

Salt Stress Highlights the Relevance of Genotype × Genotype Interaction in the Nitrogen-Fixing Symbiosis between Sinorhizobium meliloti and Alfalfa

 , , , , ,
, , , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Alfalfa-Root-Nodule Rhizobia
2.2. 16S rRNA Gene Sequence Amplification and RAPD Analysis
2.3. Growth Kinetics of Rhizobial Strains under Saline Conditions
2.4. Germination of M. sativa Cultivars on NaCl Gradient
2.5. Growth of M. sativa Cultivars on NaCl Gradient
2.6. S. meliloti—M. sativa Nodulation assay in Saline Conditions
2.7. Determination of Strains Survival on Carriers
2.8. Small-Scale in-Field Experiment
2.9. Statistical Analysis
3. Results
3.1. Isolation and Identification of Rhizobial Strains
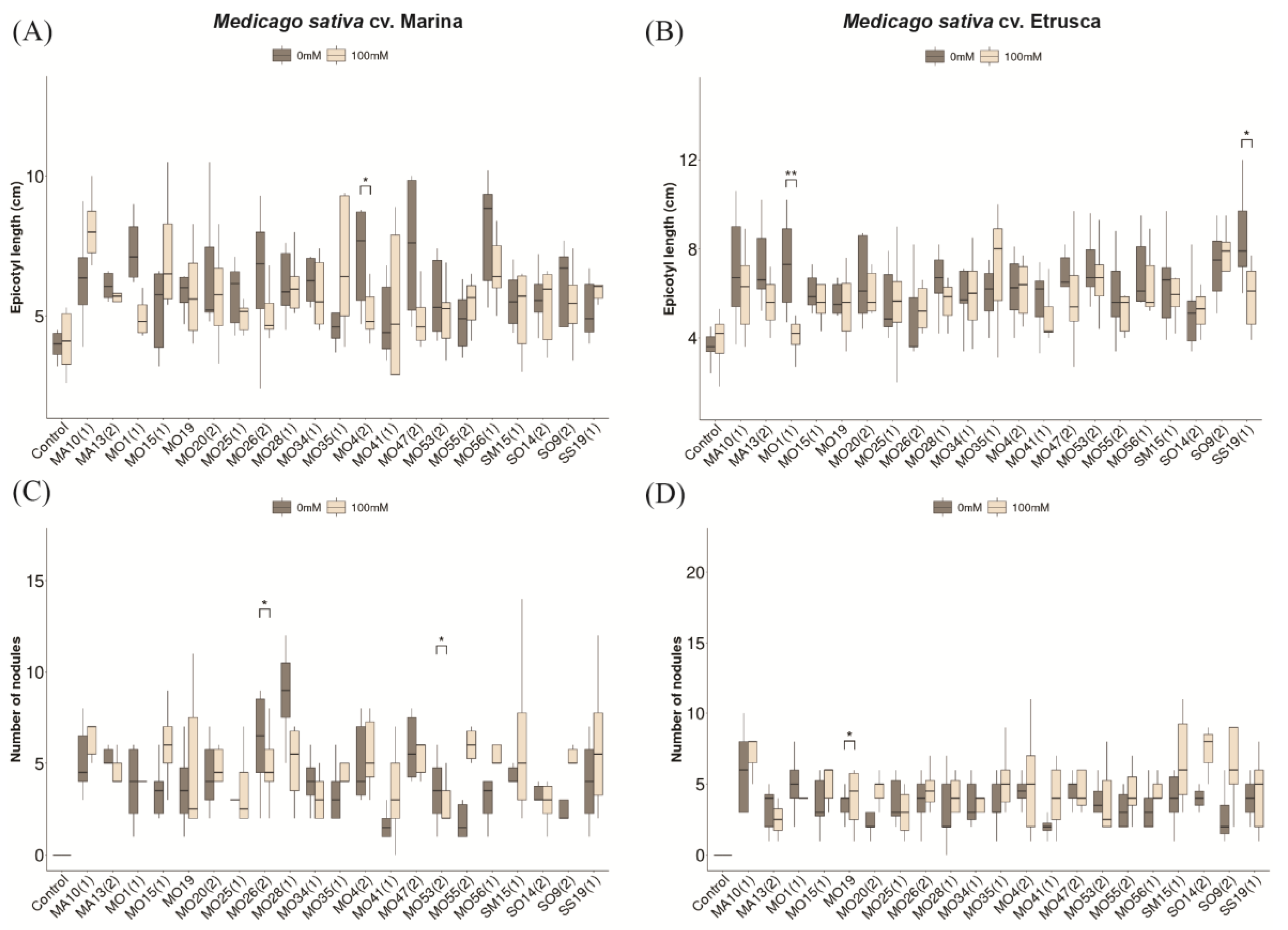
3.2. Salt Tolerance of Medicago sativa Cultivars Etrusca and Marina under Increasing Levels of NaCl Concentration
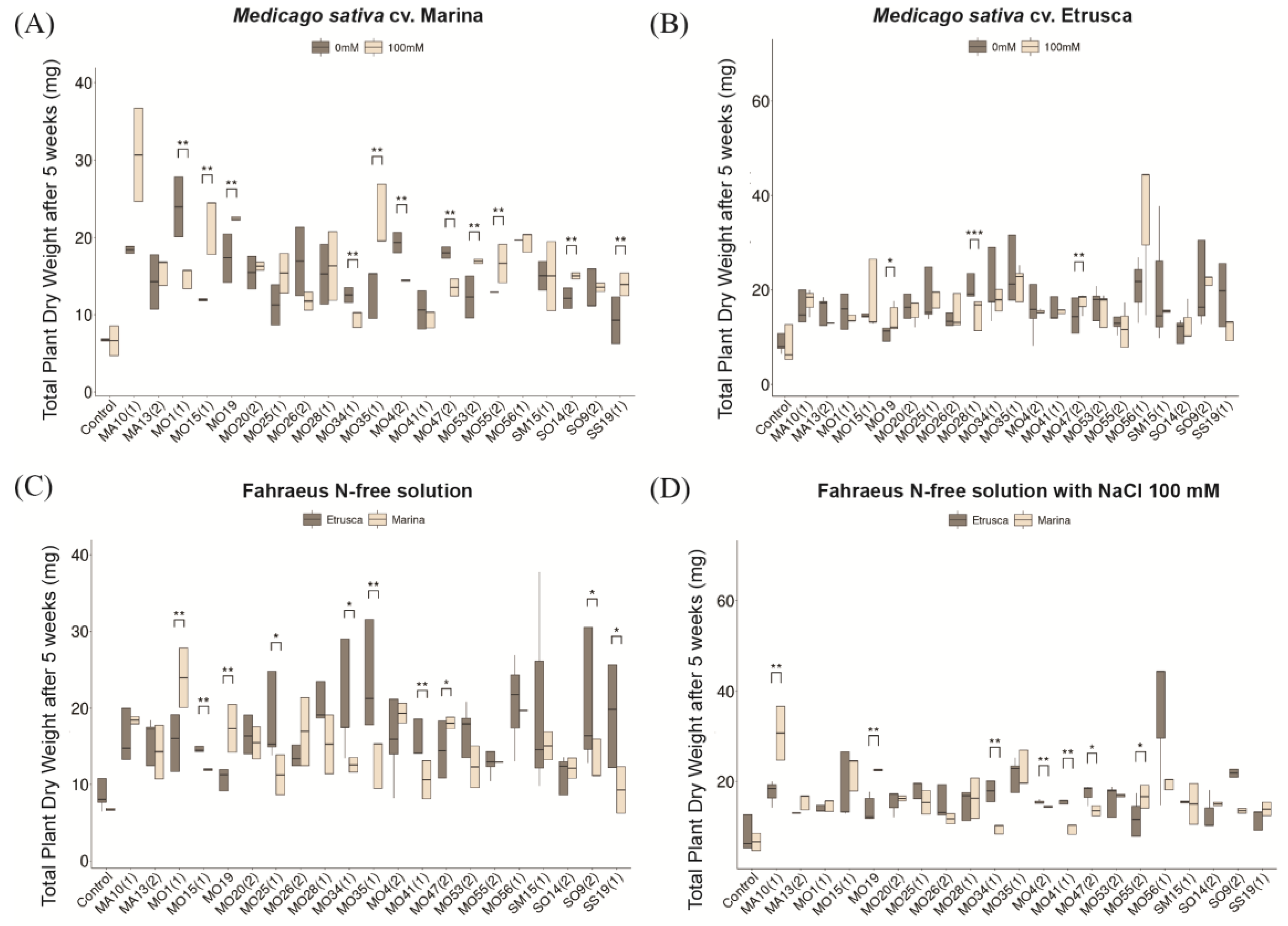
3.3. Effect of Sodium Chloride and Symbiosis on M. sativa (Cultivars Etrusca and Marina) Growth
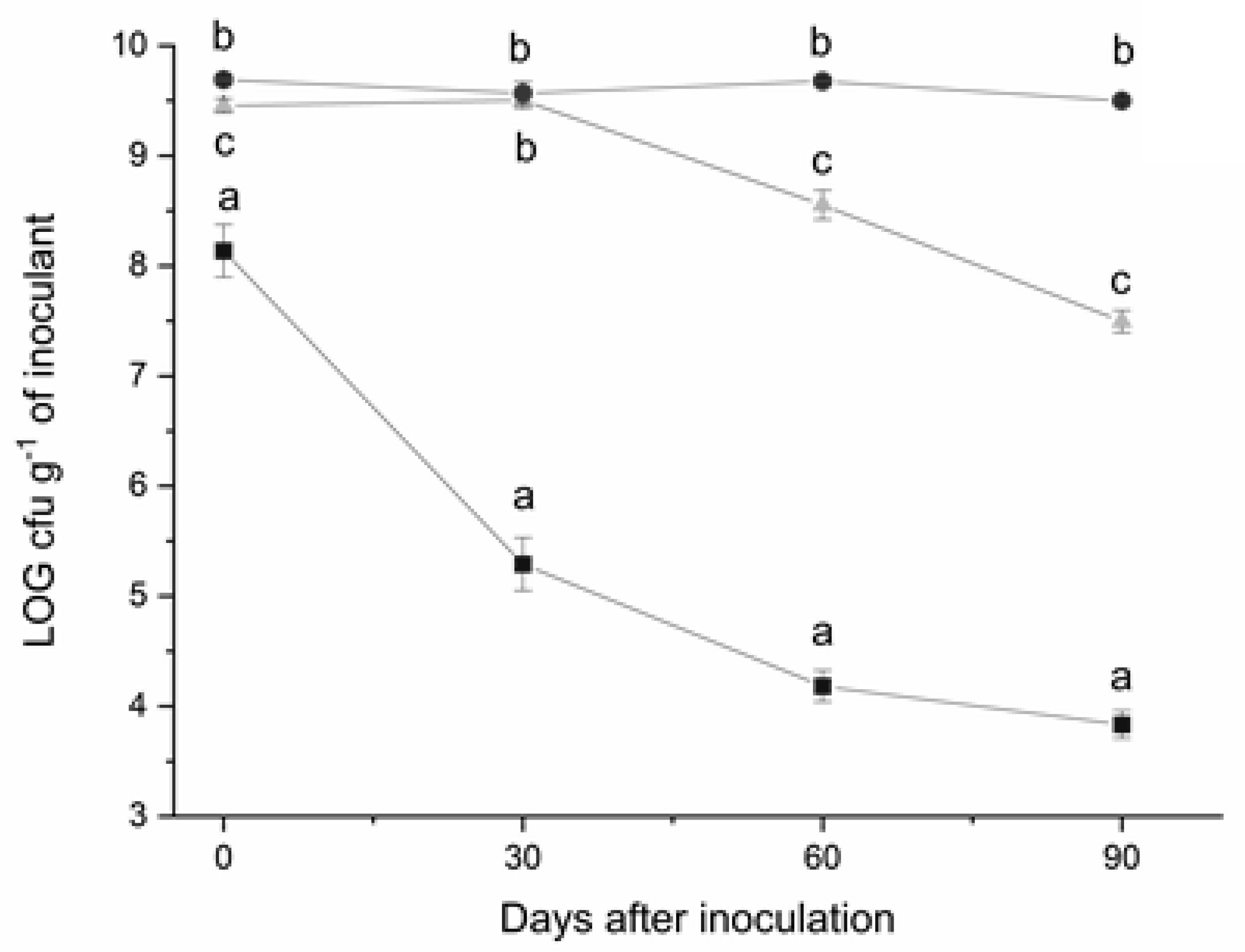
3.4. Evaluation of Carriers for the Formulation of S. meliloti Inoculants and Preliminary in-Field Trial
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Acharya, J.P.; Lopez, Y.; Gouveia, B.T.; de Bem Oliveira, I.; Resende, M.F.R.; Muñoz, P.R.; Rios, E.F. Breeding Alfalfa (Medicago sativa L.) Adapted to Subtropical Agroecosystems. Agronomy 2020, 10, 742. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Li, Y.P.; Sun, J.; Huang, G.H. Optimizing water resources allocation and soil salinity control for supporting agricultural and environmental sustainable development in Central Asia. Sci. Total Environ. 2020, 704, 135281. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Navarro, V.; Zornoza, R.; Faz, Á.; Fernández, J.A. Comparison of soil organic carbon pools, microbial activity and crop yield and quality in two vegetable multiple cropping systems under mediterranean conditions. Sci. Hortic. 2020, 261, 109025. [Google Scholar] [CrossRef]
- Arzani, A.; Ashraf, M. Smart Engineering of Genetic Resources for Enhanced Salinity Tolerance in Crop Plants. CRC Crit. Rev. Plant Sci. 2016, 35, 146–189. [Google Scholar] [CrossRef]
- Muntyan, V.S.; Roumiantseva, M.L. Molecular Phylogenetic Analysis of Salt-Tolerance-Related Genes in Root-Nodule Bacteria Species Sinorhizobium meliloti. Agronomy 2022, 12, 1968. [Google Scholar] [CrossRef]
- FAO. World Food and Agriculture—Statistical Yearbook 2021; FAO: Rome, Italy, 2021. [Google Scholar]
- Viti, C.; Bellabarba, A.; Daghio, M.; Mengoni, A.; Mele, M.; Buccioni, A.; Pacini, G.C.; Bekki, A.; Azim, K.; Hafidi, M.; et al. Alfalfa for a Sustainable Ovine Farming System: Proposed Research for a New Feeding Strategy Based on Alfalfa and Ecological Leftovers in Drought Conditions. Sustainability 2021, 13, 3880. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.K.; Gowda, C.L.L.; Krishnamurthy, L. Plant growth promoting rhizobia: Challenges and opportunities. 3 Biotech 2015, 5, 355–377. [Google Scholar] [CrossRef]
- Kaiwen, G.; Zisong, X.; Yuze, H.; Qi, S.; Yue, W.; Yanhui, C.; Jiechen, W.; Wei, L.; Huihui, Z. Effects of salt concentration, pH, and their interaction on plant growth, nutrient uptake, and photochemistry of alfalfa (Medicago sativa) leaves. Plant Signal. Behav. 2020, 15, 1832373. [Google Scholar] [CrossRef]
- Jensen, E.S.; Hauggaard-Nielsen, H. How can increased use of biological N2 fixation in agriculture benefit the environment? Plant Soil 2003, 252, 177–186. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Wang, T. Enhanced salt tolerance of alfalfa (Medicago sativa) by rstB gene transformation. Plant Sci. 2015, 234, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, S.; Biswas, D.; Fu, Y.-B.; Biligetu, B. Morphological, Physiological, and Genetic Responses to Salt Stress in Alfalfa: A Review. Agronomy 2020, 10, 577. [Google Scholar] [CrossRef]
- Jones, K.M.; Kobayashi, H.; Davies, B.W.; Taga, M.E.; Walker, G.C. How rhizobial symbionts invade plants: The Sinorhizobium-Medicago model. Nat. Rev. Microbiol. 2007, 5, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Cangioli, L.; Vaccaro, F.; Fini, M.; Mengoni, A.; Fagorzi, C. Scent of a Symbiont: The Personalized Genetic Relationships of Rhizobium-Plant Interaction. Int. J. Mol. Sci. 2022, 23, 3358. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, L.T. Evolving together, evolving apart: Measuring the fitness of rhizobial bacteria in and out of symbiosis with leguminous plants. New Phytol. 2020, 228, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Fagorzi, C.; Bacci, G.; Huang, R.; Cangioli, L.; Checcucci, A.; Fini, M.; Perrin, E.; Natali, C.; DiCenzo, G.C.; Mengoni, A. Nonadditive transcriptomic signatures of genotype-by-genotype interactions during the initiation of plant-rhizobium symbiosis. mSystems 2021, 6, e00974-20. [Google Scholar] [CrossRef]
- Kang, W.; Jiang, Z.; Chen, Y.; Wu, F.; Liu, C.; Wang, H.; Shi, S.; Zhang, X.-X. Plant transcriptome analysis reveals specific molecular interactions between alfalfa and its rhizobial symbionts below the species level. BMC Plant Biol. 2020, 20, 293. [Google Scholar] [CrossRef]
- Beringer, J.E. R factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 1974, 84, 188–198. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Wiley: New York, NY, USA, 1991. [Google Scholar]
- Kumar, P.P.; Antony, L.V.M.; Kumar, M. Phylogenetic diversity of cultivable bacteria associated with filamentous non-hetrocystous marine cyanobacteria. J. Algal Biomass Util. 2009, 1, 87–102. [Google Scholar]
- Caccamo, D.; Di Cello, F.; Fani, R.; Gugliandolo, C.; Maugeri, T.L. Polyphasic approach to the characterisation of marine luminous bacteria. Res. Microbiol. 1999, 150, 221–230. [Google Scholar] [CrossRef]
- Sneath, P.H.A.; Sokal, R.R. Numerical taxonomy. Nature 1962, 93, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software packages for education and data analysis. Palaeontol. Electron. 2001, 4, XIX–XX. [Google Scholar]
- Rumbaugh, M.D. Salt tolerance of germinating alfalfa seeds. Standard Tests to Characterize Alfalfa Cultivars. Edition, 1991, 3. Available online: http://www.naaic.org/stdtests/saltseeds.pdf (accessed on 27 September 2023).
- Scasta, J.D.; Trostle, C.L.; Foster, M.A. Evaluating Alfalfa (Medicago sativa L.) Cultivars for Salt Tolerance Using Laboratory, Greenhouse and Field Methods. J. Agric. Sci. 2012, 4, p90. [Google Scholar] [CrossRef]
- Weast, R.C. CRC Handbook of Chemistry and Physics: 1974–1975, 55th ed.; CRC Handbook of Chemistry and Physics: A Ready-reference Book of Chemical and Physical Data; CRC Press: Boca Raton, FL, USA, 1974; ISBN 9780878194544. [Google Scholar]
- Durand, J.H. Les Sols Irrigables; Etude Pedologique; Agence de Coopération Culturelle et Technique, Presses Universitaires de France: Paris, France, 1983; 339p. [Google Scholar]
- Fahraeus, G. The infection of clover root hairs by nodule bacteria studied by a simple glass slide technique. J. Gen. Microbiol. 1957, 16, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Bueis, R.; Sánchez-Cañizares, C.; James, E.K.; González-Andrés, F. Formulation of a Highly Effective Inoculant for Common Bean Based on an Autochthonous Elite Strain of Rhizobium leguminosarum bv. phaseoli, and Genomic-Based Insights into Its Agronomic Performance. Front. Microbiol. 2019, 10, 2724. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, P.; Moschini, V.; Tittarelli, F.; Ciaccia, C.; Benedettelli, S.; Vazzana, C.; Canali, S. Agronomic performance, carbon storage and nitrogen utilisation of long-term organic and conventional stockless arable systems in Mediterranean area. Eur. J. Agron. 2014, 52, 138–145. [Google Scholar] [CrossRef]
- Howieson, J.G.; Dilworth, M.J. (Eds.) Working with Rhizobia; Australian Centre for International Agricultural Research: Canberra, Australian, 2016.
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Community Ecology Package. R Package Version 2018, 2, 5-2. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef]
- Wan, W.; Liu, Q.; Zhang, C.; Li, K.; Sun, Z.; Li, Y.; Li, H. Alfalfa growth and nitrogen fixation constraints in salt-affected soils are in part offset by increased nitrogen supply. Front. Plant Sci. 2023, 14, 358. [Google Scholar] [CrossRef] [PubMed]
- Villadas, P.J.; Burgos, P.; Jording, D.; Selbitschka, W.; Pühler, A.; Toro, N. Comparative analysis of the genetic structure of a Rhizobium meliloti field population before and after environmental release of the highly competitive R. meliloti strain GR4. FEMS Microbiol. Ecol. 1996, 21, 37–45. [Google Scholar] [CrossRef]
- Bellabarba, A.; Bacci, G.; Decorosi, F.; Aun, E.; Azzarello, E.; Remm, M.; Giovannetti, L.; Viti, C.; Mengoni, A.; Pini, F.; et al. Competitiveness for Nodule Colonization in Sinorhizobium meliloti: Combined In Vitro-Tagged Strain Competition and Genome-Wide Association Analysis. mSystems 2021, 6, e00550-21. [Google Scholar] [CrossRef] [PubMed]
- Ke, J.; Wang, B.; Yoshikuni, Y. Microbiome Engineering: Synthetic Biology of Plant-Associated Microbiomes in Sustainable Agriculture. Trends Biotechnol. 2021, 39, 244–261. [Google Scholar] [CrossRef] [PubMed]
- Sessitsch, A.; Pfaffenbichler, N.; Mitter, B. Microbiome Applications from Lab to Field: Facing Complexity. Trends Plant Sci. 2019, 24, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, F.; Cangioli, L.; Mengoni, A.; Fagorzi, C. Synthetic plant microbiota challenges in nonmodel species. Trends Microbiol. 2022, 30, 922–924. [Google Scholar] [CrossRef] [PubMed]
- Evenson, R.E.; Gollin, D. Assessing the Impact of the Green Revolution, 1960 to 2000. Science 2003, 300, 758–762. [Google Scholar] [CrossRef]
- Swaraj, K.; Bishnoi, N.R. Effect of salt stress on nodulation and nitrogen fixation in legumes. Indian J. Exp. Biol. 1999, 37, 843–848. [Google Scholar]
- Miransari, M.; Smith, D.L. Overcoming the Stressful Effects of Salinity and Acidity on Soybean Nodulation and Yields Using Signal Molecule Genistein Under Field Conditions. J. Plant Nutr. 2007, 30, 1967–1992. [Google Scholar] [CrossRef]
- Batstone, R.T.; Lindgren, H.; Cassandra, M.A.; Goralka, L.A.; Riley, A.B.; Grillo, M.A.; Marshall-Colon, A.; Heath, K.D. Genome-Wide Association Studies across Environmental and Genetic Contexts Reveal Complex Genetic Architecture of Symbiotic Extended Phenotypes. MBio 2022, 13, e01823-22. [Google Scholar] [CrossRef]
- Chakraborty, S.; Driscoll, H.E.; Abrahante, J.E.; Zhang, F.; Fisher, R.F.; Harris, J.M. Salt Stress Enhances Early Symbiotic Gene Expression in Medicago truncatula and Induces a Stress-Specific Set of Rhizobium-Responsive Genes. Mol. Plant. Microbe. Interact. 2021, 34, 904–921. [Google Scholar] [CrossRef] [PubMed]
- Nishad, R.; Ahmed, T.; Rahman, V.J.; Kareem, A. Modulation of Plant Defense System in Response to Microbial Interactions. Front. Microbiol. 2020, 11, 1298. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, P.; Zhou, Y.; Hu, T.; Zhang, P.; Wu, Y. A proteomic approach to understand the impact of nodulation on salinity stress response in alfalfa (Medicago sativa L.). Plant Biol. 2022, 24, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, F.; Mengoni, A. Legume–Rhizobium Symbiosis and Beyond: Producing Synthetic Communities for Increasing Crop Production Under Climate Change Challenges BT—Bioinoculants: Biological Option for Mitigating global Climate Change; Singh, S., Prasanna, R., Pranaw, K., Eds.; Springer Nature: Singapore, 2023; pp. 147–162. ISBN 978-981-99-2973-3. [Google Scholar]
- Mitter, B.; Brader, G.; Pfaffenbichler, N.; Sessitsch, A. Next generation microbiome applications for crop production—limitations and the need of knowledge-based solutions. Curr. Opin. Microbiol. 2019, 49, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Jin, K.; Raaijmakers, J.M. Designing a home for beneficial plant microbiomes. Curr. Opin. Plant Biol. 2021, 62, 102025. [Google Scholar] [CrossRef] [PubMed]
- Hungria, M.; Loureiro, M.F.; Mendes, I.C.; Campo, R.J.; Graham, P.H. Inoculant Preparation, Production and Application BT—Nitrogen Fixation in Agriculture, Forestry, Ecology, and the Environment; Werner, D., Newton, W.E., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 223–253. ISBN 978-1-4020-3544-9. [Google Scholar]
- Albareda, M.; Rodríguez-Navarro, D.N.; Camacho, M.; Temprano, F.J. Alternatives to peat as a carrier for rhizobia inoculants: Solid and liquid formulations. Soil Biol. Biochem. 2008, 40, 2771–2779. [Google Scholar] [CrossRef]
- Ben Rebah, F.; Prévost, D.; Yezza, A.; Tyagi, R.D. Agro-industrial waste materials and wastewater sludge for rhizobial inoculant production: A review. Bioresour. Technol. 2007, 98, 3535–3546. [Google Scholar] [CrossRef]
- Beck, D.P. Suitability of charcoal-amended mineral soil as carrier for Rhizobium inoculants. Soil Biol. Biochem. 1991, 23, 41–44. [Google Scholar] [CrossRef]
- Daza, A.; Santamaría, C.; Rodríguez-Navarro, D.N.; Camacho, M.; Orive, R.; Temprano, F. Perlite as a carrier for bacterial inoculants. Soil Biol. Biochem. 2000, 32, 567–572. [Google Scholar] [CrossRef]
- Khavazi, K.; Rejali, F.; Seguin, P.; Miransari, M. Effects of carrier, sterilisation method, and incubation on survival of Bradyrhizobium japonicum in soybean (Glycine max L.) inoculants. Enzyme Microb. Technol. 2007, 41, 780–784. [Google Scholar] [CrossRef]
- Hardy, K.; Knight, J.D. Evaluation of biochars as carriers for Rhizobium leguminosarum. Can. J. Microbiol. 2020, 67, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Lupwayi, N.Z.; Olsen, P.E.; Sande, E.S.; Keyser, H.H.; Collins, M.M.; Singleton, P.W.; Rice, W.A. Inoculant quality and its evaluation. F. Crop. Res. 2000, 65, 259–270. [Google Scholar] [CrossRef]
- Wendlandt, C.E.; Regus, J.U.; Gano-Cohen, K.A.; Hollowell, A.C.; Quides, K.W.; Lyu, J.Y.; Adinata, E.S.; Sachs, J.L. Host investment into symbiosis varies among genotypes of the legume Acmispon strigosus, but host sanctions are uniform. New Phytol. 2019, 221, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Wendlandt, C.E.; Gano-Cohen, K.A.; Stokes, P.J.N.; Jonnala, B.N.R.; Zomorrodian, A.J.; Al-Moussawi, K.; Sachs, J.L. Wild legumes maintain beneficial soil rhizobia populations despite decades of nitrogen deposition. Oecologia 2022, 198, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Regus, J.U.; Gano, K.A.; Hollowell, A.C.; Sachs, J.L. Efficiency of partner choice and sanctions in Lotus is not altered by nitrogen fertilization. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132587. [Google Scholar] [CrossRef]
- Grillo, M.A.; Stinchcombe, J.R.; Heath, K.D. Nitrogen addition does not influence pre-infection partner choice in the legume-rhizobium symbiosis. Am. J. Bot. 2016, 103, 1763–1770. [Google Scholar] [CrossRef]
- Mendoza-Suárez, M.A.; Geddes, B.A.; Sánchez-Cañizares, C.; Ramírez-González, R.H.; Kirchhelle, C.; Jorrin, B.; Poole, P.S. Optimizing Rhizobium-legume symbioses by simultaneous measurement of rhizobial competitiveness and N2 fixation in nodules. Proc. Natl. Acad. Sci. USA 2020, 117, 9822–9831. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Raizada, M.N. A meta-analysis of the effectiveness of diverse rhizobia inoculants on soybean traits under field conditions. Soil Biol. Biochem. 2017, 105, 177–196. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Region Name | Location | Electrical Conductivity (dS/m) | Geographic Coordinates |
|---|---|---|---|
| Fleuris | Municipality of Ben Freha (Wilaya of Oran) | 2 | 35°41′39.968″ N, 0°24′47.605″ E |
| Es-Senia | Wilaya of Oran | 1.53 | 35°38′11.675″ N, 0°37′56.653″ E |
| El Malah | Wilaya of Ain Temouchent | 2.5 | 35°23′23.513″ N, 1°5′51.691″ E |
| Oued Sebbah | Wilaya of Ain Temouchent | 3 | 35°22′15.229″ N, 0°48′24.655″ E |
| Marina | Etrusca | |||
|---|---|---|---|---|
| 0 mM NaCl | 100 mM NaCl | 0 mM NaCl | 100 mM NaCl | |
| Dry weight | <2 × 10−16 | <2 × 10−16 | <2 × 10−16 | <2 × 10−16 |
| Epicotyl length | 0.00025 | 7.1 × 10−5 | 9.7 × 10−9 | 1.5 × 10−7 |
| Number of nodules | 4.9 × 10−8 | 0.00059 | 5.3 × 10−10 | 9.0 × 10−8 |
| 0 mM NaCl Dataset | 100 mM NaCl Dataset | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PERMANOVA | PERMDISP | PERMANOVA | PERMDISP | |||||||
| R2 | F | p | F | p | R2 | F | p | F | p | |
| Strains | 0.17920 | 4.0737 | 1 × 10−4 *** | 1.4752 | 0.084 | 0.25123 | 5.3664 | 1 × 10−4 *** | 1.1292 | 0.301 |
| Cultivar | 0.04389 | 19.9531 | 1 × 10−4 *** | 0.1532 | 0.671 | 0.04802 | 20.5154 | 1 × 10−4 *** | 0.0321 | 0.874 |
| Strains:Cultivar | 0.21824 | 4.9611 | 1 × 10−4 *** | 2.0221 | 0.001 *** | 0.22792 | 4.8685 | 1 × 10−4 *** | 1.323 | 0.13 |
| Residual | 0.55867 | 0.47283 | ||||||||
| Total | 1.00000 | 1.00000 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellabarba, A.; Decorosi, F.; Fagorzi, C.; El Hadj Mimoune, A.; Buccioni, A.; Santoni, M.; Pacini, G.C.; Bekki, A.; Azim, K.; Hafidi, M.; et al. Salt Stress Highlights the Relevance of Genotype × Genotype Interaction in the Nitrogen-Fixing Symbiosis between Sinorhizobium meliloti and Alfalfa. Soil Syst. 2023, 7, 112. https://doi.org/10.3390/soilsystems7040112
Bellabarba A, Decorosi F, Fagorzi C, El Hadj Mimoune A, Buccioni A, Santoni M, Pacini GC, Bekki A, Azim K, Hafidi M, et al. Salt Stress Highlights the Relevance of Genotype × Genotype Interaction in the Nitrogen-Fixing Symbiosis between Sinorhizobium meliloti and Alfalfa. Soil Systems. 2023; 7(4):112. https://doi.org/10.3390/soilsystems7040112
Chicago/Turabian StyleBellabarba, Agnese, Francesca Decorosi, Camilla Fagorzi, Amina El Hadj Mimoune, Arianna Buccioni, Margherita Santoni, Gaio Cesare Pacini, Abdelkader Bekki, Khalid Azim, Majida Hafidi, and et al. 2023. "Salt Stress Highlights the Relevance of Genotype × Genotype Interaction in the Nitrogen-Fixing Symbiosis between Sinorhizobium meliloti and Alfalfa" Soil Systems 7, no. 4: 112. https://doi.org/10.3390/soilsystems7040112
APA StyleBellabarba, A., Decorosi, F., Fagorzi, C., El Hadj Mimoune, A., Buccioni, A., Santoni, M., Pacini, G. C., Bekki, A., Azim, K., Hafidi, M., Mazzoncini, M., Mengoni, A., Pini, F., & Viti, C. (2023). Salt Stress Highlights the Relevance of Genotype × Genotype Interaction in the Nitrogen-Fixing Symbiosis between Sinorhizobium meliloti and Alfalfa. Soil Systems, 7(4), 112. https://doi.org/10.3390/soilsystems7040112