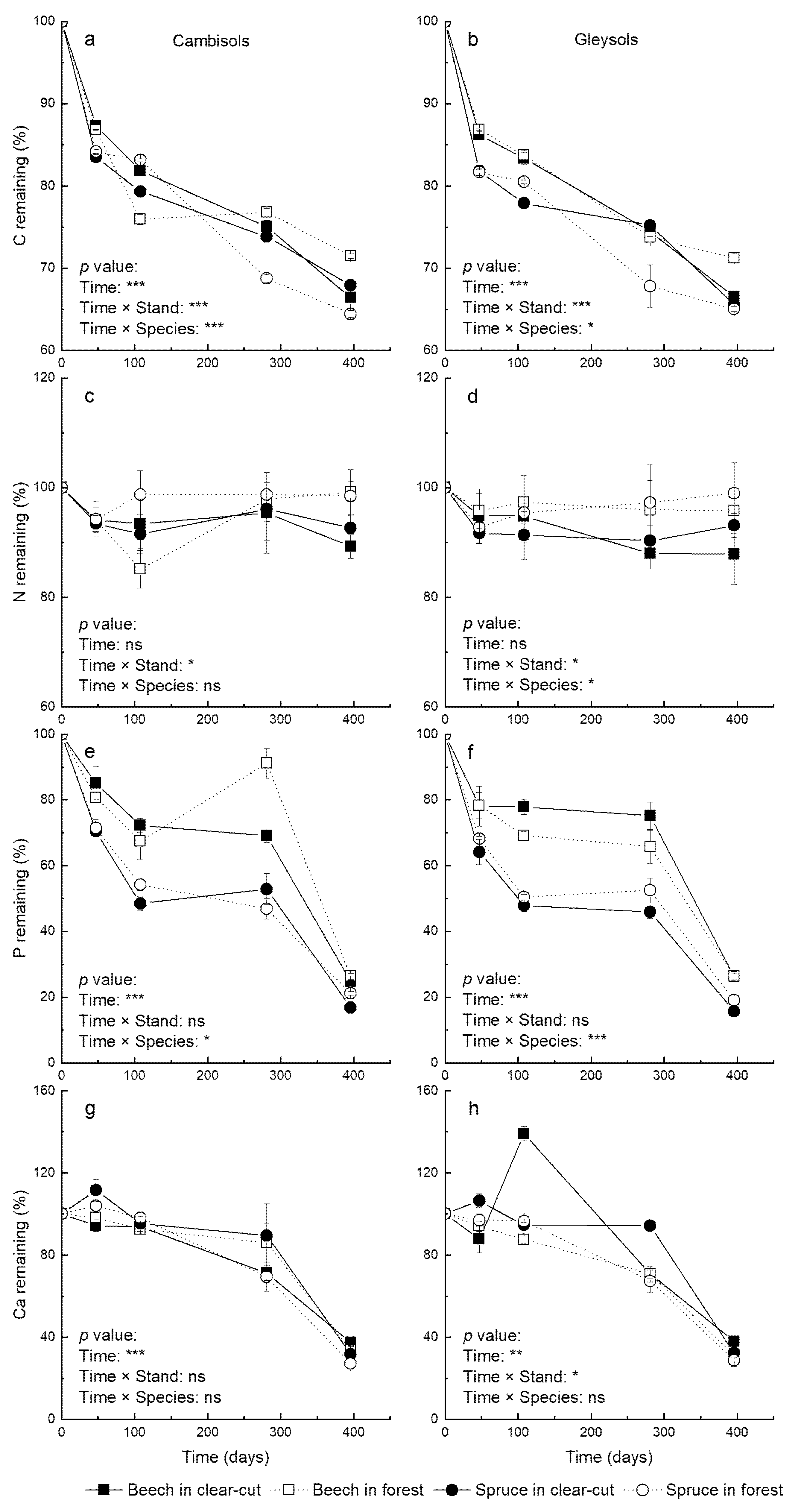
3.2. Initial Litter Quality and Litter Nutrients after One Year of Decomposition
3.3. The Effect of Home-Field Advantage on Litter Decomposition Rates
3.4. C and N Dynamics and Their HFA
3.5. The Dynamic of Litter Nutrients Release during Decomposition
3.6. Correlation between Litter Mass Loss Rate and Residual Quality
3.7. Isotopic Change during Decomposition
4. Discussion
4.1. HFA in Forest and Post-Harvest Decomposition
4.2. Litter Chemistry Regulated Decomposition of Norway Spruce in Original Forest
4.3. Clearcutting Promoted Beech Decomposition and Nutrient Release Pattern
4.4. Soil Moisture as a Mediator of Litter Decomposition and HFA
4.5. Dynamics of the Natural Abundance of δ13C and δ15N during Decomposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Farber, S.; Costanza, R.; Childers, D.L.; Erickson, J.; Gross, K.; Grove, M.; Hopkinson, C.S.; Kahn, J.; Pincetl, S.; Troy, A.; et al. Linking Ecology and Economics for Ecosystem Management. BioScience 2006, 56, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Both, S.; Elias, D.M.; Kritzler, U.H.; Ostle, N.J.; Johnson, D. Land use not litter quality is a stronger driver of decomposition in hyperdiverse tropical forest. Ecol. Evol. 2017, 7, 9307–9318. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Prescott, C.E.; Abaker, W.E.; Augusto, L.; Cécillon, L.; Ferreira, G.W.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.-P. Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. For. Ecol. Manag. 2020, 466, 118127. [Google Scholar] [CrossRef]
- Achilles, F.; Tischer, A.; Bernhardt-Romermann, M.; Heinze, M.; Reinhardt, F.; Makeschin, F.; Michalzik, B. European beech leads to more bioactive humus forms but stronger mineral soil acidification as Norway spruce and Scots pine—Results of a repeated site assessment after 63 and 82 years of forest conversion in Central Germany. For. Ecol. Manag. 2021, 483, 15. [Google Scholar] [CrossRef]
- Trap, J.; Bureau, F.; Brethes, A.; Jabiol, B.; Ponge, J.-F.; Chauvat, M.; Decaens, T.; Aubert, M. Does moder development along a pure beech (Fagus sylvatica L.) chronosequence result from changes in litter production or in decomposition rates? Soil Biol. Biochem. 2011, 43, 1490–1497. [Google Scholar] [CrossRef] [Green Version]
- Xuluc-Tolosa, F.J.; Vester, H.F.M.; Ramı, X.; Rez-Marcial, N.; Castellanos-Albores, J.; Lawrence, D. Leaf litter decomposition of tree species in three successional phases of tropical dry secondary forest in Campeche, Mexico. For. Ecol. Manag. 2003, 174, 401–412. [Google Scholar] [CrossRef]
- Zhang, W.; Gao, D.X.; Chen, Z.X.; Li, H.; Deng, J.; Qiao, W.J.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Huang, J.Y. Substrate quality and soil environmental conditions predict litter decomposition and drive soil nutrient dynamics following afforestation on the Loess Plateau of China. Geoderma 2018, 325, 152–161. [Google Scholar] [CrossRef]
- Allison, S.D.; Lu, Y.; Weihe, C.; Goulden, M.L.; Martiny, A.C.; Treseder, K.K.; Martiny, J.B.H. Microbial abundance and composition influence litter decomposition response to environmental change. Ecology 2013, 94, 714–725. [Google Scholar] [CrossRef] [Green Version]
- Wilpert, K.V. Forest Soils—What’s Their Peculiarity? Soil Syst. 2022, 6, 5–10. [Google Scholar] [CrossRef]
- Spiecker, H. Norway Spruce Conversion: Options and Consequences; Brill: Leiden, The Netherland; Boston, MA, USA; Köln, Germany, 2004. [Google Scholar]
- Otto, H.J. Die verwirklichung des LÖWE_Regierungsprogramms. AFZ/Wald 1995, 50, 10269. [Google Scholar]
- Wulf, M. Forest policy in the EU and its influence on the plant diversity of woodlands. J. Environ. Manag. 2003, 67, 15–25. [Google Scholar] [CrossRef]
- Asplund, J.; Kauserud, H.; Bokhorst, S.; Lie, M.H.; Ohlson, M.; Nybakken, L. Fungal communities influence decomposition rates of plant litter from two dominant tree species. Fungal Ecol. 2018, 32, 1–8. [Google Scholar] [CrossRef]
- Berger, T.W.; Berger, P. Greater accumulation of litter in spruce (Picea abies) compared to beech (Fagus sylvatica) stands is not a consequence of the inherent recalcitrance of needles. Plant Soil 2012, 358, 349–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vesterdal, L. Influence of soil type on mass loss and nutrient release from decomposing foliage litter of beech and Norway spruce. Can. J. For. Res. 1999, 29, 95–105. [Google Scholar] [CrossRef]
- Liu, Q.; Zhuang, L.; Yin, R.; Ni, X.; You, C.; Yue, K.; Tan, B.; Liu, Y.; Zhang, L.; Xu, Z. Root diameter controls the accumulation of humic substances in decomposing root litter. Geoderma 2019, 348, 68–75. [Google Scholar] [CrossRef]
- Fanin, N.; Bertrand, I. Aboveground litter quality is a better predictor than belowground microbial communities when estimating carbon mineralization along a land-use gradient. Soil Biol. Biochem. 2016, 94, 48–60. [Google Scholar] [CrossRef]
- Gholz, H.L.; Wedin, D.A.; Smitherman, S.M.; Harmon, M.E.; Parton, W.J. Long-term dynamics of pine and hardwood litter in contrasting environments: Toward a global model of decomposition. Glob. Change Biol. 2000, 6, 751–765. [Google Scholar] [CrossRef]
- Ayres, E.; Steltzer, H.; Simmons, B.L.; Simpson, R.T.; Steinweg, J.M.; Wallenstein, M.D.; Mellor, N.; Parton, W.J.; Moore, J.C.; Wall, D.H. Home-field advantage accelerates leaf litter decomposition in forests. Soil Biol. Biochem. 2009, 41, 606–610. [Google Scholar] [CrossRef]
- Kang, H.; Gao, H.; Yu, W.; Yi, Y.; Wang, Y.; Ning, M. Changes in soil microbial community structure and function after afforestation depend on species and age: Case study in a subtropical alluvial island. Sci. Total Environ. 2018, 625, 1423–1432. [Google Scholar] [CrossRef]
- Prescott, C.E.; Blevins, L.L.; Staley, C.L. Effects of clear-cutting on decomposition rates of litter and forest floor in forests of British Columbia. Can. J. For. Res.-Rev. Can. Rech. For. 2000, 30, 1751–1757. [Google Scholar] [CrossRef]
- Ishikawa, H.; Osono, T.; Takeda, H. Effects of clear-cutting on decomposition processes in leaf litter and the nitrogen and lignin dynamics in a temperate secondary forest. J. For. Res. 2007, 12, 247–254. [Google Scholar] [CrossRef]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth—A meta-analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar] [CrossRef]
- Pennanen, T.; Liski, J.; Bååth, E.; Kitunen, V.; Uotila, J.; Westman, C.J.; Fritze, H. Structure of the microbial communities in coniferous forest soils in relation to site fertility and stand development stage. Microb. Ecol. 1999, 38, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Kohout, P.; Charvátová, M.; Štursová, M.; Mašínová, T.; Tomšovský, M.; Baldrian, P. Clearcutting alters decomposition processes and initiates complex restructuring of fungal communities in soil and tree roots. ISME J. 2018, 12, 692. [Google Scholar] [CrossRef] [PubMed]
- Finér, L.; Jurgensen, M.; Palviainen, M.; Piirainen, S.; Page-Dumroese, D. Does clear-cut harvesting accelerate initial wood decomposition? A five-year study with standard wood material. For. Ecol. Manag. 2016, 372, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Gliksman, D.; Haenel, S.; Osem, Y.; Yakir, D.; Zangy, E.; Preisler, Y.; Grünzweig, J.M. Litter decomposition in Mediterranean pine forests is enhanced by reduced canopy cover. Plant Soil 2018, 422, 317–329. [Google Scholar] [CrossRef]
- Keiser, A.D.; Keiser, D.A.; Strickland, M.S.; Bradford, M.A. Disentangling the mechanisms underlying functional differences among decomposer communities. J. Ecol. 2014, 102, 603–609. [Google Scholar] [CrossRef]
- Paterson, E.; Osler, G.; Dawson, L.A.; Gebbing, T.; Sim, A.; Ord, B. Labile and recalcitrant plant fractions are utilised by distinct microbial communities in soil: Independent of the presence of root and mycorrhizal fungi. Soil Biol. Biochem. 2008, 40, 1103–1113. [Google Scholar] [CrossRef]
- Chomel, M.; Guittonny-Larchevêque, M.; DesRochers, A.; Baldy, V. Home Field Advantage of Litter Decomposition in Pure and Mixed Plantations under Boreal Climate. Ecosystems 2015, 18, 1014–1028. [Google Scholar] [CrossRef] [Green Version]
- Veen, G.F.; Sundqvist, M.K.; Wardle, D.A. Environmental factors and traits that drive plant litter decomposition do not determine home-field advantage effects. Funct. Ecol. 2015, 29, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Veen, G.F.; Freschet, G.T.; Ordonez, A.; Wardle, D.A. Litter quality and environmental controls of home-field advantage effects on litter decomposition. Oikos 2015, 124, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Michener, R.; Lajtha, K. Stable Isotopes in Ecology and Environmental Science; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Gautam, M.K.; Lee, K.-S.; Song, B.-Y.; Lee, D.; Bong, Y.-S. Early-stage changes in natural 13C and 15N abundance and nutrient dynamics during different litter decomposition. J. Plant Res. 2016, 129, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Osono, T.; Takeda, H.; Azuma, J.-i. Carbon isotope dynamics during leaf litter decomposition with reference to lignin fractions. Ecol. Res. 2008, 23, 51–55. [Google Scholar] [CrossRef]
- Dijkstra, P.; LaViolette, C.M.; Coyle, J.S.; Doucett, R.R.; Schwartz, E.; Hart, S.C.; Hungate, B.A. 15N enrichment as an integrator of the effects of C and N on microbial metabolism and ecosystem function. Ecol. Lett. 2008, 11, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Pardo, L.H.; Hemond, H.F.; Montoya, J.P.; Fahey, T.J.; Siccama, T.G. Response of the natural abundance of 15N in forest soils and foliage to high nitrate loss following clear-cutting. Can. J. For. Res. 2002, 32, 1126–1136. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Aidar, M.P.M.; Bustamante, M.; Dawson, T.E.; Hobbie, E.A.; Kahmen, A.; Mack, M.C.; McLauchlan, K.K.; Michelsen, A.; et al. Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nitrient concentrations, and nitrogen availability. New Phytol. 2009, 183, 980–992. [Google Scholar] [CrossRef]
- Jiang, C.; Séquaris, J.-M.; Vereecken, H.; Klumpp, E. Effects of temperature and associated organic carbon on the fractionation of water-dispersible colloids from three silt loam topsoils under different land use. Geoderma 2017, 299, 43–53. [Google Scholar] [CrossRef]
- Siebers, N.; Kruse, J. Short-term impacts of forest clear-cut on soil structure and consequences for organic matter composition and nutrient speciation: A case study. PLoS ONE 2019, 14, e0220476. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, U.; Bogena, H.R.; Herbst, M.; Huisman, J.A.; Peterson, T.J.; Weuthen, A.; Western, A.W.; Vereecken, H. Seasonal and event dynamics of spatial soil moisture patterns at the small catchment scale. Water Resour. Res. 2012, 48, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Wiekenkamp, I.; Huisman, J.A.; Bogena, H.R.; Graf, A.; Lin, H.S.; Druee, C.; Vereecken, H. Changes in measured spatiotemporal patterns of hydrogical response after partial deforestation in a headwater catchment. J. Hydrol. 2016, 542, 648–661. [Google Scholar] [CrossRef]
- Havlik, D. Das Klima von Aachen. Aachen. Geogr. Arb. 2002, 36, 1–20. [Google Scholar]
- Etmann, M. Dendrologische Aufnahmen im Wassereinzugsgebiet oberer Wüstebach anhand verschiedener Mess-und Schätzverfahren. Master’s Thesis, Westfälische Wilhelms-University Münster, Münster, Germany, 2009. [Google Scholar]
- Bradford, M.A.; Tordoff, G.M.; Eggers, T.; Jones, T.H.; Newington, J.E. Microbiota, fauna, and mesh size interactions in litter decomposition. Oikos 2002, 99, 317–323. [Google Scholar] [CrossRef]
- Wang, Q.K.; Zhong, M.C.; He, T.X. Home-field advantage of litter decomposition and nitrogen release in forest ecosystems. Biol. Fertil. Soils 2013, 49, 427–434. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Aneja, M.K.; Sharma, S.; Fleischmann, F.; Stich, S.; Heller, W.; Bahnweg, G.; Munch, J.C.; Schloter, M. Microbial colonization of beech and spruce litter—Influence of decomopsition site and plant litter species on the diversity of microbial community. Micobial Ecol. 2006, 52, 127–135. [Google Scholar] [CrossRef]
- Zhou, X.; Zhu, H.; Wen, Y.; Goodale, U.M.; Li, X.; You, Y.; Ye, D.; Liang, H. Effects of understory management on trade-offs and synergies between biomass carbon stock, plant diversity and timber production in eucalyptus plantations. For. Ecol. Manag. 2018, 410, 164–173. [Google Scholar] [CrossRef]
- Heim, A.; Frey, B. Early stage litter decomposition rates for Swiss forests. Biogeochemistry 2004, 70, 299–313. [Google Scholar] [CrossRef]
- McDowell, N.G.; Allen, C.D.; Marshall, L. Growth, carbon-isotope discrimination, and drought-associated mortality across a Pinus ponderosa elevational transect. Glob. Change Biol. 2010, 16, 399–415. [Google Scholar] [CrossRef]
- Rajala, T.; Peltoniemi, M.; Hantula, J.; Mäkipää, R.; Pennanen, T. RNA reveals a succession of active fungi during the decay of Norway spruce logs. Fungal Ecol. 2011, 4, 437–448. [Google Scholar] [CrossRef]
- St John, M.G.; Orwin, K.H.; Dickie, I.A. No ‘home’ versus ‘away’ effects of decomposition found in a grassland-forest reciprocal litter transplant study. Soil Biol. Biochem. 2011, 43, 1482–1489. [Google Scholar] [CrossRef]
- Freschet, G.T.; Aerts, R.; Cornelissen, J.H.C. Multiple mechanisms for trait effects on litter decomposition: Moving beyond home-field advantage with a new hypothesis. J. Ecol. 2012, 100, 619–630. [Google Scholar] [CrossRef]
- Petraglia, A.; Cacciatori, C.; Chelli, S.; Fenu, G.; Calderisi, G.; Gargano, D.; Abeli, T.; Orsenigo, S.; Carbognani, M. Litter decomposition: Effects of temperature driven by soil moisture and vegetation type. Plant Soil 2019, 435, 187–200. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776. [Google Scholar] [CrossRef]
- Mansfield, S.D.; Bärlocher, F. Determination of Soluble Carbohydrates. In Methods to Study Litter Decomposition: A Practical Guide; Graça, M.A.S., Bärlocher, F., Mark, O., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 85–90. [Google Scholar]
- Ahmed, I.U.; Mengistie, H.K.; Godbold, D.L.; Sandén, H. Soil moisture integrates the influence of land-use and season on soil microbial community composition in the Ethiopian highlands. Appl. Soil Ecol. 2019, 135, 85–90. [Google Scholar] [CrossRef]
- Mueller, P.; Schile-Beers, L.M.; Mozdzer, T.J.; Chmura, G.L.; Dinter, T.; Kuzyakov, Y.; de Groot, A.V.; Esselink, P.; Smit, C.; D’Alpaos, A.; et al. Global-change effects on early-stage decomposition processes in tidal wetlands—Implications from a global survey using standardized litter. Biogeosciences 2018, 15, 3189–3202. [Google Scholar] [CrossRef] [Green Version]
- Preston, C.M.; Nault, J.R.; Trofymow, J.A. Chemical Changes During 6 Years of Decomposition of 11 Litters in Some Canadian Forest Sites. Part 2. 13C Abundance, Solid-State 13C NMR Spectroscopy and the Meaning of “Lignin”. Ecosystems 2009, 12, 1078–1102. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Högberg, P. Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen dynamics. New Phytol. 2012, 196, 367–382. [Google Scholar] [CrossRef]
- Ngao, J.; Cotrufo, M.F. Carbon isotope discrimination during litter decomposition can be explained by selective use of substrate with differing δ13C. Biogeosci. Discuss 2011, 2011, 51–82. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Litter Quality | Residual Quality after 1 Year of Decomposition | |||||
|---|---|---|---|---|---|---|
| Beech | Spruce | Forest | Clear-Cut | |||
| Beech | Spruce | Beech | Spruce | |||
| C (%) | 47.1 ± 0.2a | 48.4 ± 0.1b | 48.2 ± 0.8a | 49.5 ± 0.6b | 46.1 ± 0.6a | 47.9 ± 0.5b |
| N (%) | 2.1 ± 0.1a | 1.2 ± 0.0b | 3.0 ± 0.1a | 1.9 ± 0.8b | 2.8 ± 0.1a | 1.6 ± 0.1b |
| P (mg kg−1) | 278.4 ± 10.4a | 254.9 ± 8.6b | 105.2 ± 6.3a | 81.0 ± 8.9b | 104.5 ± 6.0a | 61.5 ± 4.9b |
| C:N | 22.4 ± 0.5a | 39.0 ± 1.3b | 20.2 ± 0.6a | 34.4 ± 0.7b | 20.8 ± 0.8a | 35.1 ± 0.4b |
| Ca (mg kg−1) | 2.18 ± 0.04a | 1.67 ± 0.01b | 1.02 ± 0.09a | 0.74 ± 0.08b | 1.20 ± 0.05a | 0.79 ± 0.06b |
| Cambisol | Gleysol | |
|---|---|---|
| Mass loss | 11.2 ± 0.5a | 3.7 ± 1.0b |
| C release | 14.0 ± 2.5a | 10.7 ± 0.9b |
| N release | 28.3 ± 0.9a | 43.1 ± 5.4b |
| Effects | Remaining | ||||
|---|---|---|---|---|---|
| df | C | N | P | Ca | |
| Soil type | 1 | 6.8 * | 0.7 | 8.4 * | 0.1 |
| Stand × Soil type | 1 | 2.9 | 2.1 | 4.3 | 15.6 ** |
| Soil type × Species | 1 | 31.6 *** | 0.6 | 0.0 | 0.0 |
| Stand × Soil type × Species | 1 | 6.3 * | 1.3 | 8.5* | 3.2 |
| Variables | Coefficients | R2 | |
|---|---|---|---|
| Spruce | |||
| Clear-cut | N, P | 0.65, −0.36 | 0.93 |
| Forest | N, Ca | 0.77, −0.23 | 0.97 |
| Beech | |||
| Clear-cut | Ca, C:N | −0.51, −0.49 | 0.85 |
| Forest | C:N | −0.82 | 0.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuang, L.; Schnepf, A.; Unger, K.; Liang, Z.; Bol, R. Home-Field Advantage of Litter Decomposition Faded 8 Years after Spruce Forest Clearcutting in Western Germany. Soil Syst. 2022, 6, 26. https://doi.org/10.3390/soilsystems6010026
Zhuang L, Schnepf A, Unger K, Liang Z, Bol R. Home-Field Advantage of Litter Decomposition Faded 8 Years after Spruce Forest Clearcutting in Western Germany. Soil Systems. 2022; 6(1):26. https://doi.org/10.3390/soilsystems6010026
Chicago/Turabian StyleZhuang, Liyan, Andrea Schnepf, Kirsten Unger, Ziyi Liang, and Roland Bol. 2022. "Home-Field Advantage of Litter Decomposition Faded 8 Years after Spruce Forest Clearcutting in Western Germany" Soil Systems 6, no. 1: 26. https://doi.org/10.3390/soilsystems6010026
APA StyleZhuang, L., Schnepf, A., Unger, K., Liang, Z., & Bol, R. (2022). Home-Field Advantage of Litter Decomposition Faded 8 Years after Spruce Forest Clearcutting in Western Germany. Soil Systems, 6(1), 26. https://doi.org/10.3390/soilsystems6010026