In the present study, microbes were isolated from the soil collected from TBL (a landfill that was originally contaminated with leachate). The isolated microbes were then used for bioaugmentation as a method for remediating soil microcosm systems (previously non-contaminated urban soil) that were contaminated with leachate for bioremediation trials. The leachate taken from JSL was used to induce metal contamination in the microcosms. The results of toxic metal removal from leachate-contaminated soils are presented in this section.
3.2.1. Microbial Isolation from Originally Contaminated Soil
The original leachate-contaminated soil was characterized for bacteria based on separation and identification. Nine bacteria that were isolated from the leachate-contaminated soil are listed in
Table 5. Both Gram-positive and Gram-negative bacteria were isolated. Even though some of the identified microbes may not be depicted in the literature as bioremediation enhancers, their presence in leachate-contaminated soil sparks curiosity about their existence and role in the contaminated environment. Notably, it is observed that some microorganisms possess the capacity to survive in environments contaminated with toxic metals and even promote changes that minimize their toxicity. This suggests that they could be effective in bioremediation.
The Gram-positive bacteria in the group, according to
Table 5, were
Lysinibacillus sphaericus,
Rhodococcus wratislaviensis,
Bacillus thuringiensis, and
Microbacterium maritypicum. Gram-negative bacteria were identified as
Brevundimonas vesicularis,
Stenotrophonomas maltophila, Flavimonas oryzihabitans, Acinetobacter schindleri, and
Pseudomonas putida biotype B.
Bacillus thuringiensis is a prevalent soil bacterium; it is related to the presence of
Bacillus cereus.
Bacillus cereus is often seen as a biochemical indicator for concealed mineralisation [
31]. Despite its high similarity with
B. cereus,
B. thuringiensis is widely known for its insecticidal and nematicidal properties [
32,
33]. Bio-control agents derived from
B. thuringiensis are widely used in agriculture and medicine for eradicating pests [
34,
35,
36].
Bacillus species have been reported to be useful for the removal of toxic metals from contaminated soils [
37,
38], and their presence in soil has been linked with metal abundance [
39,
40].
Likewise,
Pseudomonas putida biotype B is also an organism that is common in polluted soil [
41,
42]. Though the organism may have some clinical implications, especially in pathogenicity, its resistive nature to pollution, especially to some toxic metals from aqueous solutions, may have influenced its presence in the leachate-contaminated soil [
43,
44].
Stenotrophomonas maltophila, a Gram-negative aerobic bacterium, was also isolated from the leachate-contaminated soil. It is pervasive in the environment [
45]. It inhabits the rhizosphere of plants such as potatoes, maize, wheat, oilseed, cucumber, and oat [
46,
47]. It can degrade xenobiotic compounds [
48,
49]. It can detoxify high molecular weight polycyclic aromatic hydrocarbons [
45,
50].
Lysinibacillus sphaericus is another Gram-positive bacterium that was isolated in this study. There have been reports of its existence in contaminated areas and wastewater [
51,
52]. Its significance in this work can be supported by [
53], which found that isolated
L. sphaericus have a bioremediation effect by acting as a metal binding site, and [
54] which showed the organism’s ability to reduce and oxidize manganese in water.
The aerobic, irregularly rod-shaped and Gram-positive bacteria
Rhodococcus wratislaviensis was also isolated from the leachate-contaminated site. It is predominant in soils [
55,
56]. Ref. [
57] reported that the
R. wratislaviensis strain that was isolated from forest soil was able to degrade nitroaromatic compounds. The isolation of the microbe in this work may be significant in terms of its ability of direct degradation or symbiotic potential to enhance bioremediation, as depicted in the studies [
58,
59].
It is important to note that despite the isolation of only nine bacteria species, the high resistive nature of microbes to leachate-contaminated sites in general, and toxic metals in particular, abounds. Therefore, considering this potential relevance of microorganisms, the present study further grouped the microbes into different classes, as shown in
Table 5. The groups of isolated bacteria were used for the bioaugmentation of leachate artificially contaminated soil for bioremediation tests.
3.2.2. Bioreduction of Metals
The concentrations of toxic metals were ascertained before and after treatment (
Table 6). Following bioremediation, the metal concentrations were determined based on the metal available in the soil after treatment (residual concentration).
Upon bioaugmentation of the leachate-contaminated soil, reductions in the concentrations of the toxic metals were observed across all days of biomonitoring and treatments (Groups A–D). The initial mean concentration of each metal in the leachate-contaminated soil was the same for all the treatments (Groups A–D). The residual mean concentrations following 100 days of biomonitoring are shown in
Table 6. The metals investigated in the present study are discussed individually across time and treatment groups.
The concentration of the toxic metal in the leachate-spiked soil at day 1 is denoted as “0”, followed by intermittent monitoring for the next 100 days at 20-day intervals. Therefore, the last monitoring was represented as “5”. According to the one-way analysis of variance (ANOVA) for Pb, there were significant differences between the initial day of contamination (day 1) and the final periods of monitoring. The level of significance was higher with the amended (inoculated) treatments than with the control experiment (
p = 0.002). This is probably due to the role played by the introduced microorganisms. This agrees with published findings that have highlighted the effectiveness of microbes in the bioremediation of metal-contaminated soils [
60,
61].
The highest removal of Pb was recorded in Group B (
Table 6). This was expected for Group C, where all 9 isolated bacteria species were utilized. The trend observed for Group B (with only three Gram-positive bacteria) may have occurred due to the interactions that exist among microbes when concentrations are manipulated. For instance,
B. thuringiensis, which is present in two of the inocula-amended microcosms (Groups B and C), has been found to possess a high capacity for toxic metal removal. Ref. [
37] reported that about 77% of Pb was removed from an extract medium of a mine tailing that contained 100 mg/L of Pb. In spite of the presence of
B. thuringiensis in Group C, the removal of Pb was more evident in treatments A and B; therefore, the order of Pb removal across the groups was D < C < A < B (
Table 6).
Figure 1 shows the degree to which Pb was removed from the different treatments. It is observed that while about 71% Pb was removed in Group B in 100 days, only about 42% was removed in the control experiment (Group D). Possibly, natural attenuation may have played a significant role with respect to the removal of Pb in Group D. After all, the soil was not sterilized or autoclaved; therefore, naturally inherent microbes may have some effect, though to a reduced extent in comparison to other treatments. Using a one-way ANOVA, F = 15.566, the difference in Pb removal between Groups B (treatment that showed highest removal) and D (control experiment showing least removal) was significant at
p (0.017) < 0.05. Therefore, a significant reduction in the concentration of Pb was observed with the introduction of inocula into the leachate-contaminated soil.
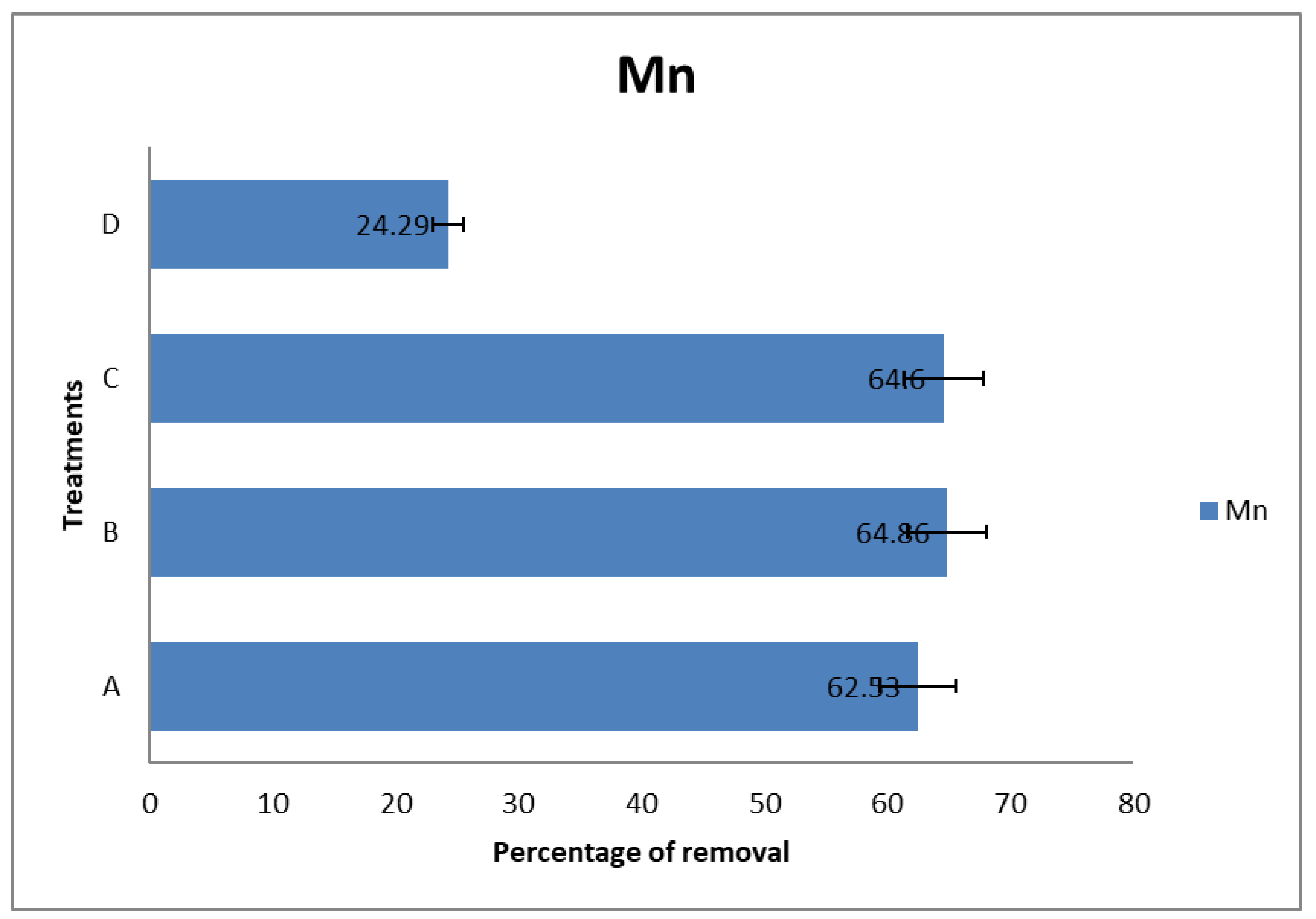
It can be observed that the amended soils (Groups A, B, and C) reduced the Mn content three times more than the non-amended soil (Group D) (
Figure 2;
Table 7). It can be inferred that microbial activities could initiate metabolic reactions favorable for the biodegradation of Mn. This suggestion is in consonance with the findings reported in [
62,
63].
In Groups A, B, C, and D, the mean residual concentrations of Cu were 0.07, 0.03, 0.09, and 0.11 (mg/kg) after 100 days of biomonitoring (
Table 7).
Figure 3 shows that the quantity of Cu that was removed in Group B is slightly greater than 86%, in comparison to 69%, 64%, and 52% recorded for treatments A, C, and D, respectively. According to the one-way ANOVA that was performed, there was a significant difference between B and D (
p = 0.003) and even with the other groups, for instance, AB (
p = 0.033) and BC (
p = 0.013). It is probable that
B. thuringiensis may have influenced the rate of reduction, especially in treatments B and C where it was present (
Table 6). This conclusion is supported by the findings in the study [
64], where the potential of
B. thuringiensis in the removal of Cu from soil contaminated with industrial effluent was reported. The optimal removal of Cu from contaminated soil recorded in Treatment B (
Figure 3,
Table 6) suggests that there may be a synergistic effect occurring among the selection that includes
B. thuringiensis,
L. sphaericus, and
R. wratislaviensis.
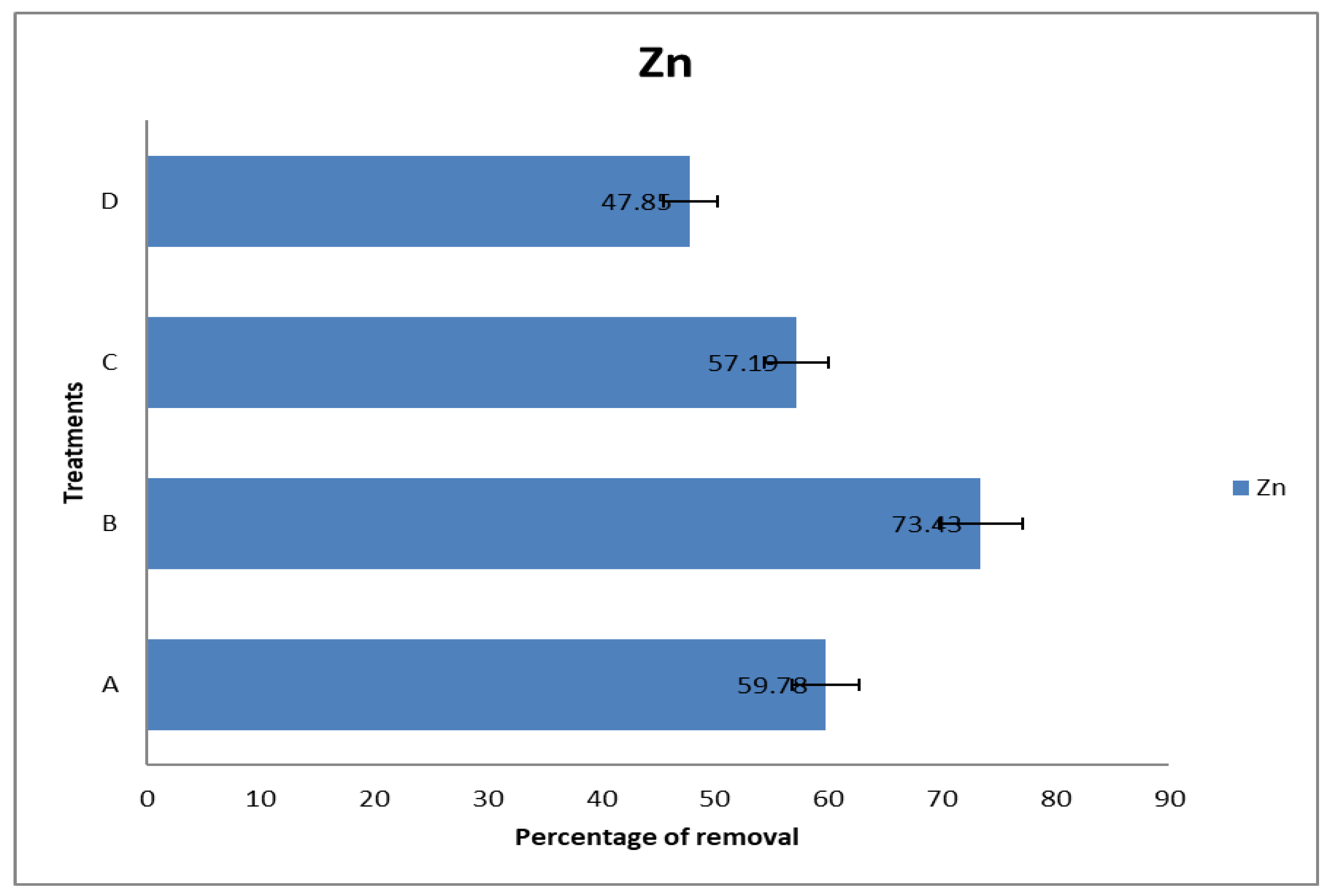
The highest removal of Zn (73.4%) was also recorded in Treatment B (
Figure 4). These bacteria species found across treatments A, B, and C, namely
B.
thuringiensis,
P. putida biotype B, and
M. maritypicum may have been significantly involved in the bioremediation of Zn. Several studies have found them to be effective in the removal of Zn. According to the study in [
65], about 14–68% of Zn was removed from polluted residue via
Microbacterium sp. Also, ref. [
66] reported the effective removal of Zn in the presence of
P. putida strains, MH3, MH6, and MH7. Similarly,
B. thuringiensis GDB-1 was able to remove about 64% of Zn in a metal-contaminated site [
37]. Therefore, a difference with respect to removal efficiency for Zn is observed between the inoculated microcosms and the control experiment (
Figure 4).
There was also better performance in terms of the removal of Fe in the amended treatments than in the control experiment, with the highest removal observed in Group B (
Figure 5;
Table 7).
In
Figure 6, Groups A and B exhibited the same degree of Cr reduction (81%) compared to 67% and 64% recorded for treatments C and D, respectively. This indicates that while the natural attenuation of Cr from leachate-contaminated soil is possible (Treatment D), improved removal can be achieved via bioaugmentation using the isolated microbes. The similarity observed between treatments A and B (
Figure 6) may be related to the presence of specific bacteria species that are known to be able to degrade Cr in contaminated media. Such microbes include
Stentrophomonas maltophilia [
67];
Pseudomonas sp. [
68,
69];
Bacillus sp. [
68,
70];
Acinetobacter sp. [
71];
Rhodococcus sp. [
69]; and
L. sphaericus [
72].
3.2.3. Rate Constant of Toxic Metal Removal
In the present study, we also studied the removal rate constants of the toxic metals (Pb, Mn, Cu, Zn, Fe, and Cr) per day in the different microcosms via the first-order kinetic model. This elucidates the removal ability of the microbial combinations daily, as it relates to each specific metal. This was then compared to the control experiment. It is evident from
Table 7 that the rate constant for the removal of Cu in the contaminated soil that was amended with only three bacteria species (
B. thuringiensis,
L. sphaericus, and
R. wratislaviensis) (Group B) within the 100-day study is the highest among the selected metals. It is possible that since the C-terminus of the S-layer protein SbpA of
L. sphaericus possess a hex-histidine tag (His
6—tag), this hex-histidine tag (metal binding property) is better expressed when it is combined with only
B. thuringiensis and
R. wratislaviensis (Group B) unlike with eight other species in Group C. Hence, the bioremediation edge was observed for Group B throughout the study (
Table 7).
The results from Group D show that some metals within the group may undergo natural remediation especially when contaminated soil is left undisturbed (
Table 7). Conversely, other metals in Group D that exhibited low removal rates in comparison to Groups A, B, and C could be a result of the inability of the pre-existing microbes to be self-sufficient regarding the bioremediation of contaminated soil. It is apparent that the amended soils showed greater bioremediation ability than the control experiment. However, the amendments prioritized the removal of toxic metals differently. For instance, Groups A and C prioritized Cr removal at 0.0167 day
−1 and 0.0108 day
−1, respectively, while Group B prioritized Cu removal (0.0212 day
−1) (
Table 7). Therefore, it can be inferred that complex microbial interactions exist within the different microcosms. Consequently, blending the microbes affords the maximum removal of toxic metals as is evident in Group B.
Specifically, with respect to the impact of leachate on the soil after the artificial contamination of the non-contaminated soil that was obtained from a garden, soil chemical characteristics such as soil pH and soil redox potential were monitored throughout the 100-day bioremediation period. Firstly, the pH of the soil from the garden at the University of Malaya, Malaysia, was found to be 7.52 before leachate contamination. After artificial contamination using leachate, at the onset of the bioremediation trials (day 0), the pH of the soils (Groups A-D) was found to be 8.2 (
Figure 7). It is worth mentioning that the leachate used for artificial contamination was obtained from JSL, which showed neutral–slightly alkaline pH, typical of an old landfill in its methanogenic phase. The pH of the artificially contaminated soil (8.2) was conducive to the growth of microbes. Regarding the pH changes in the soil during the bioremediation study, on day 20, the pH range for treatments A, B, C, and D was 7.65, 7.3, 7.47, and 7.68, whereas on day 100, the pH range was 8.07, 8.01, 8.11, and 8.21 for treatments A, B, C, and D, respectively (
Figure 7). The variation in soil pH could be due to binding of metals upon the introduction of bacterial inoculum. The optimal pH condition for microbial growth and maximum removal of chromium ion (Cr (VI)) was found to be in the pH range of 7–8 in the study in [
67]. This study corroborates the findings from our study where the bioreduction of hazardous metals and metalloids has also been observed at a similar pH range (neutral–slightly alkaline).
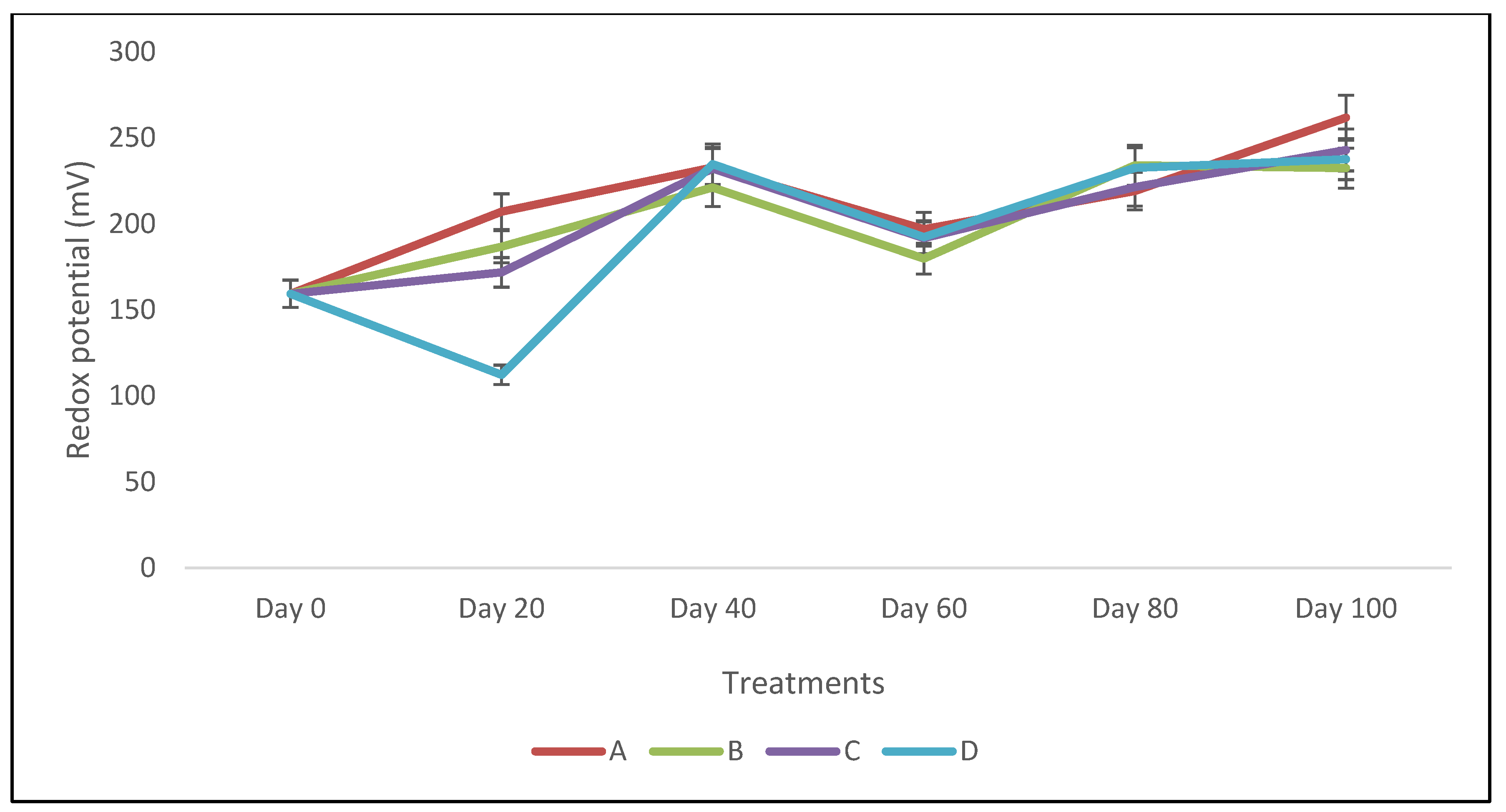
Also, in the course of the bioremediation process, the soil redox potential was evaluated for the soil(s) subjected to remediation.
Figure 8 shows that there is a mild but progressive increase in the redox potential (mV) from day 20 to day 100, except with a slight dip at day 60, for all the treatments. For instance, the redox potential ranged from 207.13 mV at day 20 to 261.73 at day 100, and from 186.73 mV at day 20 to 232.43 mV at day 100 for treatments A and B, respectively (
Figure 8). The increase in redox potential during bioremediation trials is indicative of oxidizing conditions [
73]. For example, a metal ion such as Fe
2+ under neutral pH and oxidizing condition could be converted to Fe
3+ in the form of iron hydroxide, Fe(OH)
3, which then precipitates out of the solution. The findings from this study for both soil pH and soil redox potential are indicative of neutral–slightly alkaline pH (pH 7–8) and oxidizing conditions. Both conditions enable the alteration of oxidation states of metal ions and the occurrence of metal transformations [
73,
74].
In addition, besides the analyses of the toxic metals’ concentrations, to further understand the interaction of the inoculated bacteria with the contaminated soil, the bacterial count was evaluated. During the 100-day remediation period, the bacterial count was performed every twenty days. The bacterial count over time for treatments A–D is presented in
Figure S1. It is crucial to remember that low inocula can hinder the survival of bioaugmented bacteria when considering the potential effects of other microbes. This study employed high inocula because bioaugmented bacteria colonization is a vital component of bioaugmentation success. To be precise, the inoculum in each treatment constituted roughly 3 × 10
9 CFU/g. Understanding the distribution of bacteria in a bioremediation setup is essential because the number of bacteria in the soil is indicative of the survival of microbes in metal-contaminated soils. The bacterial count showed a fluctuating trend throughout the course of the 100 days. For the first 20 days, the bacterial count ranged from 3.0 × 10
11 to 6.5 × 10
10 CFU/g for the amended treatments (
Figure S1). Ref. [
75] observed similar patterns in the number of bacteria throughout their bioremediation studies. The rise in the number of bacteria at days 40 and 60 is most likely due to the availability of nutrients in the soil. The advent of conditions that are conducive to cell duplication may have also contributed to an increase in the number of bacteria. As the experiment came to an end (Day 100), there was a noticeable decrease in the number of bacteria for Groups A–D (
Figure S1); depicting the extent to which natural and inocula bacteria bio-remove hazardous metals.
The findings in this study can be corroborated by the study in [
76]. In that study, indigenous bacteria that were isolated from a tannery sludge in India were able to remove Zn (69.9%) and Mn (78.4%) from metal-contaminated soil. In the present study, the His
6—tag at the C-terminus of the S-layer protein of
L. sphaericus may have enhanced its metal binding effect in the presence of
R. wratislaviensis and
B. thuringiensis. The results in the study in [
77] also lend credence to the findings of the present study.



 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}