
Circadian Rhythms of the Hypothalamus: From Function to Physiology


Abstract
:1. Introduction
2. Chronology of Clocks in the Hypothalamic Nuclei
3. A Regulatory Role for Hypothalamic Clocks in Feeding and Food-Anticipatory Activity
3.1. Feeding and the Circadian Clock in Neuronal Subtypes of the ARC
3.2. Feeding and the Circadian Clock in Neuronal Subtypes of the LH, PVN, DMH and VMH
4. Involvement of the Intrinsic Hypothalamic Clocks in Metabolic Regulation
4.1. Circadian Regulation of Metabolism by the ARC
4.2. Circadian Regulation of Metabolism by the PVN
4.3. Circadian Regulation of Metabolism by the VMH, DMH and LH
5. The Emerging Regulatory Role of Hypothalamic Clocks in Thirst Anticipation and Fluid Balance
6. The Circadian Link between Metabolism, Obesity and Sleep
7. Circadian Regulation of Metabolism by Neuroendocrine Hormones
7.1. Cortisol-Releasing Hormone (CRH)
7.2. Melatonin
7.3. Gut-Derived Polypeptides
8. The Role of Intrinsic Clocks of the Hypothalamus in Obesity
8.1. Time-Restricted Feeding
8.2. The Influences of Diet on the Hypothalamic Clocks
8.3. Sex-Specific Differences
9. The Role of Intrinsic Clocks of the Hypothalamus in Neurological Diseases and Disorders
9.1. Alzheimer’s Disease, Neuroinflammation and the Disruption of Sleep–Wake Cycles
9.2. Sundowning Syndrome
9.3. Seasonal Affective Disorder
9.4. Mood Disorders
10. Treatments and Therapeutics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| βOHB | β- hydroxybutyrate |
| AD | Alzheimer’s Disease |
| ADIPOQ | Adiponectin |
| AgRP | Agouti-Related Peptide |
| AMPK | 5′Adenosine Monophosphate-Activated Protein Kinase |
| ANS | Autonomic Nervous System |
| App-KI | Amyloid precursor protein knock-in |
| ARC | Arcuate Nucleus of the Hypothalamus |
| AVP | Arginine Vasopressin |
| BAT | Brown Adipose Tissue |
| BBB | Blood–Brain Barrier |
| BD | Bipolar Disorder |
| BDNF | Brain-Derived Neurotrophic Factor |
| BMAL1 | Brain Muscle Arnt-Like Protein 1 |
| Camk2a | Calcium/Calmodulin-Dependent Protein Kinase II Alpha |
| CART | Cocaine And Amphetamine-Regulated Transcript |
| CB1R | Cannabinoid Receptor 1 |
| CCG | Core Clock Genes |
| CCK | Cholecystokinin |
| CLOCK | Circadian Locomotor Output Cycles Protein Kaput |
| CK1δ | Casein Kinase 1 Delta |
| CK1ϵ | Casein Kinase 1 Epsilon |
| CNS | Central Nervous System |
| CORT | Corticotrophin |
| CREB | Camp Response Element-Binding Protein |
| CRH | Corticotrophin-Releasing Hormone |
| CRY | Cryptochrome |
| CRY1 | Cryptochrome 1 |
| CRY2 | Cryptochrome 2 |
| CTP1c | Carnitine Palmitoyltransferase 1c |
| dlPFC | Dorsal Lateral Prefrontal Cortex |
| DMH | Dorsal Medial Hypothalamus |
| dpHb | Dorsal Perihabenular Nucleus |
| eCB | Endocannabinoid |
| FAA | Food-Anticipatory Activity |
| FEO | Food-Entrainable Oscillator |
| FGF21 | Fibroblast Growth Factor 21 |
| GABA | Gamma-Aminobutyric Acid |
| GLP-1 | Glucagon-Like Peptide 1 |
| GRP | Gastrin-Releasing Peptide |
| HFD | High-Fat Diet |
| HPA | Hypothalamic–Pituitary Axis |
| ipRGCs | Induced Pluripotent Retinal Ganglion Cells |
| JNK | C-Jun N-Terminal Kinase |
| Kiss1 | Kisspeptin 1 |
| Lep | Leptin |
| LH | Lateral Hypothalamus |
| MCH | Melanin-Concentrating Hormone |
| MC4R | Melanocortin Receptor 4 |
| MPOA | Medial Preoptic Area |
| mTOR | Mammalian Target of Rapamycin |
| mTORc1 | Mammalian Target of Rapamycin Complex 1 |
| NAD+ | Nicotinamide Adenine Dinucleotide |
| NAM | Nicotinamide |
| NAMPT | Nicotinamide Phosphoribosyltransferase |
| NMN | Nicotinamide Mononucleotide |
| NOB | Nobiletin |
| NPAS2 | Neuronal PAS Domain Protein 2 |
| NPY | Neuropeptide Y |
| NR1D1 | Nuclear Receptor Subfamily 1 D Member 1 |
| NR1D2 | Nuclear Receptor Subfamily 1 D Member 2 |
| NREM | Non-Rapid Eye Movement |
| NTS | Nucleus Tractus Soilaris |
| OT | Oxytocin |
| OX | Orexin |
| PACAP | Pituitary Adenylate Cyclase-Activating Polypeptide |
| PB | Parabrachial Nucleus |
| PDYN | Prodynophin |
| PER | Period |
| PER1 | Period 1 |
| PER2 | Period 2 |
| PNS | Parasympathetic Nervous System |
| POMC | Proopiomelanocortin |
| PPARγ | Peroxisome Protein Activator Receptor Gamma |
| PVN | Paraventricular Nucleus |
| p75NTR | Neurotrophin Receptor P75 |
| REV-ERBα | Nr1d1 |
| REV-ERBβ | Nr1d2 |
| RF | Restricted Feeding |
| RHT | Retinohypothalamic Tract |
| RORs | Retinoic Acid Receptor-Related Orphan Receptors |
| SAD | Seasonal Affective Disorder |
| SCN | Suprachiasmatic Nucleus |
| SF1 | Steroidogenic Factor 1 |
| SIM1 | Single-Minded Homolog 1 |
| SIRT1 | Sirtuin 1 |
| SPVZ | Subparaventricular Zone |
| TRF | Time-Restricted Feeding |
| TRH | Thyrotropin-Releasing Hormone |
| TrkB | Tropomyosin Receptor Kinase B |
| TRPM2 | Transient Receptor Potential Cation Channel, Subfamily M, Member 2 |
| TF | Transcription Factor |
| TTLF | Transcription–Translation Feedback Loop |
| VLPO | Ventral Lateral Preoptic Area |
| VTA | Ventral Tegmental Area |
References
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [Green Version]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crumbley, C.; Wang, Y.; Kojetin, D.J.; Burris, T.P. Characterization of the core mammalian clock component, NPAS2, as a REV-ERBα/RORα target gene. J. Biol. Chem. 2010, 285, 35386–35392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhart-Hines, Z.; Feng, D.; Emmett, M.J.; Everett, L.J.; Loro, E.; Briggs, E.R.; Bugge, A.; Hou, C.; Ferrara, C.; Seale, P.; et al. The nuclear receptor Rev-erbα controls circadian thermogenic plasticity. Nature 2013, 503, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Kornmann, B.; Schaad, O.; Bujard, H.; Takahashi, J.S.; Schibler, U. System-driven and oscillator-dependent circadian transcription in mice with a conditionally active liver clock. PLoS Biol. 2007, 5, e34. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, Y.; Sahar, S.; Astarita, G.; Kaluzova, M.; Sassone-Corsi, P. Circadian Control of the NAD+ Salvage Pathway by CLOCK-SIRT1. Science 2009, 324, 654–657. [Google Scholar] [CrossRef]
- Bunger, M.K.; Wilsbacher, L.D.; Moran, S.M.; Clendenin, C.; Radcliffe, L.A.; Hogenesch, J.B.; Simon, M.C.; Takahashi, J.S.; Bradfield, C.A. Mop3 is an essential component of the master circadian pacemaker in mammals. Cell 2000, 103, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Baggs, J.E.; Price, T.S.; Ditacchio, L.; Panda, S.; Fitzgerald, G.A.; Hogenesch, J.B. Network features of the mammalian circadian clock. PLoS Biol. 2009, 7, e1000052. [Google Scholar] [CrossRef] [PubMed]
- Van Der Horst, G.T.J.; Muijtjens, M.; Kobayashi, K.; Takano, R.; Kanno, S.I.; Takao, M.; De Wit, J.; Verkerk, A.; Eker, A.P.M.; Van Leenen, D.; et al. Mammalian Cry1 and Cry2 are essential for maintenance of circadian rhythms. Nature 1999, 398, 627–630. [Google Scholar] [CrossRef]
- Bae, K.; Jin, X.; Maywood, E.S.; Hastings, M.H.; Reppert, S.M.; Weaver, D.R. Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock. Neuron 2001. [Google Scholar] [CrossRef] [Green Version]
- Musiek, E.S.; Xiong, D.D.; Holtzman, D.M. Sleep, circadian rhythms, and the pathogenesis of Alzheimer disease. Exp. Mol. Med. 2015, 47, e148. [Google Scholar] [CrossRef] [Green Version]
- Bokenberger, K.; Sjölander, A.; Dahl Aslan, A.K.; Karlsson, I.K.; Åkerstedt, T.; Pedersen, N.L. Shift work and risk of incident dementia: A study of two population-based cohorts. Eur. J. Epidemiol. 2018, 33, 977–987. [Google Scholar] [CrossRef] [Green Version]
- van der Vinne, V.; Martin Burgos, B.; Harrington, M.E.; Weaver, D.R. Deconstructing circadian disruption: Assessing the contribution of reduced peripheral oscillator amplitude on obesity and glucose intolerance in mice. J. Pineal Res. 2020, 69, 1–12. [Google Scholar] [CrossRef]
- Parsons, M.J.; Moffitt, T.E.; Gregory, A.M.; Goldman-Mellor, S.; Nolan, P.M.; Poulton, R.; Caspi, A. Social jetlag, obesity and metabolic disorder: Investigation in a cohort study. Int. J. Obes. 2015, 39, 842–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.; Ennaceur, A.; Cole, J.C.; Suh, C.K. Chronic jet lag produces cognitive deficits. J. Neurosci. 2000, 20, 1–5. [Google Scholar] [CrossRef]
- Reutrakul, S.; Knutson, K.L. Consequences of Circadian Disruption on Cardiometabolic Health. Sleep Med. Clin. 2015, 10, 455–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.Q.; Ansari, T.S.; McGuinness, O.P.; Wasserman, D.H.; Johnson, C.H. Circadian disruption leads to insulin resistance and obesity. Curr. Biol. 2013, 23, 372–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, S.M.; Honn, K.A.; Gaddameedhi, S.; Van Dongen, H.P.A. Shift Work: Disrupted Circadian Rhythms and Sleep—Implications for Health and Well-being. Curr. Sleep Med. Reports 2017, 3, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Borbely, A.A. A two process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar] [PubMed]
- Daan, S.; Beersma, D.G.M.; Borbely, A.A. Timing of human sleep: Recovery process gated by a circadian pacemaker. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1984, 15. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.J.; Czeisler, C.A. Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci. Lett. 1994, 166, 63–68. [Google Scholar] [CrossRef]
- Kohsaka, A.; Laposky, A.D.; Ramsey, K.M.; Estrada, C.; Joshu, C.; Kobayashi, Y.; Turek, F.W.; Bass, J. High-Fat Diet Disrupts Behavioral and Molecular Circadian Rhythms in Mice. Cell Metab. 2007, 6, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Navara, K.J.; Nelson, R.J. The dark side of light at night: Physiological, epidemiological, and ecological consequences. J. Pineal Res. 2007, 43, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, I.; Leinweber, B.; Brandenburger, M.; Oster, H. Circadian clock network desynchrony promotes weight gain and alters glucose homeostasis in mice. Mol. Metab. 2019, 30, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.P.; Hawkes, C.D. Increased Spontaneous activity and food intake produced in rats by removal of the frontal poles of the brain by. J. Neurol. Psychiatry 1939, 2, 231. [Google Scholar] [CrossRef] [Green Version]
- Richter, C.P. Biological clocks in medicine and psychiatry: Shock-phase hypothesis. Proc. Natl. Acad. Sci. USA 1960, 46, 1506–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, R.Y.; Lenn, N.J. A Retinohypothalamic Projection in the Rat. J. Comp. Neurol. 1972, 146, 1–14. [Google Scholar] [CrossRef]
- Stephan, F.K.; Zucker, I. Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc. Natl. Acad. Sci. USA 1972, 69, 1583–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, R.Y.; Eichler, V.B. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res. 1972, 42, 201–206. [Google Scholar] [CrossRef]
- Kawamura, H. Hypothalamic “Island” Containing the Suprachiasmatic Nucleus. Neurobiology 1979, 76, 5962–5966. [Google Scholar]
- Ralph, M.R.; Foster, R.G.; Davis, F.C. Transplanted Suprachiasmatic Nucleus Deternines Circadian Period. Science 1990, 247, 975–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, E.D.; Hermanstyne, T.; Smyllie, N.J.; Hastings, M.H. Regulating the suprachiasmatic nucleus (SCN) circadian clockwork: Interplay between cell- autonomous and circuit-level mechanisms. Cold Spring Harb. Perspect. Biol. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Allaman-Pillet, N.; Roduit, R.; Oberson, A.; Abdelli, S.; Ruiz, J.; Beckmann, J.S.; Schorderet, D.F.; Bonny, C. Circadian regulation of islet genes involved in insulin production and secretion. Mol. Cell. Endocrinol. 2004, 226, 59–66. [Google Scholar] [CrossRef]
- Keller, M.; Mazuch, J.; Abraham, U.; Eom, G.D.; Herzog, E.D.; Volk, H.D.; Kramer, A.; Maier, B. A circadian clock in macrophages controls inflammatory immune responses. Proc. Natl. Acad. Sci. USA 2009, 106, 21407–21412. [Google Scholar] [CrossRef] [Green Version]
- Schmitt & Segert NIH Public Access. Bone 2008, 23, 1–7. [CrossRef]
- Shostak, A.; Meyer-Kovac, J.; Oster, H. Circadian regulation of lipid mobilization in white adipose tissues. Diabetes 2013, 62, 2195–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Zhang, Y.; Zhang, F.; Xia, Y.; Liu, J.; Huang, R.; Wang, Y.; Hu, Y.; Wu, J.; Dai, C.; et al. CLOCK/BMAL1 regulates circadian change of mouse hepatic insulin sensitivity by SIRT1. Hepatology 2014, 59, 2196–2206. [Google Scholar] [CrossRef]
- Carrasco-Benso, M.P.; Rivero-Gutierrez, B.; Lopez-Minguez, J.; Anzola, A.; Diez-Noguera, A.; Madrid, J.A.; Lujan, J.A.; Martínez-Augustin, O.; Scheer, F.A.J.L.; Garaulet, M. Human adipose tissue expresses intrinsic circadian rhythm in insulin sensitivity. FASEB J. 2016, 30, 3117–3123. [Google Scholar] [CrossRef] [Green Version]
- Boucher, H.; Vanneaux, V.; Domet, T.; Parouchev, A.; Larghero, J. Circadian clock genes modulate human bone marrow mesenchymal stem cell differentiation, migration and cell cycle. PLoS ONE 2016, 11, e0146674. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, W.; Liu, C. Integration of peripheral circadian clock and energy metabolism in metabolic tissues. J. Mol. Cell Biol. 2020, 12, 481–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.; Yamazaki, S.; Lowrey, P.L.; Shimomura, K.; Ko, C.H.; Buhr, E.D.; Siepka, S.M.; Hong, H.; Oh, W.J.; Yoo, O.J.; et al. PERIOD2: LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc. Natl. Acad. Sci. USA 2004, 101, 5339–5346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cedernaes, J.; Waldeck, N.; Bass, J. Neurogenetic basis for circadian regulation of metabolism by the hypothalamus. Genes Dev. 2019, 33, 1136–1158. [Google Scholar] [CrossRef] [Green Version]
- Greco, C.M.; Corsi, P.S. Circadian blueprint of metabolic pathways in the brain. Nat. Rev. Neurosci. 2019, 20, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Begemann, K.; Neumann, A.M.; Oster, H. Regulation and function of extra-SCN circadian oscillators in the brain. Acta Physiol. 2020, 229, 1–14. [Google Scholar] [CrossRef]
- Paul, J.R.; Davis, J.A.; Goode, L.K.; Becker, B.K.; Fusilier, A.; Meador-Woodruff, A.; Gamble, K.L. Circadian regulation of membrane physiology in neural oscillators throughout the brain. Eur. J. Neurosci. 2020, 51, 109–138. [Google Scholar] [CrossRef] [Green Version]
- Stephan, F.K.; Zucker, I. Circadian Rhythms. Science 1972, 69, 1583–1586. [Google Scholar]
- Stephan, F.K. The “other” circadian system: Food as a zeitgeber. J. Biol. Rhythms 2002, 17, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Morin, L.P. The circadian visual system. Brain Res. Rev. 1994, 19, 102–127. [Google Scholar] [CrossRef]
- Abrahamson, E.E.; Moore, R.Y. Suprachiasmatic nucleus in the mouse: Retinal innervation, intrinsic organization and efferent projections. Brain Res. 2001, 916, 172–191. [Google Scholar] [CrossRef]
- Swanson, L.W.; Cowan, W.M. The efferent connections of the suprachiasmatic nucleus of the hypothalamus. J. Comp. Neurol. 1975. [Google Scholar] [CrossRef] [PubMed]
- Berk, M.L.; Finkelstein, J.A. An autoradiographic determination of the efferent projections of the suprachiasmatic nucleus of the hypothalamus. Brain Res. 1981. [Google Scholar] [CrossRef]
- Kriegsfeld, L.J.; Leak, R.K.; Yackulic, C.B.; Sauter, J.L.E.; Silver, R.A.E. Organization of Suprachiasmatic Nucleus Projections in Syrian Hamsters (Mesocricetus auratus): An Anterograde and Retrograde Analysis. J. Comp. Neurol. 2004, 379, 361–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnick, I.; Krishtal, O.A.; Colmers, W.F. Integration of energy homeostasis and stress by parvocellular neurons in rat hypothalamic paraventricular nucleus. J. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Billington, C.J.; Briggs, J.E.; Harker, S.; Grace, M.; Levine, A.S. Neuropeptide Y in hypothalamic paraventricular nucleus: A center coordinating energy metabolism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1994, 266. [Google Scholar] [CrossRef]
- Satoh, N.; Ogawa, Y.; Katsuura, G.; Hayase, M.; Tsuji, T.; Imagawa, K.; Yoshimasa, Y.; Nishi, S.; Hosoda, K.; Nakao, K. The arcuate nucleus as a primary site of satiety effect of leptin in rats. Neurosci. Lett. 1997, 224, 149–152. [Google Scholar] [CrossRef]
- Padilla, S.L.; Perez, J.G.; Ben-Hamo, M.; Johnson, C.W.; Sanchez, R.E.A.; Bussi, I.L.; Palmiter, R.D.; de la Iglesia, H.O. Kisspeptin Neurons in the Arcuate Nucleus of the Hypothalamus Orchestrate Circadian Rhythms and Metabolism. Curr. Biol. 2019, 29, 592–604. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.W.; Donato, J.; Berglund, E.D.; Choi, Y.H.; Kohno, D.; Elias, C.F.; DePinho, R.A.; Elmquist, J.K. FOXO1 in the ventromedial hypothalamus regulates energy balance. J. Clin. Investig. 2012. [Google Scholar] [CrossRef]
- Çakir, I.; Perello, M.; Lansari, O.; Messier, N.J.; Vaslet, C.A.; Nillni, E.A. Hypothalamic Sirt1 regulates food intake in a rodent model system. PLoS ONE 2009. [Google Scholar] [CrossRef] [Green Version]
- Leibowitz, S.F. Reciprocal hunger-regulating circuits involving alpha- and beta-adrenergic receptors located, respectively, in the ventromedial and lateral hypothalamus. Proc. Natl. Acad. Sci. USA 1970, 67, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Berthoud, H.R.; Münzberg, H. The lateral hypothalamus as integrator of metabolic and environmental needs: From electrical self-stimulation to opto-genetics. Physiol. Behav. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.; Harrold, J.A.; Cutler, D.J. The hypothalamus and the regulation of energy homeostasis: Lifting the lid on a black box. Proc. Nutr. Soc. 2000, 59, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Arrigoni, E.; Chee, M.J.S.; Fuller, P.M. To eat or to sleep: That is a lateral hypothalamic question. Neuropharmacology 2019, 154, 34–49. [Google Scholar] [CrossRef]
- Nader, N.; Chrousos, G.P.; Kino, T. Interactions of the circadian CLOCK system and the HPA axis. Trends Endocrinol. Metab. 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, L.W.; Kuypers, H.G.J.M. The paraventricular nucleus of the hypothalamus: Cytoarchitectonic subdivisions and organization of projections to the pituitary, dorsal vagal complex, and spinal cord as demonstrated by retrograde fluorescence double-labeling methods. J. Comp. Neurol. 1980, 194, 555–570. [Google Scholar] [CrossRef]
- Chou, T.C.; Scammell, T.E.; Gooley, J.J.; Gaus, S.E.; Saper, C.B.; Lu, J. Critical Role of Dorsomedial Hypothalamic Nucleus in a Wide Range of Behavioral Circadian Rhythms. J. Neurosci. 2003, 23, 10691–10702. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, Y.H.; Chou, T.C.; Gaus, S.E.; Elmquist, J.K.; Shiromani, R.; Saper, C.B. Contrasting effects of ibotenate lesions of the paraventricular nucleus and subparaventricular zone on sleep-wake cycle and temperature regulation. J. Neurosci. 2001, 21, 4864–4874. [Google Scholar] [CrossRef] [PubMed]
- Asala, S.A.; Okano, Y.; Honda, K.; Inoué, S. Effects of medial preoptic area lesions on sleep and wakefulness in unrestrained rats. Neurosci. Lett. 1990, 114, 300–304. [Google Scholar] [CrossRef]
- Mondino, A.A.; Hambrecht-wiedbusch, V.; Li, D.; York, A.K. Glutamatergic neurons in the preoptic hypothalamus promote wakefulness, destabilize NREM sleep, suppress REM sleep, and regulate cortical dynamics. bioRxiv 2020. [Google Scholar] [CrossRef]
- Tosini, G.; Menaker, M. Circadian Rhythms in Cultured Mammalian Retina. Science 1996, 272, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Herzog, E.D.; Yamazaki, S.; Straume, M.; Tei, H.; Sakaki, Y.; Menaker, M.; Block, G.D. Circadian rhythms in isolated brain regions. J. Neurosci. 2002, 22, 350–356. [Google Scholar] [CrossRef]
- Anand, B.; Brobeck, J. Hypothalamic control of food intake in rats and cats * Marked variations in food intake have been described in various species following injury to certain parts of the hypothalamus, including an thalamus, especially lesions in or ventrolateral to the ve. Yale J. Biol. Med. 1951, 24, 123–140. [Google Scholar] [PubMed]
- Richter, C.P. A Behavioristic Study of the Activity of the Rat. Comp. Psychol. Monogr. 1922, 1, 56. [Google Scholar]
- Aschoff, J.; Von Goetz, C. Effects of Feeding Cycles on Circadian Rhythms in Squirrel Monkeys. J. Biol. Rhythm. 1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistlberger, R.E. Circadian food-anticipatory activity: Formal models and physiological mechanisms. Neurosci. Biobehav. Rev. 1994, 18, 171–195. [Google Scholar] [CrossRef]
- Meyer-Lohmann, J. Über den Einfluß täglicher Futtergaben auf die 24-Stunden-Periodik der lokomotorischen Aktivität weißer Mäuse. Pflugers Arch. Gesamte Physiol. Menschen Tiere 1955. [Google Scholar] [CrossRef]
- Edmonds, S.C.; Adler, N.T. Food and light as entrainers of circadian running activity in the rat. Physiol. Behav. 1977, 18, 915–919. [Google Scholar] [CrossRef]
- Mendoza, J.; Graff, C.; Dardente, H.; Pevet, P.; Challet, E. Feeding cues alter clock gene oscillations and photic responses in the suprachiasmatic nuclei of mice exposed to a light/dark cycle. J. Neurosci. 2005, 25, 1514–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumo, M.; Pejchal, M.; Schook, A.C.; Lange, R.P.; Walisser, J.A.; Sato, T.R.; Wang, X.; Bradfield, C.A.; Takahashi, J.S. Differential effects of light and feeding on circadian organization of peripheral clocks in a forebrain Bmal1 mutant. Elife 2014, 3, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mieda, M.; Sakurai, T. Bmal1 in the nervous system is essential for normal adaptation of circadian locomotor activity and food intake to periodic feeding. J. Neurosci. 2011, 31, 15391–15396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puelles, L.; Rubenstein, J.L.R. Forebrain gene expression domains and the evolving prosomeric model. Trends Neurosci. 2003, 26, 469–476. [Google Scholar] [CrossRef]
- Shimogori, T.; Lee, D.A.; Miranda-Angulo, A.; Yang, Y.; Wang, H.; Jiang, L.; Yoshida, A.C.; Kataoka, A.; Mashiko, H.; Avetisyan, M.; et al. A genomic atlas of mouse hypothalamic development. Nat. Neurosci. 2010, 13, 767–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mieda, M.; Hasegawa, E.; Kessaris, N.; Sakurai, T. Fine-tuning circadian rhythms: The importance of Bmal1 expression in the ventral forebrain. Front. Neurosci. 2017, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Delezie, J.; Dumont, S.; Sandu, C.; Reibel, S.; Pevet, P.; Challet, E. Rev-erbα in the brain is essential for circadian food entrainment. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef]
- Cho, H.; Zhao, X.; Hatori, M.; Yu, R.T.; Barish, G.D.; Lam, M.T.; Chong, L.; Ditacchio, L.; Atkins, A.R.; Glass, C.K.; et al. HHS Public Access. Nature 2012, 485, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Mang, G.M.; La Spada, F.; Emmenegger, Y.; Chappuis, S.; Ripperger, J.A.; Albrecht, U.; Franken, P. Altered sleep homeostasis in REV-ERBα knockout mice. Sleep 2016, 39, 589–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudley, C.A.; Erbel-Sieler, C.; Estill, S.J.; Reick, M.; Franken, P.; Pitts, S.N.; McKnight, S.L. Altered patterns of sleep and behavioral adaptability in NPAS2-deficient mice. Science 2003, 301, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.M.; Hsu, C.T.; Gunapala, K.M.; Parfyonov, M.; Chang, C.H.; Mistlberger, R.E.; Steele, A.D. Behavioral and neural correlates of acute and scheduled hunger in C57BL/6 mice. PLoS ONE 2014, 9, e95990. [Google Scholar] [CrossRef]
- Wang, D.; Opperhuizen, A.L.; Reznick, J.; Turner, N.; Su, Y.; Cooney, G.J.; Kalsbeek, A. Effects of feeding time on daily rhythms of neuropeptide and clock gene expression in the rat hypothalamus. Brain Res. 2017, 1671, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Sellix, M.T.; Egli, M.; Poletini, M.O.; Mckee, D.N.T.; Fitch, C.A.; Freeman, M.E. Author Manuscript anatomical and functional characterization of. Am. J. Physiol. 2007, 290, 1–30. [Google Scholar]
- Guilding, C.; Hughes, A.T.; Brown, T.M.; Namvar, S.; Piggins, H.D. A riot of rhythms: Neuronal and glial circadian oscillators in the mediobasal hypothalamus. Mol. Brain 2009, 2, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.; Knight, Z.A.; Friedman, J.M. Ablation of AgRP neurons impairs adaption to restricted feeding. Mol. Metab. 2014, 3, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Cedernaes, J.; Huang, W.; Ramsey, K.M.; Waldeck, N.; Cheng, L.; Marcheva, B.; Omura, C.; Kobayashi, Y.; Peek, C.B.; Levine, D.C.; et al. Transcriptional Basis for Rhythmic Control of Hunger and Metabolism within the AgRP Neuron. Cell Metab. 2019, 29, 1078–1091. [Google Scholar] [CrossRef]
- Wiater, M.F.; Mukherjee, S.; Li, A.J.; Dinh, T.T.; Rooney, E.M.; Simasko, S.M.; Ritter, S. Circadian integration of sleep-wake and feeding requires NPY receptorexpressing neurons in the mediobasal hypothalamus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, 1569–1583. [Google Scholar] [CrossRef]
- Wiater, M.F.; Li, A.J.; Dinh, T.T.; Jansen, H.T.; Ritter, S. Leptin-sensitive neurons in the arcuate nucleus integrate activity and temperature circadian rhythms and anticipatory responses to food restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, 949–960. [Google Scholar] [CrossRef] [Green Version]
- Henry, F.E.; Sugino, K.; Tozer, A.; Branco, T.; Sternson, S.M. Cell type-specific transcriptomics of hypothalamic energy-sensing neuron responses to weight-loss. Elife 2015, 4, 1–30. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, L.D.; Roa, S.L.; Bueno, A.C.; Coeli-Lacchini, F.B.; Martins, C.S.; Uchoa, E.T.; Antunes-Rodrigues, J.; Elias, L.L.; Elias, P.C.; Moreira, A.C.; et al. Restricted feeding schedules modulate in a different manner the expression of clock genes in rat hypothalamic nuclei. Front. Neurosci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Elmquist, J.K.; Coppari, R.; Balthasar, N.; Ichinose, M.; Lowell, B.B. Identifying hypothalamic pathways controlling food intake, body weight, and glucose homeostasis. J. Comp. Neurol. 2005, 493, 63–71. [Google Scholar] [CrossRef]
- Podyma, B.; Johnson, D.A.; Sipe, L.; Remcho, T.P.; Battin, K.; Liu, Y.; Yoon, S.O.; Deppmann, C.D.; Güler, A.D. The p75 neurotrophin receptor in AgRP neurons is necessary for homeostatic feeding and food anticipation. Elife 2020, 9, 1–19. [Google Scholar] [CrossRef]
- Baeza-raja, B.; Eckel-mahan, K.; Zhang, L.; Vagena, E.; Tsigelny, I.F.; Sassone-corsi, P.; Pta, L.J. p75 Neurotrophin Receptor Is a Clock Gene That Regulates Oscillatory Components of Circadian and Metabolic Networks. J. Neurosci. 2013, 33, 10221–10234. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Inoki, K.; Mu, H.; Opland, D.; Faouzi, M.; Villanueva, E.C.; Ikenoue, T.; Kwiatkowski, D.; Macdougald, O.A.; Myers, M.G.; et al. Critical Role for Hypothalamic mTOR Activity in Energy Balance. Cell Metab. 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cota, D.; Proulx, K.; Blake Smith, K.A.; Kozma, S.C.; Thomas, G.; Woods, S.C.; Seeley, R.J. Hypothalamic mTOR signaling regulates food intake. Science 2006, 312, 927–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albert, V.; Cornu, M.; Hall, M.N. mTORC1 signaling in Agrp neurons mediates circadian expression of Agrp and NPY but is dispensable for regulation of feeding behavior. Biochem. Biophys. Res. Commun. 2015, 464, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Haissaguerre, M.; Ferrière, A.; Simon, V.; Saucisse, N.; Dupuy, N.; André, C.; Clark, S.; Guzman-Quevedo, O.; Tabarin, A.; Cota, D. mTORC1-dependent increase in oxidative metabolism in POMC neurons regulates food intake and action of leptin. Mol. Metab. 2018, 12, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Woodworth, H.L.; Leinninger, G.M. To ingest or rest? Specialized roles of lateral hypothalamic area neurons in coordinating energy balance. Front. Syst. Neurosci. 2015, 9, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venner, A.; Karnani, M.M.; Gonzalez, J.A.; Jensen, L.T.; Fugger, L.; Burdakov, D. Orexin neurons as conditional glucosensors: Paradoxical regulation of sugar sensing by intracellular fuels. J. Physiol. 2011, 589, 5701–5708. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C.; Schwartz, M.W.; Baskin, D.G.; Seeley, R.J. Food I Ntake and the R Egulation of body weight. Annu. Rev. Psychol. 2000, 51, 255–277. [Google Scholar] [CrossRef] [Green Version]
- Nakamachi, T.; Matsuda, K.; Maruyama, K.; Miura, T.; Uchiyama, M.; Funahashi, H.; Sakurai, T.; Shioda, S. Regulation by orexin of feeding behaviour and locomotor activity in the goldfish. J. Neuroendocrinol. 2006, 18, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Mistlberger, R.E.; Rusak, B. Food-anticipatory circadian rhythms in rats with paraventricular and lateral hypothalamic ablations. J. Biol. Rhythm. 1988, 3, 277–291. [Google Scholar] [CrossRef]
- Mieda, M.; Williams, S.C.; Sinton, C.M.; Richardson, J.A.; Sakurai, T.; Yanagisawa, M. Orexin neurons function in an efferent pathway of a food-entrainable circadian oscillator in eliciting food-anticipatory activity and wakefulness. J. Neurosci. 2004, 24, 10493–10501. [Google Scholar] [CrossRef]
- Hill, J. PVN pathways controlling energy homeostasis. Indian J. Endocrinol. Metab. 2012. [Google Scholar] [CrossRef] [PubMed]
- Saeb-Parsy, K.; Lombardelli, S.; Khan, F.Z.; McDowall, K.; Au-Yong, I.T.H.; Dyball, R.E.J. Neural connections of hypothalamic neuroendocrine nuclei in the rat. J. Neuroendocrinol. 2000. [Google Scholar] [CrossRef] [PubMed]
- Xi, D.; Roizen, J.; Lai, M.; Gandhi, N.; Kublaoui, B. Paraventricular nucleus sim1 neuron ablation mediated obesity is resistant to high fat diet. PLoS ONE 2013, 8, e81087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tousson, E.; Meissl, H. Suprachiasmatic Nuclei Grafts Restore the Circadian Rhythm in the Paraventricular Nucleus of the Hypothalamus. J. Neurosci. 2004, 24, 2983–2988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, D.; Mukai, Y.; Hung, C.J.; Chowdhury, S.; Sugiyama, T.; Yamanaka, A. Neuron The mammalian circadian pacemaker regulates wakefulness via CRF neurons in the paraventricular nucleus of the hypothalamus. Sci. Adv. 2020, 6, eabd0384. [Google Scholar] [CrossRef]
- Sims, J.S.; Lorden, J.F. Effect of paraventricular nucleus lesions on body weight, food intake and insulin levels. Behav. Brain Res. 1986, 22, 265–281. [Google Scholar] [CrossRef]
- Yoshioka, K.; Yoshida, T.; Kawara, H. Lesions of the Hypothalamic Norepinephrine Paraventricular Turnover in Rats. Endocrinol. Jpn. 1989, 36, 187–194. [Google Scholar]
- Kalsbeek, A.; Palm, I.F.; La Fleur, S.E.; Scheer, F.A.J.L.; Perreau-Lenz, S.; Ruiter, M.; Kreier, F.; Cailotto, C.; Buijs, R.M. SCN outputs and the hypothalamic balance of life. J. Biol. Rhythms 2006, 21, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caba, M.; Huerta, C.; Meza, E.; Hernández, M.; Rovirosa-Hernández, M.J. Oxytocinergic Cells of the Hypothalamic Paraventricular Nucleus Are Involved in Food Entrainment. Front. Neurosci. 2020, 14, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buijs, R.M.; Kalsbeek, A. Hypothalamic integration of central and peripheral clocks. Nat. Rev. Neurosci. 2001, 2, 1–6. [Google Scholar] [CrossRef]
- Buijs, R.M.; Van Eden, C.G. The integration of stress by the hypothalamus, amygdala and prefrontal cortex: Balance between the autonomic nervous system and the neuroendocrine system. Prog. Brain Res. 2000, 126, 117–132. [Google Scholar] [CrossRef]
- Li, M.M.; Madara, J.C.; Steger, J.S.; Conley, N.J.; Garfield, A.S.; Lowell, B.B.; Li, M.M.; Madara, J.C.; Steger, J.S.; Krashes, M.J.; et al. The Paraventricular Hypothalamus Regulates Satiety and Prevents Obesity via Two Genetically Article The Paraventricular Hypothalamus Regulates Satiety and Prevents Obesity via Two Genetically Distinct Circuits. Neuron 2019, 102, 653–667.e6. [Google Scholar] [CrossRef] [PubMed]
- Xi, D.; Gandhi, N.; Lai, M.; Kublaoui, B.M. Ablation of Sim1 neurons causes obesity through hyperphagia and reduced energy expenditure. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Garza, J.C.; Kim, C.S.; Liu, J.; Zhang, W.; Lu, X.Y. Adeno-associated virus-mediated knockdown of melanocortin-4 receptor in the paraventricular nucleus of the hypothalamus promotes high-fat diet-induced hyperphagia and obesity. J. Endocrinol. 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalra, S.P.; Dube, M.G.; Sahu, A.; Phelps, C.P.; Kalra, P.S. Neuropeptide Y secretion increases in the paraventricular nucleus in association with increased appetite for food. Proc. Natl. Acad. Sci. USA 1991, 88, 10931–10935. [Google Scholar] [CrossRef] [Green Version]
- Santoso, P.; Nakata, M.; Ueta, Y.; Yada, T. CNS control of metabolism: Suprachiasmatic vasopressin to paraventricular oxytocin neurocircuit in the hypothalamus relays light reception to inhibit feeding behavior. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E478–E488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mieda, M.; Williams, S.C.; Richardson, J.A.; Tanaka, K.; Yanagisawa, M. The dorsomedial hypothalamic nucleus as a putative food-entrainable circadian pacemaker. Proc. Natl. Acad. Sci. USA 2006, 103, 12150–12155. [Google Scholar] [CrossRef] [Green Version]
- Gooley, J.J.; Schomer, A.; Saper, C.B. The dorsomedial hypothalamic nucleus is critical for the expression of food-entrainable circadian rhythms. Nat. Neurosci. 2006, 9, 398–407. [Google Scholar] [CrossRef]
- Fuller, P.M.; Lu, J.; Saper, C.B. Food-Entrainable Circadian Rhythms. Science 2008, 320, 1074–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landry, G.J.; Yamakawa, G.R.; Webb, I.C.; Mear, R.J.; Mistlberger, R.E. The dorsomedial hypothalamic nucleus is not necessary for the expression of circadian food-anticipatory activity in rats. J. Biol. Rhythm. 2007, 22, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Mistlberger, R.E.; Buijs, R.M.; Challet, E.; Escobar, C.; Landry, G.J.; Kalsbeek, A.; Pevet, P.; Shibata, S. Food anticipation in Bmal1-/- and AAV-Bmal1 rescued mice: A reply to Fuller. J. Circadian Rhythm. 2009, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Landry, G.J.; Simon, M.M.; Webb, I.C.; Mistlberger, R.E. Persistence of a behavioral food-anticipatory circadian rhythm following dorsomedial hypothalamic ablation in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Galvan, G.; Yi, C.X.; Van Der Vliet, J.; Jhamandas, J.H.; Panula, P.; Angeles-Castellanos, M.; Del Carmen Basualdo, M.; Escobar, C.; Buijs, R.M. Interaction between hypothalamic dorsomedial nucleus and the suprachiasmatic nucleus determines intensity of food anticipatory behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 5813–5818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, G.Y.; Kinney, C.E.; An, J.J.; Xu, B. TrkB-expressing neurons in the dorsomedial hypothalamus are necessary and sufficient to suppress homeostatic feeding. Proc. Natl. Acad. Sci. USA 2019, 116, 3256–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otgon-Uul, Z.; Suyama, S.; Onodera, H.; Yada, T. Optogenetic activation of leptin- and glucose-regulated GABAergic neurons in dorsomedial hypothalamus promotes food intake via inhibitory synaptic transmission to paraventricular nucleus of hypothalamus. Mol. Metab. 2016. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.C.; Kurrasch, D.M.; Liang, J.K.; Ingraham, H.A. Genetic labeling of steroidogenic factor-1 (SF-1) neurons in mice reveals ventromedial nucleus of the hypothalamus (VMH) circuitry beginning at neurogenesis and development of a separate non-SF-1 neuronal cluster in the ventrolateral VMH. J. Comp. Neurol. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, J.D.; Swanson, L.W. Connections of the lateral hypothalamic area juxtadorsomedial region in the male rat. J. Comp. Neurol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Minokoshi, Y.; Haque, M.S.; Shimazu, T. Microinjection of leptin into the ventromedial hypothalamus increases glucose uptake in peripheral tissues in rats. Diabetes 1999. [Google Scholar] [CrossRef]
- Choi, Y.H.; Fujikawa, T.; Lee, J.; Reuter, A.; Kim, K.W. Revisiting the ventral medial nucleus of the hypothalamus: The roles of SF-1 neurons in energy homeostasis. Front. Neurosci. 2013. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, H.; Zigman, J.M.; Ye, C.; Lee, C.E.; McGovern, R.A.; Tang, V.; Kenny, C.D.; Christiansen, L.M.; White, R.D.; Edelstein, E.A.; et al. Leptin directly activates SF1 neurons in the VMH, and this action by leptin is required for normal body-weight homeostasis. Neuron 2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, D.T. Ventromedial hypothalamic lesions abolish food-shifted circadian adrenal and temperature rhythinicity. Endocrinology 1980, 106, 649–654. [Google Scholar] [CrossRef]
- Mistlberger, R.E.; Rechtschaffen, A. Recovery of anticipatory activity to restricted feeding in rats with ventromedial hypothalamic lesions. Physiol. Behav. 1984, 33, 227–235. [Google Scholar] [CrossRef]
- Davidson, A.J.; Poole, A.S.; Yamazaki, S.; Menaker, M. Is the food-entrainable circadian oscillator in the digestive system? Genes, Brain Behav. 2003, 2, 32–39. [Google Scholar] [CrossRef]
- Carneiro, B.T.S.; Araujo, J.F. The food-entrainable oscillator: A network of interconnected brain structures entrained by humoral signals? Chronobiol. Int. 2009, 26, 1273–1289. [Google Scholar] [CrossRef]
- Pendergast, J.S.; Yamazaki, S. The Mysterious Food-Entrainable Oscillator: Insights from Mutant and Engineered Mouse Models. J. Biol. Rhythm. 2018, 33, 458–474. [Google Scholar] [CrossRef]
- Chavan, R.; Feillet, C.; Costa, S.S.F.; Delorme, J.E.; Okabe, T.; Ripperger, J.A.; Albrecht, U. Liver-derived ketone bodies are necessary for food anticipation. Nat. Commun. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, A.H.; Koch, C.E.; Kiehn, J.T.; Schmidt, C.X.; Oster, H. An adipokine feedback regulating diurnal food intake rhythms in mice. Elife 2020, 9, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowrey, P.L.; Shimomura, K.; Antoch, M.P.; Yamazaki, S.; Zemenides, P.D.; Ralph, M.R.; Menaker, M.; Takahashi, J.S. Positional syntenic cloning and functional characterization of the mammalian circadian mutation tau. Science 2000, 288, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamia, K.A.; Storch, K.F.; Weitz, C.J. Physiological significance of a peripheral tissue circadian clock. Proc. Natl. Acad. Sci. USA 2008, 105, 15172–15177. [Google Scholar] [CrossRef] [Green Version]
- Sadacca, L.A.; Lamia, K.A.; DeLemos, A.S.; Blum, B.; Weitz, C.J. An intrinsic circadian clock of the pancreas is required for normal insulin release and glucose homeostasis in mice. Diabetologia 2011, 54, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Seale, P.; Conroe, H.M.; Estall, J.; Kajimura, S.; Frontini, A.; Ishibashi, J.; Cohen, P.; Cinti, S.; Spiegelman, B.M. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J. Clin. Investig. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, F.F.; Kleiner, S.; Douris, N.; Fox, E.C.; Mepani, R.J.; Verdeguer, F.; Wu, J.; Kharitonenkov, A.; Flier, J.S.; Maratos-Flier, E.; et al. FGF21 regulates PGC-1α and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012, 26, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingenspor, M. Cold-induced recruitment of brown adipose tissue thermogenesis. Exp. Physiol. 2003, 88, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Bartness, T.J.; Bamshad, M. Innervation of mammalian white adipose tissue: Implications for the regulation of total body fat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 275, 1399–1411. [Google Scholar] [CrossRef]
- Wang, P.; Loh, K.H.; Wu, M.; Morgan, D.A.; Schneeberger, M.; Yu, X.; Chi, J.; Kosse, C.; Kim, D.; Rahmouni, K.; et al. A leptin–BDNF pathway regulating sympathetic innervation of adipose tissue. Nature 2020, 583, 839–844. [Google Scholar] [CrossRef]
- Yi, C.X.; Van Der Vliet, J.; Dai, J.; Yin, G.; Ru, L.; Buijs, R.M. Ventromedial arcuate nucleus communicates peripheral metabolic information to the suprachiasmatic nucleus. Endocrinology 2006, 147, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Chao, D.H.M.; León-Mercado, L.; Foppen, E.; Guzmán-Ruiz, M.; Basualdo, M.C.; Escobar, C.; Buijs, R.M. The suprachiasmatic nucleus modulates the sensitivity of arcuate nucleus to hypoglycemia in the male rat. Endocrinology 2016, 157, 3439–3451. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Ruiz, M.; Saderi, N.; Cazarez-Márquez, F.; Guerrero-Vargas, N.N.; Basualdo, M.C.; Acosta-Galván, G.; Buijs, R.M. The suprachiasmatic nucleus changes the daily activity of the arcuate nucleus α-MSH neurons in male rats. Endocrinology 2014, 155, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Buijs, F.N.; Guzmán-Ruiz, M.; León-Mercado, L.; Basualdo, M.C.; Escobar, C.; Kalsbeek, A.; Buijs, R.M. Suprachiasmatic nucleus interaction with the arcuate nucleus; Essential for organizing physiological rhythms. eNeuro 2017, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shieh, K.R.; Yang, S.C.; Lu, X.Y.; Akil, H.; Watson, S.J. Diurnal rhythmic expression of the rhythm-related genes, rPeriod1, rPeriod2, and rClock, in the rat brain. J. Biomed. Sci. 2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cedernaes, J.; Huang, W.; Ramsey, K.M.; Waldeck, N.; Marcheva, B.; Peek, C.B.; Levine, D.C.; Awatramani, R.; Bradfield, C.A.; Wang, X.A.; et al. Transcriptional basis for rhythmic control of hunger and metabolism within the AgRP neuron. Sleep Med. 2019. [Google Scholar] [CrossRef]
- Uyama, N.; Geerts, A.; Reynaert, H. Neural connections between the hypothalamus and the liver. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2004, 280, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Bell, B.B.; Harlan, S.M.; Morgan, D.A.; Guo, D.F.; Rahmouni, K. Differential contribution of POMC and AgRP neurons to the regulation of regional autonomic nerve activity by leptin. Mol. Metab. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Fliers, E.; Buijs, R.M. circadian disruption and scn control of energy. FEBS Lett. 2012, 585, 1412–1426. [Google Scholar] [CrossRef]
- Resch, J.M.; Maunze, B.; Phillips, K.A.; Choi, S.J. Inhibition of food intake by PACAP in the hypothalamic ventromedial nuclei is mediated by NMDA receptors. Physiol. Behav. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mounien, L.; Do Rego, J.C.; Bizet, P.; Boutelet, I.; Gourcerol, G.; Fournier, A.; Brabet, P.; Costentin, J.; Vaudry, H.; Jégou, S. Pituitary adenylate cyclase-activating polypeptide inhibits food intake in mice through activation of the hypothalamic melanocortin system. Neuropsychopharmacology 2009, 34, 424–435. [Google Scholar] [CrossRef] [Green Version]
- Morrison, C.D.; Xi, A.; White, C.L.; Ye, J.; Martin, R.J. Amino acids inhibit Agrp gene expression via an mTOR-dependent mechanism. Am. J. Physiol. Endocrinol. Metab. 2007, 293, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Kikuchi, O.; Shimpuku, M.; Susanti, V.Y.; Yokota-Hashimoto, H.; Taguchi, R.; Shibusawa, N.; Sato, T.; Tang, L.; Amano, K.; et al. Hypothalamic SIRT1 prevents age-associated weight gain by improving leptin sensitivity in mice. Diabetologia 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geerling, J.C.; Shin, J.W.; Chimenti, P.C.; Loewy, A.D. Paraventricular hypothalamic nucleus: Axonal projections to the brainstem. J. Comp. Neurol. 2010. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.V.; Latchford, K.J.; Sanon, W.K. The paraventricular nucleus of the hypothalamus—A potential target for integrative treatment of autonomic dysfunction. Expert Opin. Ther. Targets 2008, 12, 717–727. [Google Scholar] [CrossRef]
- Geerling, J.J.; Boon, M.R.; Kooijman, S.; Parlevliet, E.T.; Havekes, L.M.; Romijn, J.A.; Meurs, I.M.; Rensen, P.C.N. Sympathetic nervous system control of triglyceride metabolism: Novel concepts derived from recent studies. J. Lipid Res. 2014, 55, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.R.; Xu, Y.; Cassidy, R.M.; Lu, Y.; Yang, Y.; Tian, J.; Li, D.P.; Van Drunen, R.; Ribas-Latre, A.; Cai, Z.L.; et al. Paraventricular hypothalamus mediates diurnal rhythm of metabolism. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mazier, W.; Saucisse, N.; Simon, V.; Cannich, A.; Marsicano, G.; Massa, F.; Cota, D. mTORC1 and CB1 receptor signaling regulate excitatory glutamatergic inputs onto the hypothalamic paraventricular nucleus in response to energy availability. Mol. Metab. 2019, 28, 151–159. [Google Scholar] [CrossRef]
- King, B.M. The rise, fall, and resurrection of the ventromedial hypothalamus in the regulation of feeding behavior and body weight. Physiol. Behav. 2006, 87, 221–244. [Google Scholar] [CrossRef]
- Ishikawa, K.; Shimazu, T. Circadian rhythm of liver glycogen metabolism in rats: Effects of hypothalamic lesions. Am. J. Physiol. Endocrinol. Metab. 1980. [Google Scholar] [CrossRef]
- Shimizu, N.; Oomura, Y.; Plata-Salamán, C.R.; Morimoto, M. Hyperphagia and obesity in rats with bilateral ibotenic acid-induced lesions of the ventromedial hypothalamic nucleus. Brain Res. 1987, 416, 153–156. [Google Scholar] [CrossRef]
- Tong, Q.; Ye, C.P.; McCrimmon, R.J.; Dhillon, H.; Choi, B.; Kramer, M.D.; Yu, J.; Yang, Z.; Christiansen, L.M.; Lee, C.E.; et al. Synaptic Glutamate Release by Ventromedial Hypothalamic Neurons Is Part of the Neurocircuitry that Prevents Hypoglycemia. Cell Metab. 2007, 5, 383–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orozco-Solis, R.; Aguilar-Arnal, L.; Murakami, M.; Peruquetti, R.; Ramadori, G.; Coppari, R.; Sassone-Corsi, P. The circadian clock in the ventromedial hypothalamus controls cyclic energy expenditure. Cell Metab. 2016, 23, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Khodai, T.; Nunn, N.; Worth, A.A.; Feetham, C.H.; Belle, M.D.C.; Piggins, H.D.; Luckman, S.M. PACAP neurons in the ventromedial hypothalamic nucleus are glucose inhibited and their selective activation induces hyperglycaemia. Front. Endocrinol. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Hawke, Z.; Ivanov, T.R.; Bechtold, D.A.; Dhillon, H.; Lowell, B.B.; Luckman, S.M. PACAP Neurons in the Hypothalamic Ventromedial Nucleus Are Targets of Central Leptin Signaling. J. Neurosci. 2009, 29, 14828–14835. [Google Scholar] [CrossRef] [Green Version]
- Bookout, A.L.; De Groot, M.H.M.; Owen, B.M.; Lee, S.; Gautron, L.; Lawrence, H.L.; Ding, X.; Elmquist, J.K.; Takahashi, J.S.; Mangelsdorf, D.J.; et al. FGF21 regulates metabolism and circadian behavior by acting on the nervous system. Nat. Med. 2013. [Google Scholar] [CrossRef]
- Jensen-Cody, S.O.; Flippo, K.H.; Claflin, K.E.; Yavuz, Y.; Sapouckey, S.A.; Walters, G.C.; Usachev, Y.M.; Atasoy, D.; Gillum, M.P.; Potthoff, M.J. FGF21 Signals to Glutamatergic Neurons in the Ventromedial Hypothalamus to Suppress Carbohydrate Intake. Cell Metab. 2020. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Sánchez, N.; Seoane-Collazo, P.; Contreras, C.; Varela, L.; Villarroya, J.; Rial-Pensado, E.; Buqué, X.; Aurrekoetxea, I.; Delgado, T.C.; Vázquez-Martínez, R.; et al. Hypothalamic AMPK-ER Stress-JNK1 Axis Mediates the Central Actions of Thyroid Hormones on Energy Balance. Cell Metab. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramadori, G.; Fujikawa, T.; Anderson, J.; Berglund, E.D.; Frazao, R.; Michán, S.; Vianna, C.R.; Sinclair, D.A.; Elias, C.F.; Coppari, R. SIRT1 deacetylase in SF1 neurons protects against metabolic imbalance. Cell Metab. 2011, 14, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.H.; Canteras, N.S.; Swanson, L.W. Organization of projections from the dorsomedial nucleus of the hypothalamus: A PHA-L study in the rat. J. Comp. Neurol. 1996. [Google Scholar] [CrossRef]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.S.; Xu, M.; Zhang, Z.; Chang, W.C.; Gaj, T.; Schaffer, D.V.; Dan, Y. A Hypothalamic Switch for REM and Non-REM Sleep. Neuron 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriguchi, T.; Sakurai, T.; Nambu, T.; Yanagisawa, M.; Goto, K. Neurons containing orexin in the lateral hypothalamic area of the adult rat brain are activated by insulin-induced acute hypoglycemia. Neurosci. Lett. 1999. [Google Scholar] [CrossRef]
- Gold, R.M. Hypothalamic obesity: The myth of the ventromedial nucleus. Science 1973. [Google Scholar] [CrossRef] [PubMed]
- Bernardis, L.L.; Bellinger, L.L. The lateral hypothalamic area revisited: Neuroanatomy, body weight regulation, neuroendocrinology and metabolism. Neurosci. Biobehav. Rev. 1993. [Google Scholar] [CrossRef]
- Oomura, Y.; Ooyama, H.; Sugimori, M.; Nakamura, T.; Yamada, Y. Glucose inhibition of the glucose-sensitive neurone in the rat lateral hypothalamus. Nature 1974. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.R.; Patterson, L.M.; Sutton, G.M.; Morrison, C.; Zheng, H. Orexin inputs to caudal raphé neurons involved in thermal, cardiovascular, and gastrointestinal regulation. Histochem. Cell Biol. 2005. [Google Scholar] [CrossRef]
- Martins, L.; Seoane-Collazo, P.; Contreras, C.; González-García, I.; Martínez-Sánchez, N.; González, F.; Zalvide, J.; Gallego, R.; Diéguez, C.; Nogueiras, R.; et al. A Functional Link between AMPK and Orexin Mediates the Effect of BMP8B on Energy Balance. Cell Rep. 2016, 16, 2231–2242. [Google Scholar] [CrossRef] [Green Version]
- López-Ferreras, L.; Richard, J.E.; Noble, E.E.; Eerola, K.; Anderberg, R.H.; Olandersson, K.; Taing, L.; Kanoski, S.E.; Hayes, M.R.; Skibicka, K.P. Lateral hypothalamic GLP-1 receptors are critical for the control of food reinforcement, ingestive behavior and body weight. Mol. Psychiatry 2018, 23, 1157–1168. [Google Scholar] [CrossRef] [Green Version]
- Everson, C.A.; Bergmann, B.M.; Rechtschaffen, A. Sleep deprivation in the rat: III. Total sleep deprivation. Sleep 1989, 12, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Lin, L.; Austin, D.; Young, T.; Mignot, E. Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index. PLoS Med. 2004. [Google Scholar] [CrossRef]
- ANDERSSON., B. The Effect of Injections of Hypertonic NaCl-Solutions into Different Parts of the Hypothalamus of Goats. Acta Physiol. Scand. 1953. [Google Scholar] [CrossRef]
- ANDERSSON, B.; McCANN, S.M. Drinking, Antidiuresis and Milk Ejection from Electrical Stimulation within the Hypothalamus of the Goat. Acta Physiol. Scand. 1955. [Google Scholar] [CrossRef]
- Llewellyn, T.; Zheng, H.; Liu, X.; Xu, B.; Patel, K.P. Median preoptic nucleus and subfornical organ drive renal sympathetic nerve activity via a glutamatergic mechanism within the paraventricular nucleus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabor, A.; Leenen, F.H.H. Central neuromodulatory pathways regulating sympathetic activity in hypertension. J. Appl. Physiol. 2012, 113, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Gizowski, C.; Zaelzer, C.; Bourque, C.W. Clock-driven vasopressin neurotransmission mediates anticipatory thirst prior to sleep. Nature 2016, 537, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Trudel, E.; Bourque, C.W. Central clock excites vasopressin neurons by waking osmosensory afferents during late sleep. Nat. Neurosci. 2010. [Google Scholar] [CrossRef]
- Zimmerman, C.A.; Leib, D.E.; Knight, Z.A. Neural circuits underlying thirst and fluid homeostasis. Nat. Rev. Neurosci. 2017, 18, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Northeast, R.C.; Chrobok, L.; Hughes, A.T.L.; Petit, C.; Piggins, H.D. Keeping time in the lamina terminalis: Novel oscillator properties of forebrain sensory circumventricular organs. FASEB J. 2020, 34, 974–987. [Google Scholar] [CrossRef] [Green Version]
- Chaput, J.P.; Després, J.P.; Bouchard, C.; Tremblay, A. Association of sleep duration with type 2 diabetes and impaired glucose tolerance. Diabetologia 2007. [Google Scholar] [CrossRef] [Green Version]
- Grandner, M.A.; Schopfer, E.A.; Sands-Lincoln, M.; Jackson, N.; Malhotra, A. Relationship between sleep duration and body mass index depends on age. Obesity 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, J.A.; Eberhardt, N.L.; Jensen, M.D. Role of nonexercise activity thermogenesis in resistance to fat gain in humans. Science 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirelli, C.; Gutierrez, C.M.; Tononi, G. Extensive and Divergent Effects of Sleep and Wakefulness on Brain Gene Expression. Neuron 2004. [Google Scholar] [CrossRef] [Green Version]
- Kroeger, D.; Absi, G.; Gagliardi, C.; Bandaru, S.S.; Madara, J.C.; Ferrari, L.L.; Arrigoni, E.; Münzberg, H.; Scammell, T.E.; Saper, C.B.; et al. Galanin neurons in the ventrolateral preoptic area promote sleep and heat loss in mice. Nat. Commun. 2018, 9, 4129. [Google Scholar] [CrossRef] [Green Version]
- Tasali, E.; Leproult, R.; Ehrmann, D.A.; Van Cauter, E. Slow-wave sleep and the risk of type 2 diabetes in humans. Proc. Natl. Acad. Sci. USA 2008. [Google Scholar] [CrossRef] [Green Version]
- Isobe, Y.; Nishino, H. Signal transmission from the suprachiasmatic nucleus to the pineal gland via the paraventricular nucleus: Analysed from arg-vasopressin peptide, rPer2 mRNA and AVP mRNA changes and pineal AA-NAT mRNA after the melatonin injection during light and dark per. Brain Res. 2004. [Google Scholar] [CrossRef]
- Saper, C.B.; Cano, G.; Scammell, T.E. Homeostatic, circadian, and emotional regulation of sleep. J. Comp. Neurol. 2005, 493, 92–98. [Google Scholar] [CrossRef]
- Adamantidis, A.; de Lecea, L. Sleep and metabolism: Shared circuits, new connections. Trends Endocrinol. Metab. 2008, 19, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Funato, H.; Tsai, A.L.; Willie, J.T.; Kisanuki, Y.; Williams, S.C.; Sakurai, T.; Yanagisawa, M. Enhanced Orexin Receptor-2 Signaling Prevents Diet-Induced Obesity and Improves Leptin Sensitivity. Cell Metab. 2009. [Google Scholar] [CrossRef] [Green Version]
- Gonzàlez, J.A.; Reimann, F.; Burdakov, D. Dissociation between sensing and metabolism of glucose in sugar sensing neurones. J. Physiol. 2009, 587, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Borniger, J.C.; Walker, W.H.; sURBHI; Emmer, K.M.; Zhang, N.; Zalenski, A.A.; Muscarella, S.L.; Fitzgerald, J.A.; Smith, A.N.; Braam, C.J.; et al. A Role for Hypocretin/Orexin in Metabolic and Sleep Abnormalities in a Mouse Model of Non-metastatic Breast Cancer. Cell Metab. 2018, 28, 118–129.e5. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.A.; Kumar, S.; McGinty, D.; Alam, M.N.; Szymusiak, R. Neuronal activity in the preoptic hypothalamus during sleep deprivation and recovery sleep. J. Neurophysiol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Varin, C.; Rancillac, A.; Geoffroy, H.; Arthaud, S.; Fort, P.; Gallopin, T. Glucose induces slow-wave sleep by exciting the sleep- promoting neurons in the ventrolateral preoptic nucleus: A new link between sleep and metabolism. J. Neurosci. 2015, 35, 9900–9911. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C.; Bjorkum, A.A.; Gaus, S.E.; Lu, J.; Scammell, T.E.; Saper, C.B. Afferents to the ventrolateral preoptic nucleus. J. Neurosci. 2002. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, H.; Nakamaru-Ogiso, E.; Hamada, K.; Hensch, T.K. Serotonergic integration of circadian clock and ultradian sleep-wake cycles. J. Neurosci. 2012, 32, 14794–14803. [Google Scholar] [CrossRef] [Green Version]
- Saper, C.B.; Fuller, P.M.; Pedersen, N.P.; Lu, J.; Scammell, T.E. Sleep State Switching. Neuron 2010, 68, 1023–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minokoshi, Y.; Kim, Y.B.; Peroni, O.D.; Fryer, L.G.D.; Müller, C.; Carling, D.; Kahn, B.B. Leptin stimulates fatty-acid oxidation by activating AMP-activated protein kinase. Nature 2002, 415, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, L.; Gamber, K.; Greeley, S.; Silva, J.; Huntoon, N.; Leng, X.H.; Bjørbæk, C. Leptin-Dependent Control of Glucose Balance and Locomotor Activity by POMC Neurons. Cell Metab. 2009, 9, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Kettner, N.M.; Mayo, S.A.; Hua, J.; Lee, C.; Moore, D.D.; Fu, L. Circadian dysfunction induces leptin resistance in mice. Cell Metab. 2015, 22, 448–459. [Google Scholar] [CrossRef] [Green Version]
- Paschos, G.K.; Ibrahim, S.; Song, W.L.; Kunieda, T.; Grant, G.; Reyes, T.M.; Bradfield, C.A.; Vaughan, C.H.; Eiden, M.; Masoodi, M.; et al. Obesity in mice with adipocyte-specific deletion of clock component Arntl. Nat. Med. 2012. [Google Scholar] [CrossRef] [Green Version]
- Jimeno, B.; Hau, M.; Verhulst, S. Corticosterone levels reflect variation in metabolic rate, independent of ‘stress’. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Leon-Mercado, L.; Chao, D.H.M.; del Carmen Basualdo, M.; Kawata, M.; Escobar, C.; Buijs, R.M. The arcuate nucleus: A site of fast negative feedback for corticosterone secretion in male rats. eNeuro 2017, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Cavagnini, F.; Croci, M.; Putignano, P.; Petroni, M.L.; Invitti, C. Glucocorticoids and neuroendocrine function. Int. J. Obes. 2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuperman, Y.; Weiss, M.; Dine, J.; Staikin, K.; Golani, O.; Ramot, A.; Nahum, T.; Kühne, C.; Shemesh, Y.; Wurst, W.; et al. CRFR1 in AgRP Neurons Modulates Sympathetic Nervous System Activity to Adapt to Cold Stress and Fasting. Cell Metab. 2016, 23, 1185–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sefton, C.; Harno, E.; Davies, A.; Small, H.; Allen, T.J.; Wray, J.R.; Lawrence, C.B.; Coll, A.P.; White, A. Elevated hypothalamic glucocorticoid levels are associated with obesity and hyperphagia in male mice. Endocrinology 2016, 157, 4257–4265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kas, M.J.H.; Bruijnzeel, A.W.; Haanstra, J.R.; Wiegant, V.M.; Adan, R.A.H. Differential regulation of agouti-related protein and neuropeptide Y in hypothalamic neurons following a stressful event. J. Mol. Endocrinol. 2005. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, E.L.; DeJoseph, M.R.; Brownfield, M.S.; Urban, J.H. Involvement of neuropeptide Y Y1 receptors in the regulation of neuroendocrine corticotropin-releasing hormone neuronal activity. Endocrinology 2007, 148, 3666–3673. [Google Scholar] [CrossRef] [Green Version]
- Stanley, B.G.; Leibowitz, S.F. Neuropeptide Y injected in the paraventricular hypothalamus: A powerful stimulant of feeding behavior. Proc. Natl. Acad. Sci. USA 1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erion, R.; King, A.N.; Wu, G.; Hogenesch, J.B.; Sehgal, A. Neural clocks and neuropeptide F/Y regulate circadian gene expression in a peripheral metabolic tissue. Elife 2016, 5, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Borjigin, J.; Samantha Zhang, L.; Calinescu, A.A. Circadian regulation of pineal gland rhythmicity. Mol. Cell. Endocrinol. 2012, 349, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Forrestel, A.C.; Miedlich, S.U.; Yurcheshen, M.; Wittlin, S.D.; Sellix, M.T. Chronomedicine and type 2 diabetes: Shining some light on melatonin. Diabetologia 2017, 60, 808–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Fleur, S.E.; Kalsbeek, A.; Wortel, J.; Van Der Vliet, J.; Buijs, R.M. Role for the pineal and melatonin in glucose homeostasis: Pinealectomy increases night- time glucose concentrations. J. Neuroendocrinol. 2001. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; La Fleur, S.; Van Heijningen, C.; Buijs, R.M. Suprachiasmatic GABAergic inputs to the paraventricular nucleus control plasma glucose concentrations in the rat via sympathetic innervation of the liver. J. Neurosci. 2004, 24, 7604–7613. [Google Scholar] [CrossRef] [Green Version]
- Gamble, K.L.; Allen, G.C.; Zhou, T.; McMahon, D.G. Gastrin-releasing peptide mediates light-like resetting of the suprachiasmatic nucleus circadian pacemaker through cAMP response element-binding protein and Per1 activation. J. Neurosci. 2007, 27, 12078–12087. [Google Scholar] [CrossRef]
- Alvarez, Y.; Glotfelty, L.G.; Blank, N.; Dohnalová, L.; Thaiss, C.A. The Microbiome as a Circadian Coordinator of Metabolism. Endocrinology 2020, 161, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016. [Google Scholar] [CrossRef] [Green Version]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom control of microbiota diurnal oscillations promotes metabolic homeostasis. Cell 2014. [Google Scholar] [CrossRef] [Green Version]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of diurnal variation of gut microbes and high-fat feeding on host circadian clock function and metabolism. Cell Host Microbe 2015. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Bushman, F.D.; FitzGerald, G.A. Rhythmicity of the intestinal microbiota is regulated by gender and the host circadian clock. Proc. Natl. Acad. Sci. USA 2015. [Google Scholar] [CrossRef] [Green Version]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and metabolic syndrome in circadian Clock mutant nice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Liu, A.; Weidenhammer, A.; Cooksey, R.C.; McClain, D.; Kim, M.K.; Aguilera, G.; Abel, E.D.; Chung, J.H. The role of mPer2 clock gene in glucocorticoid and feeding rhythms. Endocrinology 2009. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.R.; Hu, F.B. Short sleep duration and weight gain: A systematic review. Obesity 2008, 16, 643–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bo, S.; Ciccone, G.; Durazzo, M.; Ghinamo, L.; Villois, P.; Canil, S.; Gambino, R.; Cassader, M.; Gentile, L.; Cavallo-Perin, P. Contributors to the obesity and hyperglycemia epidemics. A prospective study in a population-based cohort. Int. J. Obes. 2011. [Google Scholar] [CrossRef] [Green Version]
- Shan, Z.; Li, Y.; Zong, G.; Guo, Y.; Li, J.; Manson, J.E.; Hu, F.B.; Willett, W.C.; Schernhammer, E.S.; Bhupathiraju, S.N. Rotating night shift work and adherence to unhealthy lifestyle in predicting risk of type 2 diabetes: Results from two large US cohorts of female nurses. BMJ 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Åkerstedt, T. Shift work and disturbed sleep/wakefulness. Occup. Med. 2003, 53, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crispim, C.A.; Waterhouse, J.; Dâmaso, A.R.; Zimberg, I.Z.; Padilha, H.G.; Oyama, L.M.; Tufik, S.; De Mello, M.T. Hormonal appetite control is altered by shift work: A preliminary study. Metabolism 2011. [Google Scholar] [CrossRef] [Green Version]
- Scheer, F.A.J.L.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009. [Google Scholar] [CrossRef] [Green Version]
- Sack, R.L.; Auckley, D.; Auger, R.R.; Carskadon, M.A.; Wright, K.P.; Vitiello, M.V.; Zhdanova, I.V. Circadian rhythm sleep disorders: Part I, basic principles, shift work and jet lag disorders. An American Academy of Sleep Medicine review. Sleep 2007, 30, 1460–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markwald, R.R.; Melanson, E.L.; Smith, M.R.; Higgins, J.; Perreault, L.; Eckel, R.H.; Wright, K.P. Impact of insufficient sleep on total daily energy expenditure, food intake, and weight gain. Proc. Natl. Acad. Sci. USA 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; Ditacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.J.; et al. Article Time-Restricted Feeding without Reducing Caloric Intake Prevents Metabolic Diseases in Mice Fed a High-Fat Diet. Cell Metab. 2012. [Google Scholar] [CrossRef] [Green Version]
- Kurose, T.; Ueta, Y.; Yamamoto, Y.; Serino, R.; Ozaki, Y.; Saito, J.; Nagata, S.; Yamashita, H. Effects of restricted feeding on the activity of hypothalamic Orexin (OX)-A containing neurons and OX2 receptor mRNA level in the paraventricular nucleus of rats. Regul. Pept. 2002, 104, 145–151. [Google Scholar] [CrossRef]
- Sundaram, S.; Yan, L. Time-restricted feeding reduces adiposity in mice fed a high-fat diet. Nutr. Res. 2016, 36, 603–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasumoto, Y.; Hashimoto, C.; Nakao, R.; Yamazaki, H.; Hiroyama, H.; Nemoto, T.; Yamamoto, S.; Sakurai, M.; Oike, H.; Wada, N.; et al. Short-term feeding at the wrong time is sufficient to desynchronize peripheral clocks and induce obesity with hyperphagia, physical inactivity and metabolic disorders in mice. Metabolism 2016. [Google Scholar] [CrossRef]
- Christie, S.; Vincent, A.D.; Li, H.; Frisby, C.L.; Kentish, S.J.; O’rielly, R.; Wittert, G.A.; Page, A.J. A rotating light cycle promotes weight gain and hepatic lipid storage in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, H.; Hanzawa, F.; Kim, D.; Sun, S.; Laurent, T.; Umeki, M.; Ikeda, S.; Mochizuki, S.; Oda, H. Delayed first active-phase meal, a breakfastskipping model, led to increased body weight and shifted the circadian oscillation of the hepatic clock and lipid metabolism-related genes in rats fed a high-fat diet. PLoS ONE 2018. [Google Scholar] [CrossRef]
- Chaix, A.; Zarrinpar, A.; Miu, P.; Panda, S. Time-restricted feeding is a preventative and therapeutic intervention against diverse nutritional challenges. Cell Metab. 2014. [Google Scholar] [CrossRef] [Green Version]
- Chaix, A.; Lin, T.; Le, H.D.; Chang, M.W.; Panda, S. Time-Restricted Feeding Prevents Obesity and Metabolic Syndrome in Mice Lacking a Circadian Clock. Cell Metab. 2019, 29, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Olsen, M.K.; Choi, M.H.; Kulseng, B.; Zhao, C.M.; Chen, D. Time-restricted feeding on weekdays restricts weight gain: A study using rat models of high-fat diet-induced obesity. Physiol. Behav. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.T.; Chiang, W.Y.; Hsieh, W.H.; Escobar, C.; Buijs, R.M.; Hu, K. Interactive effects of dorsomedial hypothalamic nucleus and time-restricted feeding on fractal motor activity regulation. Front. Physiol. 2016. [Google Scholar] [CrossRef] [Green Version]
- Boucsein, A.; Rizwan, M.Z.; Tups, A. Hypothalamic leptin sensitivity and health benefits of time-restricted feeding are dependent on the time of day in male mice. FASEB J. 2019, 33, 12175–12187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrell, J.; Yates, E.; Rivir, M.; Woods, S.C.; Hogenesch, J.B.; Perez-Tilve, D. The central melanocortin system mediates the benefits of time-restricted feeding on energy balance. Physiol. Behav. 2020, 227, 113132. [Google Scholar] [CrossRef] [PubMed]
- Dyar, K.A.; Lutter, D.; Artati, A.; Ceglia, N.J.; Liu, Y.; Armenta, D.; Jastroch, M.; Schneider, S.; de Mateo, S.; Cervantes, M.; et al. Atlas of Circadian Metabolism Reveals System-wide Coordination and Communication between Clocks. Cell 2018, 174, 1571–1585.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckel-Mahan, K.L.; Patel, V.R.; De Mateo, S.; Orozco-Solis, R.; Ceglia, N.J.; Sahar, S.; Dilag-Penilla, S.A.; Dyar, K.A.; Baldi, P.; Sassone-Corsi, P. Reprogramming of the circadian clock by nutritional challenge. Cell 2013, 155, 1464–1478. [Google Scholar] [CrossRef] [Green Version]
- Clemenzi, M.N.; Martchenko, A.; Loganathan, N.; Tse, E.K.; Brubaker, P.L.; Belsham, D.D. Analysis of Western diet, palmitate and BMAL1 regulation of neuropeptide Y expression in the murine hypothalamus and BMAL1 knockout cell models. Mol. Cell. Endocrinol. 2020, 507, 110773. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.; He, W.; Jiang, N.; Chen, J.T.C.; Belsham, D.D. NAMPT and BMAL1 Are Independently Involved in the Palmitate-Mediated Induction of Neuroinflammation in Hypothalamic Neurons. Front. Endocrinol. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Zhu, C.; Xu, Y.; Jiang, Z.; Tian, J.B.; Cassidy, R.M.; Cai, Z.; Shu, G.; Xu, Y.; Xue, M.; Arenkiel, B.R.; et al. Disrupted hypothalamic CRH neuron responsiveness contributes to diet-induced obesity. EMBO Rep. 2020, 1–14. [Google Scholar] [CrossRef]
- Mazzone, C.M.; Liang-Guallpa, J.; Li, C.; Wolcott, N.S.; Boone, M.H.; Southern, M.; Kobzar, N.P.; de Araujo Salgado, I.; Reddy, D.M.; Sun, F.; et al. High-fat food biases hypothalamic and mesolimbic expression of consummatory drives. Nat. Neurosci. 2020, 23, 1253–1266. [Google Scholar] [CrossRef]
- Morrison, S.F.; Madden, C.J.; Tupone, D. Central neural regulation of brown adipose tissue thermogenesis and energy expenditure. Cell Metab. 2014, 19, 741–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, S.; Sato, T.; Tateyama, M.; Kageyama, H.; Maejima, Y.; Nakata, M.; Hirako, S.; Matsuo, T.; Kyaw, S.; Shiuchi, T.; et al. Activation of AMPK-Regulated CRH Neurons in the PVH is Sufficient and Necessary to Induce Dietary Preference for Carbohydrate over Fat. Cell Rep. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, J.; Morris, C.J.; Caputo, R.; Wang, W.; Garaulet, M.; Scheer, F.A.J.L. Sex differences in the circadian misalignment effects on energy regulation. Proc. Natl. Acad. Sci. USA 2019, 116, 23806–23812. [Google Scholar] [CrossRef]
- Cespedes Feliciano, E.M.; Rifas-Shiman, S.L.; Quante, M.; Redline, S.; Oken, E.; Taveras, E.M. Chronotype, Social Jet Lag, and Cardiometabolic Risk Factors in Early Adolescence. JAMA Pediatr. 2019. [Google Scholar] [CrossRef]
- Leng, Y.; Blackwell, T.; Cawthon, P.M.; Ancoli-Israel, S.; Stone, K.L.; Yaffe, K. Association of Circadian Abnormalities in Older Adults with an Increased Risk of Developing Parkinson Disease. JAMA Neurol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Nicolas, A.; Madrid, J.A.; García, F.J.; Campos, M.; Moreno-Casbas, M.T.; Almaida-Pagán, P.F.; Lucas-Sánchez, A.; Rol, M.A. Circadian monitoring as an aging predictor. Sci. Rep. 2018. [Google Scholar] [CrossRef]
- Bellanti, F.; Iannelli, G.; Blonda, M.; Tamborra, R.; Villani, R.; Romano, A.; Calcagnini, S.; Mazzoccoli, G.; Vinciguerra, M.; Gaetani, S.; et al. Alterations of Clock Gene RNA Expression in Brain Regions of a Triple Transgenic Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 59, 615–631. [Google Scholar] [CrossRef] [Green Version]
- Nordengen, K.; Kirsebom, B.E.; Henjum, K.; Selnes, P.; Gísladóttir, B.; Wettergreen, M.; Torsetnes, S.B.; Grøntvedt, G.R.; Waterloo, K.K.; Aarsland, D.; et al. Glial activation and inflammation along the Alzheimer’s disease continuum. J. Neuroinflamm. 2019, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Garwood, C.J.; Pooler, A.M.; Atherton, J.; Hanger, D.P.; Noble, W. Astrocytes are important mediators of Aβ-induced neurotoxicity and tau phosphorylation in primary culture. Cell Death Dis. 2011. [Google Scholar] [CrossRef] [Green Version]
- Fakhoury, M. Microglia and astrocytes in Alzheimer’s disease: Implications for therapy. Curr. Neuropharmacol. 2017. [Google Scholar] [CrossRef]
- Hayashi, Y.; Koyanagi, S.; Kusunose, N.; Okada, R.; Wu, Z.; Tozaki-Saitoh, H.; Ukai, K.; Kohsaka, S.; Inoue, K.; Ohdo, S.; et al. The intrinsic microglial molecular clock controls synaptic strength via the circadian expression of cathepsin S. Sci. Rep. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojarskaite, L.; Bjørnstad, D.M.; Pettersen, K.H.; Cunen, C.; Hermansen, G.H.; Åbjørsbråten, K.S.; Chambers, A.R.; Sprengel, R.; Vervaeke, K.; Tang, W.; et al. Astrocytic Ca2+ signaling is reduced during sleep and is involved in the regulation of slow wave sleep. Nat. Commun. 2020. [Google Scholar] [CrossRef]
- Hayashi, Y. Diurnal Spatial Rearrangement of Microglial Processes through the Rhythmic Expression of P2Y12 Receptors. J. Neurol. Disord. 2013. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Wu, Z.; Meng, J.; Saito, T.; Saido, T.C.; Qing, H.; Nakanishi, H. An impaired intrinsic microglial clock system induces neuroinflammatory alterations in the early stage of amyloid precursor protein knock-in mouse brain. J. Neuroinflamm. 2019, 16, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonken, L.K.; Kitt, M.M.; Gaudet, A.D.; Barrientos, R.M.; Watkins, L.R.; Maier, S.F. Diminished circadian rhythms in hippocampal microglia may contribute to age-related neuroinflammatory sensitization. Neurobiol. Aging 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Ma, N.; Yu, B.; Zhang, W.; Wan, J. Transcriptomic profiling of microglia and astrocytes throughout aging. J. Neuroinflamm. 2020, 17, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, D.G.; Stopa, E.G.; Kuo-Leblanc, V.; McKee, A.C.; Asayama, K.; Volicer, L.; Kowall, N.; Satlin, A. Dorsomedial SCN neuronal subpopulations subserve different functions in human dementia. Brain 2008, 131, 1609–1617. [Google Scholar] [CrossRef] [Green Version]
- Baloyannis, S.J.; Mavroudis, I.; Mitilineos, D.; Baloyannis, I.S.; Costa, V.G. The Hypothalamus in Alzheimer’s Disease: A Golgi and Electron Microscope Study. Am. J. Alzheimer’s Dis. Other Demen. 2015, 30, 478–487. [Google Scholar] [CrossRef]
- Swaab, D.F.; Fliers, E.; Partiman, T.S. The suprachiasmatic nucleus of the human brain in relation to sex, age and senile dementia. Brain Res. 1985. [Google Scholar] [CrossRef]
- McDuff, T.; Sumi, S.M. Subcortical degeneration in Alzheimer’s disease. Neurology 1985, 35, 123. [Google Scholar] [CrossRef]
- Storandt, M.; Kaskie, B.; Von Dras, D.D. Temporal memory for remote events in healthy aging and dementia. Psychol. Aging 1998. [Google Scholar] [CrossRef]
- Carroll, J.C.; Iba, M.; Bangasser, D.A.; Valentino, R.J.; James, M.J.; Brunden, K.R.; Lee, V.M.Y.; Trojanowski, J.Q. Chronic stress exacerbates tau pathology, neurodegeneration, and cognitive performance through a corticotropin-releasing factor receptor-dependent mechanism in a transgenic mouse model of tauopathy. J. Neurosci. 2011. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Moon, M.; Choe, H.K.; Han, D.H.; Jang, C.; Kim, A.; Cho, S.; Kim, K.; Mook-Jung, I. Aβ-induced degradation of BMAL1 and CBP leads to circadian rhythm disruption in Alzheimer’s disease. Mol. Neurodegener. 2015, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kress, G.J.; Liao, F.; Dimitry, J.; Cedeno, M.R.; FitzGerald, G.A.; Holtzman, D.M.; Musiek, E.S. Regulation of amyloid-β dynamics and pathology by the circadian clock. J. Exp. Med. 2018, 215, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.E.; Lim, M.M.; Bateman, R.J.; Lee, J.J.; Smyth, L.P.; Cirrito, J.R.; Fujiki, N.; Nishino, S.; Holtzman, D.M. Amyloid-β dynamics are regulated by orexin and the sleep-wake cycle. Science 2009. [Google Scholar] [CrossRef] [Green Version]
- Musiek, E.S.; Bhimasani, M.; Zangrilli, M.A.; Morris, J.C.; Holtzman, D.M.; Ju, Y.E.S. Circadian rest-activity pattern changes in aging and preclinical Alzheimer disease. JAMA Neurol. 2018. [Google Scholar] [CrossRef]
- Cronin, P.; McCarthy, M.J.; Lim, A.S.P.; Salmon, D.P.; Galasko, D.; Masliah, E.; De Jager, P.L.; Bennett, D.A.; Desplats, P. Circadian alterations during early stages of Alzheimer’s disease are associated with aberrant cycles of DNA methylation in BMAL1. Alzheimer’s Dement. 2017. [Google Scholar] [CrossRef]
- Khachiyants, N.; Trinkle, D.; Son, S.J.; Kim, K.Y. Sundown syndrome in persons with dementia: An update. Psychiatry Investig. 2011. [Google Scholar] [CrossRef]
- Canevelli, M.; Valletta, M.; Trebbastoni, A.; Sarli, G.; D’Antonio, F.; Tariciotti, L.; de Lena, C.; Bruno, G. Sundowning in Dementia: Clinical relevance, pathophysiological determinants, and therapeutic approaches. Front. Med. 2016, 3, 73. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Mansfield, J.; Garfinkel, D.; Lipson, S. Melatonin for treatment of sundowning in elderly persons with dementia—A preliminary study. Arch. Gerontol. Geriatr. 2000, 31, 65–76. [Google Scholar] [CrossRef]
- Klaffke, S.; Staedt, J. Sundowing and circadian rhythm disorders in dementia. Acta Neurol. Belg. 2006, 106, 168. [Google Scholar]
- Yao, Y.; Zhang, W.; Ming, R.; Deng, Q.; Zuo, A.; Zhang, S.; Ying, Y.; Zhao, Y.; Ma, J. Noninvasive 40-Hz Light Flicker Rescues Circadian Behavior and Abnormal Lipid Metabolism Induced by Acute Ethanol Exposure via Improving SIRT1 and the Circadian Clock in the Liver-Brain Axis. Front. Pharmacol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, D.A.; Rodríguez, M.T.C.; Rodríguez, P.H.; Sánchez, M.F.; Mora, J.A.V.; Gago-Veiga, A.B. Is There a Characteristic Clinical Profile for Patients with Dementia and Sundown Syndrome? J. Alzheimer’s Dis. 2018. [Google Scholar] [CrossRef]
- Todd, W.D.; Fenselau, H.; Wang, J.L.; Zhang, R.; Machado, N.L.; Venner, A.; Broadhurst, R.Y.; Kaur, S.; Lynagh, T.; Olson, D.P.; et al. A hypothalamic circuit for the circadian control of aggression. Nat. Neurosci. 2018, 21, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Todd, W.D. Potential Pathways for Circadian Dysfunction and Sundowning-Related Behavioral Aggression in Alzheimer’s Disease and Related Dementias. Front. Neurosci. 2020, 14, 1–14. [Google Scholar] [CrossRef]
- Kunwar, P.S.; Zelikowsky, M.; Remedios, R.; Cai, H.; Yilmaz, M.; Meister, M.; Anderson, D.J. Ventromedial hypothalamic neurons control a defensive emotion state. Elife 2015. [Google Scholar] [CrossRef]
- Rosenthal, N.E.; Sack, D.A.; Gillin, J.C.; Lewy, A.J.; Goodwin, F.K.; Davenport, Y.; Mueller, P.S.; Newsome, D.A.; Wehr, T.A. Seasonal Affective Disorder: A Description of the Syndrome and Preliminary Findings With Light Therapy. Arch. Gen. Psychiatry 1984. [Google Scholar] [CrossRef]
- Lewy, A.J.; Lefler, B.J.; Emens, J.S.; Bauer, V.K. The circadian basis of winter depression. Proc. Natl. Acad. Sci. USA 2006, 103, 7414–7419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terman, M.; Terman, J.S.; Quitkin, F.M.; McGrath, P.J.; Stewart, J.W.; Rafferty, B. Light therapy for Seasonal Affective Disorder. A review of efficacy. Neuropsychopharmacology 1989, 2, 1–22. [Google Scholar] [CrossRef]
- Zhou, J.N.; Hofman, M.A.; Swaab, D.F. VIP neurons in the human SCN in relation to sex, age, and Alzheimer’s disease. Neurobiol. Aging 1995, 16, 571–576. [Google Scholar] [CrossRef]
- Roy, U.; Heredia-Muñoz, M.T.; Stute, L.; Höfling, C.; Matysik, J.; Meijer, J.H.; Roßner, S.; Alia, A. Degeneration of the Suprachiasmatic Nucleus in an Alzheimer’s Disease Mouse Model Monitored by in vivo Magnetic Resonance Relaxation Measurements and Immunohistochemistry. J. Alzheimer’s Dis. 2019, 69, 363–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagare, R.; Possidente, B.; Lagalwar, S.; Figueiro, M.G. Robust light–dark patterns and reduced amyloid load in an Alzheimer’s disease transgenic mouse model. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Stief, A. über die anatomischen Grundlagen der vegetativen Störungen bei Geisteskrankheiten. Dtsch. Z. Nervenheilkd. 1927. [Google Scholar] [CrossRef]
- Csernansky, J.G.; Dong, H.; Fagan, A.M.; Wang, L.; Xiong, C.; Holtzman, D.M.; Morris, J.C. Plasma cortisol and progression of dementia in subjects with Alzheimer-type dementia. Am. J. Psychiatry 2006, 163, 2164–2169. [Google Scholar] [CrossRef] [PubMed]
- Souza, E.B.D.; Whitehouse, P.J.; Price, D.L.; Vale, W.W. Abnormalities in Corticotropin-releasing Hormone (CRH) in Alzheimer’s Disease and Other Human Disorders. Ann. N. Y. Acad. Sci. 1987, 512, 237–247. [Google Scholar] [CrossRef]
- Gallone, S.; Boschi, S.; Rubino, E.; De Martino, P.; Scarpini, E.; Galimberti, D.; Fenoglio, C.; Acutis, P.L.; Maniaci, M.G.; Pinessi, L.; et al. Is HCRTR2 a genetic risk factor for Alzheimer’s disease? Dement. Geriatr. Cogn. Disord. 2014. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.M.; Kratzsch, J.; Gertz, H.J.; Tittmann, M.; Jahn, I.; Pietsch, U.C.; Kaisers, U.X.; Thiery, J.; Hegerl, U.; Schönknecht, P. Cerebrospinal Fluid Melanin-Concentrating Hormone (MCH) and Hypocretin-1 (HCRT-1, Orexin-A) in Alzheimer’s Disease. PLoS ONE 2013, 8, e63136. [Google Scholar] [CrossRef]
- Liguori, C.; Romigi, A.; Nuccetelli, M.; Zannino, S.; Sancesario, G.; Martorana, A.; Albanese, M.; Mercuri, N.B.; Izzi, F.; Bernardini, S.; et al. Orexinergic system dysregulation, sleep impairment, and cognitive decline in Alzheimer disease. JAMA Neurol. 2014, 71, 1498–1505. [Google Scholar] [CrossRef] [PubMed]
- Haller, J. The neurobiology of abnormal manifestations of aggression-A review of hypothalamic mechanisms in cats, rodents, and humans. Brain Res. Bull. 2013. [Google Scholar] [CrossRef]
- Venturelli, M.; Sollima, A.; Cè, E.; Limonta, E.; Bisconti, A.V.; Brasioli, A.; Muti, E.; Esposito, F. Effectiveness of Exercise- and Cognitive-Based Treatments on Salivary Cortisol Levels and Sundowning Syndrome Symptoms in Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2016. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adidharma, W.; Deats, S.P.; Ikeno, T.; Lipton, J.W.; Lonstein, J.S.; Yan, L. Orexinergic modulation of serotonin neurons in the dorsal raphe of a diurnal rodent, Arvicanthis niloticus. Horm. Behav. 2019. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, D.; Long, J.E.; Proulx, C.D.; Barandas, R.; Malinow, R.; Welsh, D.K. Genetic Disruption of Circadian Rhythms in the Suprachiasmatic Nucleus Causes Helplessness, Behavioral Despair, and Anxiety-like Behavior in Mice. Biol. Psychiatry 2016, 80, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Tsuchimine, S.; Hattori, K.; Ota, M.; Hidese, S.; Teraishi, T.; Sasayama, D.; Hori, H.; Noda, T.; Yoshida, S.; Yoshida, F.; et al. Reduced plasma orexin-A levels in patients with bipolar disorder. Neuropsychiatr. Dis. Treat. 2019, 15, 2221–2230. [Google Scholar] [CrossRef] [Green Version]
- Thorn, L.; Evans, P.; Cannon, A.; Hucklebridge, F.; Evans, P.; Clow, A. Seasonal differences in the diurnal pattern of cortisol secretion in healthy participants and those with self-assessed seasonal affective disorder. Psychoneuroendocrinology 2011. [Google Scholar] [CrossRef] [PubMed]
- Majrashi, N.A.; Ahearn, T.S.; Waiter, G.D. Brainstem volume mediates seasonal variation in depressive symptoms: A cross sectional study in the UK Biobank cohort. Sci. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Abulseoud, O.A.; Camsari, U.M.; Ruby, C.L.; Mohamed, K.; Abdel Gawad, N.M.; Kasasbeh, A.; Yüksel, M.Y.; Choi, D.S. Lateral hypothalamic kindling induces manic-like behavior in rats: A novel animal model. Int. J. Bipolar Disord. 2014. [Google Scholar] [CrossRef] [Green Version]
- Watson, S.; Gallagher, P.; Ritchie, J.C.; Ferrier, I.N.; Young, A.H. Hypothalamic-pituitary-adrenal axis function in patients with bipolar disorder. Br. J. Psychiatry 2004, 184, 496–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fries, G.R.; Vasconcelos-Moreno, M.P.; Gubert, C.; Dos Santos, B.T.M.Q.; Sartori, J.; Eisele, B.; Ferrari, P.; Fijtman, A.; Rüegg, J.; Gassen, N.C.; et al. Hypothalamic-pituitary-adrenal axis dysfunction and illness progression in bipolar disorder. Int. J. Neuropsychopharmacol. 2015, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, M.T.; Wester, V.L.; Vreeker, A.; Koenders, M.A.; Boks, M.P.; van Rossum, E.F.C.; Spijker, A.T. Higher cortisol levels may proceed a manic episode and are related to disease severity in patients with bipolar disorder. Psychoneuroendocrinology 2020, 119. [Google Scholar] [CrossRef]
- Manaye, K.F.; Lei, D.L.; Tizabi, Y.; Dávila-García, M.I.; Mouton, P.R.; Kelly, P.H. Selective neuron loss in the paraventricular nucleus of hypothalamus in patients suffering from major depression and bipolar disorder. J. Neuropathol. Exp. Neurol. 2005, 64, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Roybal, K.; Theobold, D.; Graham, A.; DiNieri, J.A.; Russo, S.J.; Krishnan, V.; Chakravarty, S.; Peevey, J.; Oehrlein, N.; Birnbaum, S.; et al. Mania-like behavior induced by disruption of CLOCK. Proc. Natl. Acad. Sci. USA 2007, 104, 6406–6411. [Google Scholar] [CrossRef] [Green Version]
- Adam, C.L.; Moar, K.M.; Logie, T.J.; Ross, A.W.; Barrett, P.; Morgan, P.J.; Mercer, J.G. Photoperiod regulates growth, puberty and hypothalamic neuropeptide and receptor gene expression in female Siberian hamsters. Endocrinology 2000. [Google Scholar] [CrossRef] [PubMed]
- Pail, G.; Huf, W.; Pjrek, E.; Winkler, D.; Willeit, M.; Praschak-Rieder, N.; Kasper, S. Bright-light therapy in the treatment of mood disorders. Neuropsychobiology 2011, 64, 152–162. [Google Scholar] [CrossRef]
- Rosen, L.N.; Targum, S.D.; Terman, M.; Bryant, M.J.; Hoffman, H.; Kasper, S.F.; Hamovit, J.R.; Docherty, J.P.; Welch, B.; Rosenthal, N.E. Prevalence of seasonal affective disorder at four latitudes. Psychiatry Res. 1990, 31, 131–144. [Google Scholar] [CrossRef]
- Vandewalle, G.; Hébert, M.; Beaulieu, C.; Richard, L.; Daneault, V.; Garon, M.L.; Leblanc, J.; Grandjean, D.; Maquet, P.; Schwartz, S.; et al. Abnormal hypothalamic response to light in seasonal affective disorder. Biol. Psychiatry 2011, 70, 954–961. [Google Scholar] [CrossRef] [Green Version]
- Kräuchi, K.; Wirz-Justice, A. The four seasons: Food intake frequency in seasonal affective disorder in the course of a year. Psychiatry Res. 1988, 25, 323–338. [Google Scholar] [CrossRef]
- Sandman, N.; Merikanto, I.; Määttänen, H.; Valli, K.; Kronholm, E.; Laatikainen, T.; Partonen, T.; Paunio, T. Winter is coming: Nightmares and sleep problems during seasonal affective disorder. J. Sleep Res. 2016. [Google Scholar] [CrossRef]
- Adidharma, W.; Leach, G.; Yan, L. Orexinergic signaling mediates light-induced neuronal activation in the dorsal raphe nucleus. Neuroscience 2012. [Google Scholar] [CrossRef] [Green Version]
- Deats, S.P.; Adidharma, W.; Lonstein, J.S.; Yan, L. Attenuated orexinergic signaling underlies depression-like responses induced by daytime light deficiency. Neuroscience 2014, 272, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Coque, L.; Cao, J.L.; Kumar, J.; Chakravarty, S.; Asaithamby, A.; Graham, A.; Gordon, E.; Enwright, J.F.; Dileone, R.J.; et al. Knockdown of clock in the ventral tegmental area through RNA interference results in a mixed state of mania and depression-like behavior. Biol. Psychiatry 2010. [Google Scholar] [CrossRef] [Green Version]
- Brami-Cherrier, K.; Lewis, R.G.; Cervantes, M.; Liu, Y.; Tognini, P.; Baldi, P.; Sassone-Corsi, P.; Borrelli, E. Cocaine-mediated circadian reprogramming in the striatum through dopamine D2R and PPARγ activation. Nat. Commun. 2020, 11, 4448. [Google Scholar] [CrossRef] [PubMed]
- Seney, M.L.; Cahill, K.; Enwright, J.F.; Logan, R.W.; Huo, Z.; Zong, W.; Tseng, G.; McClung, C.A. Diurnal rhythms in gene expression in the prefrontal cortex in schizophrenia. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tognin, S.; Rambaldelli, G.; Perlini, C.; Bellani, M.; Marinelli, V.; Zoccatelli, G.; Alessandrini, F.; Pizzini, F.B.; Beltramello, A.; Terlevic, R.; et al. Enlarged hypothalamic volumes in schizophrenia. Psychiatry Res. Neuroimaging 2012. [Google Scholar] [CrossRef] [PubMed]
- Busch, J.R.; Jacobsen, C.; Lynnerup, N.; Banner, J.; Møller, M. Expression of vasopressin mRNA in the hypothalamus of individuals with a diagnosis of schizophrenia. Brain Behav. 2019. [Google Scholar] [CrossRef] [Green Version]
- An, K.; Zhao, H.; Miao, Y.; Xu, Q.; Li, Y.F.; Ma, Y.Q.; Shi, Y.M.; Shen, J.W.; Meng, J.J.; Yao, Y.G.; et al. A circadian rhythm-gated subcortical pathway for nighttime-light-induced depressive-like behaviors in mice. Nat. Neurosci. 2020, 23, 869–880. [Google Scholar] [CrossRef]
- Walker, W.H.; Walton, J.C.; DeVries, A.C.; Nelson, R.J. Circadian rhythm disruption and mental health. Transl. Psychiatry 2020, 10, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasahara, T.; Kubota, M.; Miyauchi, T.; Noda, Y.; Mouri, A.; Nabeshima, T.; Kato, T. Mice with neuron-specific accumulation of mitochondrial DNA mutations show mood disorder-like phenotypes. Mol. Psychiatry 2006. [Google Scholar] [CrossRef] [PubMed]
- Logan, R.W.; Parekh, P.K.; Kaplan, G.N.; Becker-Krail, D.D.; Williams, W.P.; Yamaguchi, S.; Yoshino, J.; Shelton, M.A.; Zhu, X.; Zhang, H.; et al. NAD+ cellular redox and SIRT1 regulate the diurnal rhythms of tyrosine hydroxylase and conditioned cocaine reward. Mol. Psychiatry 2019. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Nohara, K.; Park, N.; Park, Y.S.; Guillory, B.; Zhao, Z.; Garcia, J.M.; Koike, N.; Lee, C.C.; Takahashi, J.S.; et al. The Small Molecule Nobiletin Targets the Molecular Oscillator to Enhance Circadian Rhythms and Protect against Metabolic Syndrome. Cell Metab. 2016, 23, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Nohara, K.; Mallampalli, V.; Nemkov, T.; Wirianto, M.; Yang, J.; Ye, Y.; Sun, Y.; Han, L.; Esser, K.A.; Mileykovskaya, E.; et al. Nobiletin fortifies mitochondrial respiration in skeletal muscle to promote healthy aging against metabolic challenge. Nat. Commun. 2019, 10, 3923. [Google Scholar] [CrossRef] [Green Version]
- Burke, A.C.; Sutherland, B.G.; Telford, D.E.; Morrow, M.R.; Sawyez, C.G.; Edwards, J.Y.; Drangova, M.; Huff, M.W. Intervention with citrus flavonoids reverses obesity and improves metabolic syndrome and atherosclerosis in obese Ldlr / mice. J. Lipid Res. 2018, 59, 1714–1728. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, V.; Gandasi, N.R.; Sage, D.; Tengholm, A.; Barg, S.; Dibner, C. In pancreatic islets from type 2 diabetes patients, the dampened circadian oscillators lead to reduced insulin and glucagon exocytosis. Proc. Natl. Acad. Sci. USA 2020, 117, 2484–2495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakshit, K.; Matveyenko, A.V. Induction of Core Circadian Clock Transcription Factor Bmal1 Enhances β Cell Function and Protects Against Obesity-Induced Glucose Intolerance. Diabetes 2020. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.J.; Nakamura, W.; Yamazaki, S.; Kudo, T.; Cutler, T.; Colwell, C.S.; Block, G.D. Age-related decline in circadian output. J. Neurosci. 2011. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Solanas, G.; Peixoto, F.O.; Bee, L.; Symeonidi, A.; Schmidt, M.S.; Brenner, C.; Masri, S.; Benitah, S.A.; Sassone-Corsi, P. Circadian Reprogramming in the Liver Identifies Metabolic Pathways of Aging. Cell 2017, 170, 664–677.e11. [Google Scholar] [CrossRef]
- Zwighaft, Z.; Aviram, R.; Shalev, M.; Rousso-Noori, L.; Kraut-Cohen, J.; Golik, M.; Brandis, A.; Reinke, H.; Aharoni, A.; Kahana, C.; et al. Circadian Clock Control by Polyamine Levels through a Mechanism that Declines with Age. Cell Metab. 2015, 22, 874–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, S.; Amir, S. The aging clock: Circadian rhythms and later life. J. Clin. Investig. 2017, 127, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.C.; Guarente, L. XSIRT1 mediates central circadian control in the SCN by a mechanism that decays with aging. Cell 2013, 153, 1448. [Google Scholar] [CrossRef] [Green Version]
- Levine, D.C.; Hong, H.; Weidemann, B.J.; Ramsey, K.M.; Affinati, A.H.; Schmidt, M.S.; Cedernaes, J.; Omura, C.; Braun, R.; Lee, C.; et al. NAD+ Controls Circadian Reprogramming through PER2 Nuclear Translocation to Counter Aging. Mol. Cell 2020. [Google Scholar] [CrossRef]






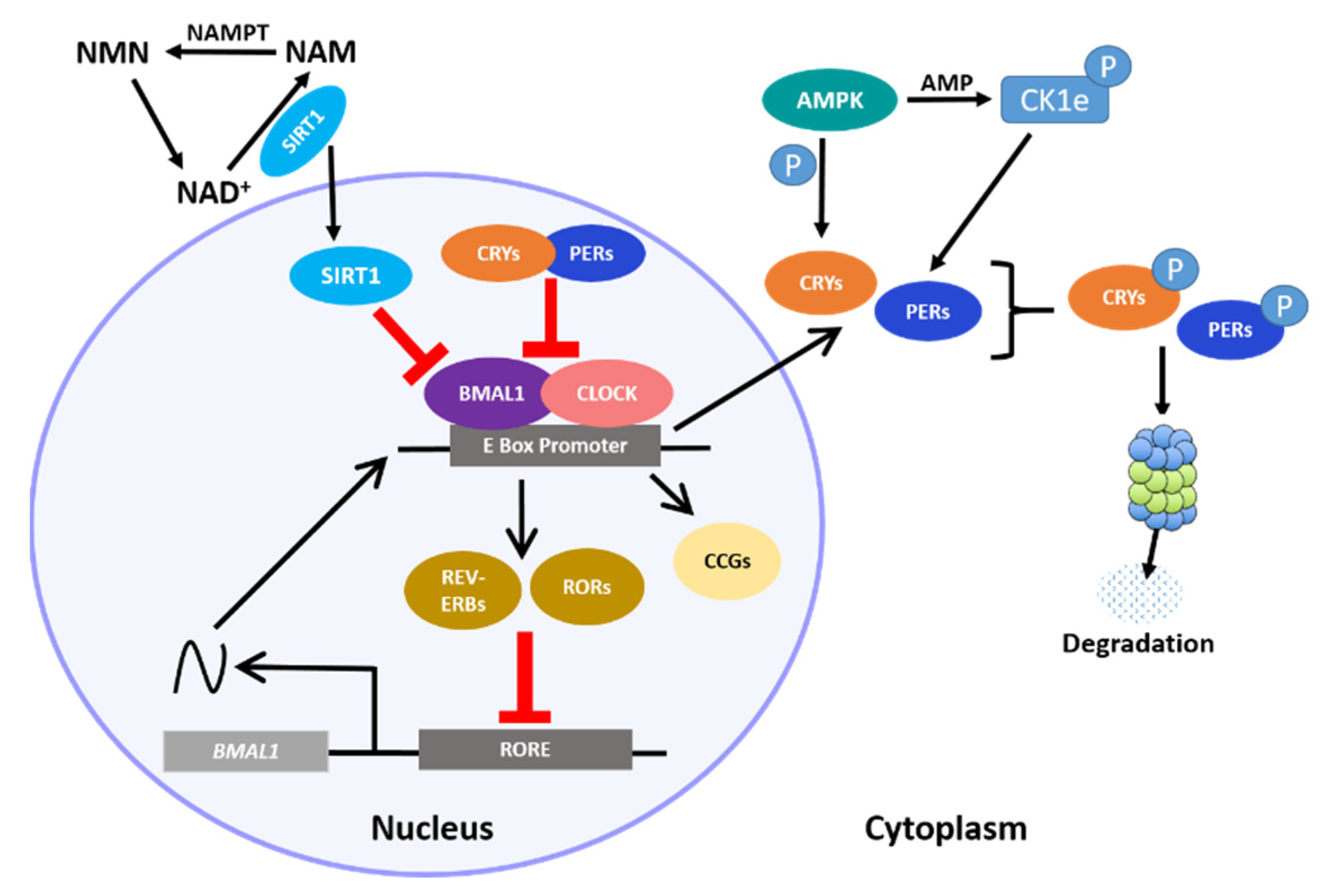{kind=link}
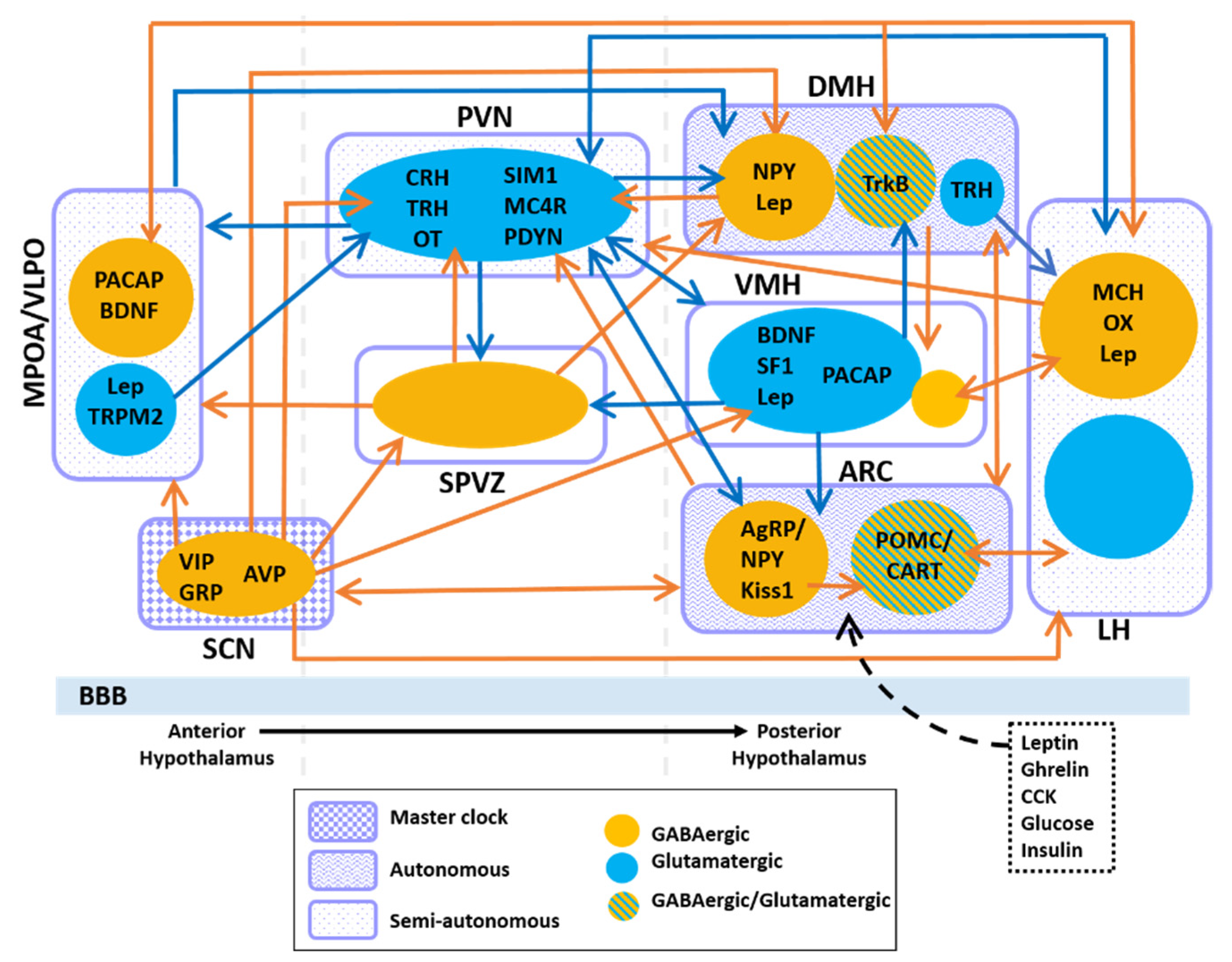{kind=link}
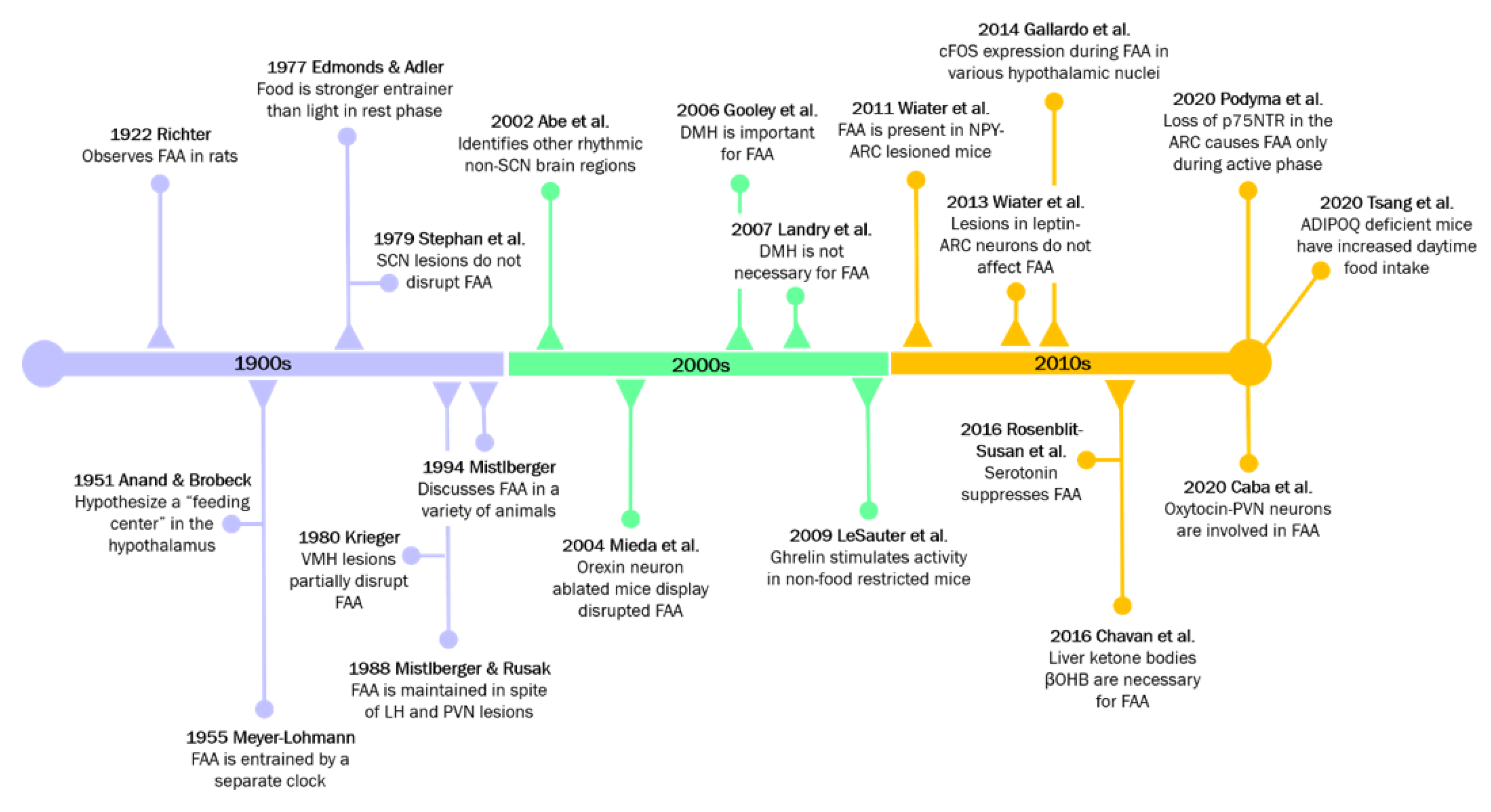{kind=link}
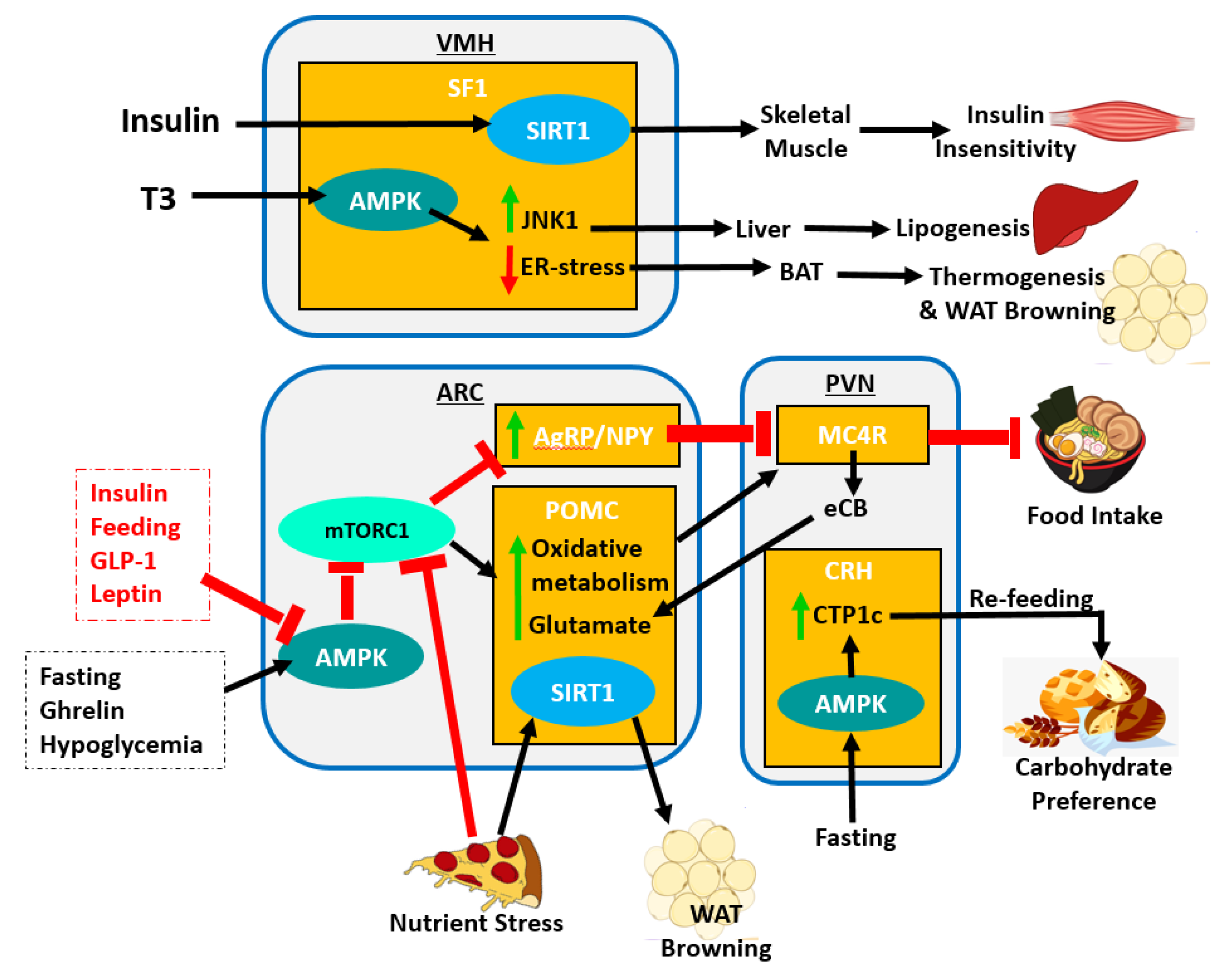{kind=link}
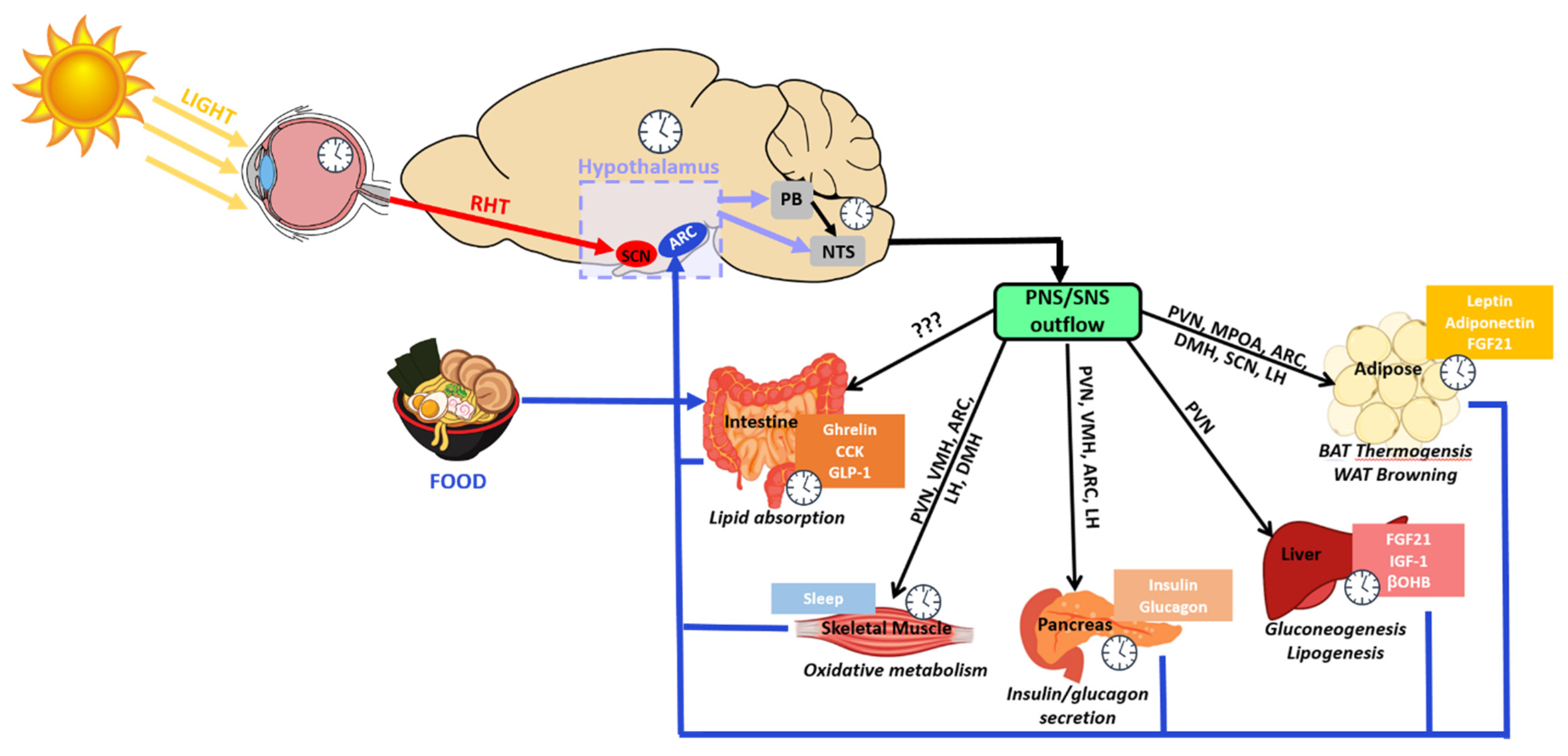{kind=link}
| Neurological Disease/Disorder | Rhythmic Regions Involved | Evidence | References |
|---|---|---|---|
| Alzheimer’s Disease (AD) | SCN |
| [291,293] [292] [314] [315] [316] |
| PVN |
| [317] [318] [319] [320] | |
| LH |
| [294] [321] [322] [323] | |
| Sundowning Syndrome | SCN |
| [302] |
| PVN |
| [324] | |
| VMH |
| [325] | |
| LH |
| [324] [326] | |
| Seasonal Affective Disorder (SAD) | SCN |
| [327] [328] |
| PVN |
| [329] [311] | |
| LH |
| [311] [328] | |
| Brainstem |
| [330] | |
| Bipolar Disorder (BD) | LH |
| [312] [331] [332] |
| PVN |
| [27,333] [334] [335] | |
| VTA |
| [336] | |
| Schizophrenia | dlPFC |
| [337] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Drunen, R.; Eckel-Mahan, K. Circadian Rhythms of the Hypothalamus: From Function to Physiology. Clocks & Sleep 2021, 3, 189-226. https://doi.org/10.3390/clockssleep3010012
Van Drunen R, Eckel-Mahan K. Circadian Rhythms of the Hypothalamus: From Function to Physiology. Clocks & Sleep. 2021; 3(1):189-226. https://doi.org/10.3390/clockssleep3010012
Chicago/Turabian StyleVan Drunen, Rachel, and Kristin Eckel-Mahan. 2021. "Circadian Rhythms of the Hypothalamus: From Function to Physiology" Clocks & Sleep 3, no. 1: 189-226. https://doi.org/10.3390/clockssleep3010012
APA StyleVan Drunen, R., & Eckel-Mahan, K. (2021). Circadian Rhythms of the Hypothalamus: From Function to Physiology. Clocks & Sleep, 3(1), 189-226. https://doi.org/10.3390/clockssleep3010012