Disorders of Arousal: A Chronobiological Perspective



Abstract
:1. Background
2. Rebound of Deep Sleep
2.1. Acute Sleep Deprivation: Sleep Deprivation Schedules in Laboratory Studies
2.2. Chronic Sleep Deprivation or Biological Misalignment: Shift or Rotational Working
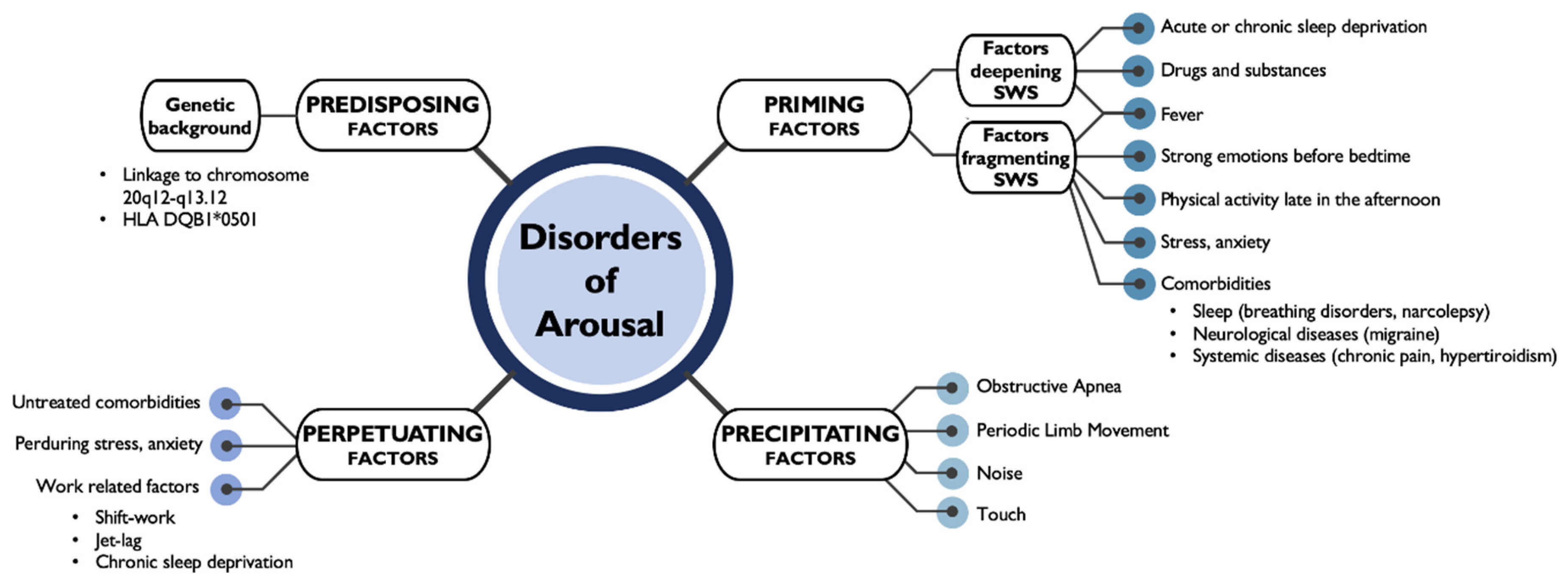
3. Sleep Disruption: Disorders of Arousal
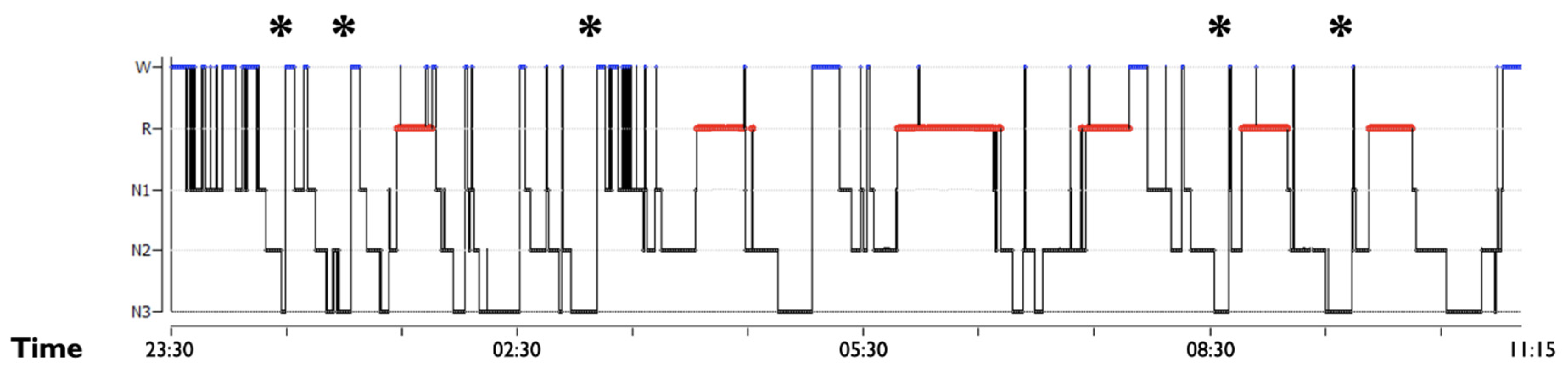
3.1. SWS Intrinsic Fragmentation
3.2. SWS Microstructural Dysfunction: SWS Distribution, Delta Spectral Power and Sleep Spindles Dynamic
3.3. Fragmentation Due to Sleep and Medical Disorders and Substances
4. Circadian Rhythm and Chronotype
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Castelnovo, A.; Lopez, R.; Proserpio, P.; Nobili, L.; Dauvilliers, Y. NREM sleep parasomnias as disorders of sleep-state dissociation. Nat. Rev. Neurol. 2018, 14, 470–481. [Google Scholar] [CrossRef] [PubMed]
- American Academy of Sleep Medicine. International Classification of Sleep Disorders, 3rd ed.; American Academy of Sleep Medicine: Darien, IL, USA, 2014. [Google Scholar]
- Stallman, H.M.; Kohler, M. Prevalence of Sleepwalking: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0164769. [Google Scholar] [CrossRef] [PubMed]
- Silber, M.H. Parasomnias Occurring in Non–Rapid Eye Movement Sleep. Contin. Lifelong Learn. Neurol. 2020, 26, 946–962. [Google Scholar] [CrossRef] [PubMed]
- Hublin, C.; Kapriob, J. Genetic aspects and genetic epidemiology of parasomnias. Sleep Med. Rev. 2003, 7, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Hublin, C.; Kaprio, J.; Partinen, M.; Heikkila, K.; Koskenvuo, M. Prevalence and Genetics of Sleepwalking: A Population-based Twin Study. Neurology 1997, 48, 177–181. [Google Scholar] [CrossRef]
- Petit, D.; Pennestri, M.-H.; Paquet, J.; Desautels, A.; Zadra, A.; Vitaro, F.; Tremblay, R.E.; Boivin, M.; Montplaisir, J. Childhood Sleepwalking and Sleep Terrors. JAMA Pediatr. 2015, 169, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Licis, A.K.; Desruisseau, D.M.; Yamada, K.A.; Duntley, S.P.; Gurnett, C.A. Novel genetic findings in an extended family pedigree with sleepwalking. Neurology 2010, 76, 49–52. [Google Scholar] [CrossRef] [Green Version]
- Lecendreux, M.; Bassetti, C.L.; Dauvilliers, Y.; Mayer, G.; Neidhart, E.; Tafti, M. HLA and genetic susceptibility to sleepwalking. Mol. Psychiatry 2003, 8, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Heidbreder, A.; Frauscher, B.; Mitterling, T.; Boentert, M.; Schirmacher, A.; Hörtnagl, P.; Schennach, H.; Massoth, C.; Happe, S.; Mayer, G.; et al. Not Only Sleepwalking But NREM Parasomnia Irrespective of the Type Is Associated with HLA DQB1*05:01. J. Clin. Sleep Med. 2016, 12, 565–570. [Google Scholar] [CrossRef] [Green Version]
- La Bassetti, C.; Vella, S.; Donati, F.; Wielepp, P.; Weder, B. SPECT during sleepwalking. Lancet 2000, 356, 484–485. [Google Scholar] [CrossRef]
- Gibbs, S.A.; Proserpio, P.; Terzaghi, M.; Pigorini, A.; Sarasso, S.; Russo, G.L.; Tassi, L.; Nobili, L. Sleep-related epileptic behaviors and non-REM-related parasomnias: Insights from stereo-EEG. Sleep Med. Rev. 2016, 25, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zadra, A.; Desautels, A.; Petit, D.; Montplaisir, J. Somnambulism: Clinical aspects and pathophysiological hypotheses. Lancet Neurol. 2013, 12, 285–294. [Google Scholar] [CrossRef]
- Leung, A.K.C.; Leung, A.A.; Wong, A.H.; Hon, K.L. Sleep Terrors: An Updated Review. Curr. Pediatr. Rev. 2020, 16, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.; Kales, A.; Lehmann, D.; Zweizig, J.R. Somnambulism: All-Night Electroencephalographic Studies. Science 1965, 148, 975–977. [Google Scholar] [CrossRef]
- Zucconi, M.; Oldani, A.; Ferini-Strambi, L.; Smirne, S. Arousal fluctuations in non-rapid eye movement parasomnias: The role of cyclic alternating pattern as a measure of sleep instability. J. Clin. Neurophysiol. 1995, 12, 147–154. [Google Scholar] [CrossRef]
- Proserpio, P.; Loddo, G.; Zubler, F.; Ferini-Strambi, L.; Licchetta, L.; Bisulli, F.; Tinuper, P.; Agostoni, E.C.; Bassetti, C.; Tassi, L.; et al. Polysomnographic features differentiating disorder of arousals from sleep-related hypermotor epilepsy. Sleep 2019, 42, zsz166. [Google Scholar] [CrossRef]
- Pressman, M.R. Factors that predispose, prime and precipitate NREM parasomnias in adults: Clinical and forensic implications. Sleep Med. Rev. 2007, 11, 5–30. [Google Scholar] [CrossRef]
- Espa, F.; Ondze, B.; Deglise, P.; Billiard, M.; Besset, A. Sleep architecture, slow wave activity, and sleep spindles in adult patients with sleepwalking and sleep terrors. Clin. Neurophysiol. 2000, 111, 929–939. [Google Scholar] [CrossRef]
- Joncas, S.; Zadra, A.; Paquet, J.; Montplaisir, J. The value of sleep deprivation as a diagnostic tool in adult sleepwalkers. Neurology 2002, 58, 936–940. [Google Scholar] [CrossRef]
- Guilleminault, C.; Poyares, D.; Abat, F.; Palombini, L. Sleep and wakefulness in somnambulism. J. Psychosom. Res. 2001, 51, 411–416. [Google Scholar] [CrossRef]
- Loddo, G.; Baldassarri, L.; Zenesini, C.; Licchetta, L.; Bisulli, F.; Cirignotta, F.; Mondini, S.; Tinuper, P.; Provini, F. Seizures with paroxysmal arousals in sleep-related hypermotor epilepsy (SHE): Dissecting epilepsy from NREM parasomnias. Epilepsia 2020, 61, 2194–2202. [Google Scholar] [CrossRef] [PubMed]
- Lopez, R.; Shen, Y.; Chenini, S.; Rassu, A.L.; Evangelista, E.; Barateau, L.; Jaussent, I.; Dauvilliers, Y. Diagnostic criteria for disorders of arousal: A video-polysomnographic assessment. Ann. Neurol. 2018, 83, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Lopez, R.; Jaussent, I.; Dauvilliers, Y. Objective daytime sleepiness in patients with somnambulism or sleep terrors. Neurology 2014, 83, 2070–2076. [Google Scholar] [CrossRef] [PubMed]
- Iranzo, A. Parasomnias and Sleep-Related Movement Disorders in Older Adults. Sleep Med. Clin. 2018, 13, 51–61. [Google Scholar] [CrossRef]
- Gau, S.S.-F.; Chiang, H.-L. Sleep Problems and Disorders among Adolescents with Persistent and Subthreshold Attention-deficit/Hyperactivity Disorders. Sleep 2009, 32, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Ming, X.; Sun, Y.-M.; Nachajon, R.V.; Brimacombe, M.B.; Walters, A.S. Prevalence of Parasomnia in Autistic Children with Sleep Disorders. Clin. Med. Insights Pediatr. 2009, 3, 1–10. [Google Scholar] [CrossRef]
- Walters, A.S.; Silvestri, R.; Zucconi, M.; Chandrashekariah, R.; Konofal, E. Review of the Possible Relationship and Hypothetical Links Between Attention Deficit Hyperactivity Disorder (ADHD) and the Simple Sleep Related Movement Disorders, Parasomnias, Hypersomnias, and Circadian Rhythm Disorders. J. Clin. Sleep Med. 2008, 4, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Hrozanova, M.; Morrison, I.; Riha, R.L. Adult NREM Parasomnias: An Update. Clocks Sleep 2018, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Bruni, O.; Ferri, R.; Novelli, L.; Finotti, E.; Miano, S.; Guilleminault, C. NREM sleep instability in children with sleep terrors: The role of slow wave activity interruptions. Clin. Neurophysiol. 2008, 119, 985–992. [Google Scholar] [CrossRef]
- Castelnovo, A.; Riedner, B.A.; Smith, R.F.; Tononi, G.; Boly, M.; Benca, R.M. Scalp and Source Power Topography in Sleepwalking and Sleep Terrors: A High-Density EEG Study. Sleep 2016, 39, 1815–1825. [Google Scholar] [CrossRef] [Green Version]
- Flamand, M.; Boudet, S.; Lopes, R.; Vignal, J.-P.; Reyns, N.; Charley-Monaca, C.; Peter-Derex, L.; Szurhaj, W. Confusional arousals during non-rapid eye movement sleep: Evidence from intracerebral recordings. Sleep 2018, 41, zsy139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, P.M.-È.; Carrier, J.; Lina, J.-M.; Fortin, B.M.; Gosselin, N.; Montplaisir, J.; Zadra, A. EEG Functional Connectivity Prior to Sleepwalking: Evidence of Interplay Between Sleep and Wakefulness. Sleep 2017, 40, zsx024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skorucak, J.; Arbon, E.L.; Dijk, D.-J.; Achermann, P. Response to chronic sleep restriction, extension, and subsequent total sleep deprivation in humans: Adaptation or preserved sleep homeostasis? Sleep 2018, 41, zsy078. [Google Scholar] [CrossRef] [PubMed]
- Mayer, G.; Neissner, V.; Schwarzmayr, P.; Meier-Ewert, K. Sleep deprivation in somnambulism. Effect of arousal, deep sleep and sleep stage changes. Nervenarzt 1998, 69, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Guilleminault, C.; Moscovitch, A.; Leger, D. Forensic Sleep Medicine: Nocturnal Wandering and Violence. Sleep 1995, 18, 740–748. [Google Scholar] [CrossRef] [Green Version]
- Pilon, M.; Montplaisir, J.; Zadra, A. Precipitating factors of somnambulism: Impact of sleep deprivation and forced arousals. Neurology 2008, 70, 2284–2290. [Google Scholar] [CrossRef] [PubMed]
- Zadra, A.; Pilon, M.; Montplaisir, J. Polysomnographic diagnosis of sleepwalking: Effects of sleep deprivation. Ann. Neurol. 2008, 63, 513–519. [Google Scholar] [CrossRef]
- Hubbard, J.; Gent, T.C.; Hoekstra, M.M.B.; Emmenegger, Y.; Mongrain, V.; Landolt, H.-P.; Adamantidis, A.R.; Franken, P. Rapid fast-delta decay following prolonged wakefulness marks a phase of wake-inertia in NREM sleep. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Ohayon, M.; Mahowald, M.W.; Dauvilliers, Y.; Krystal, A.D.; Leger, D. Prevalence and comorbidity of nocturnal wandering in the US adult general population. Neurology 2012, 78, 1583–1589. [Google Scholar] [CrossRef] [Green Version]
- Ohayon, M.M.; Guilleminault, C.; Priest, R.G. Night Terrors, Sleepwalking, and Confusional Arousals in the General Population. J. Clin. Psychiatry 1999, 60, 268–276. [Google Scholar] [CrossRef]
- Bjorvatn, B.; Grønli, J.; Pallesen, S. Prevalence of different parasomnias in the general population. Sleep Med. 2010, 11, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Menezes, M.C.R.; Pires, M.L.N.; Benedito-Silva, A.A.; Tufik, S. Sleep Parameters among Offshore Workers: An Initial Assessment in the Campos Basin, Rio De Janeiro, Brazil. Chrono Int. 2004, 21, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Bjorvatn, B.; Magerøy, N.; Moen, B.E.; Pallesen, S.; Waage, M.S. Parasomnias are more frequent in shift workers than in day workers. Chrono Int. 2015, 32, 1352–1358. [Google Scholar] [CrossRef]
- Lugaresi, E.; Provini, F.; Montagna, P. The neuroanatomy of sleep. Considerations on the role of the thalamus in sleep and a proposal for a caudorostral organization. Eur. J. Anat. 2004, 8, 85–93. [Google Scholar]
- Crunelli, V.; Lőrincz, M.L.; Connelly, W.M.; David, F.; Hughes, S.W.; Lambert, R.C.; LeResche, N.; Errington, A.C. Dual function of thalamic low-vigilance state oscillations: Rhythm-regulation and plasticity. Nat. Rev. Neurosci. 2018, 19, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Honjoh, S.; Sasai, S.; Schiereck, S.; Nagai, H.; Tononi, G.; Cirelli, C. Regulation of cortical activity and arousal by the matrix cells of the ventromedial thalamic nucleus. Nat. Commun. 2018, 9, 2100. [Google Scholar] [CrossRef]
- Satpute, A.B.; Kragel, P.A.; Barrett, L.F.; Wager, T.D.; Bianciardi, M. Deconstructing arousal into wakeful, autonomic and affective varieties. Neurosci. Lett. 2019, 693, 19–28. [Google Scholar] [CrossRef]
- Zou, G.; Xu, J.; Zhou, S.; Liu, J.; Su, Z.H.; Zou, Q.; Gao, J. Functional MRI of Arousals in Non-rapid Eye Movement Sleep. Sleep 2020, 43, zsz218. [Google Scholar] [CrossRef]
- Peter-Derex, L.; Magnin, M.; Bastuji, H. Heterogeneity of arousals in human sleep: A stereo-electroencephalographic study. NeuroImage 2015, 123, 229–244. [Google Scholar] [CrossRef]
- Montagna, P.; Provini, F.; Bisulli, F.; Tinuper, P. Nocturnal epileptic seizures versus the arousal parasomnias. Somnologie Schlafforschung Schlafmed. 2008, 12, 25–37. [Google Scholar] [CrossRef]
- Gaudreau, H.; Joncas, S.; Zadra, A.; Montplaisir, J.Y. Dynamics of slow-wave activity during the NREM sleep of sleepwalkers and control subjects. Sleep 2000, 23, 755–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, R.; Laganière, C.; Chenini, S.; Rassu, A.L.; Evangelista, E.; Barateau, L.; Jaussent, I.; Dauvilliers, Y. Video-Polysomnographic Assessment for the Diagnosis of Disorders of Arousal in Children. Neurology 2021, 96, e121–e130. [Google Scholar] [CrossRef] [PubMed]
- Guilleminault, C. Hypersynchronous Slow Delta, Cyclic Alternating Pattern and Sleepwalking. Sleep 2006, 29, 14–15. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.A.; Daan, S.; Wirz-Justice, A.; DeBoer, T. The two-process model of sleep regulation: A reappraisal. J. Sleep Res. 2016, 25, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Loddo, G.; Sessagesimi, E.; Mignani, F.; Cirignotta, F.; Mondini, S.; Licchetta, L.; Bisulli, F.; Tinuper, P.; Provini, F. Specific motor patterns of arousal disorders in adults: A video-polysomnographic analysis of 184 episodes. Sleep Med. 2018, 41, 102–109. [Google Scholar] [CrossRef]
- Weiner, O.M.; Dang-Vu, T.T. Spindle Oscillations in Sleep Disorders: A Systematic Review. Neural Plast. 2016, 2016, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, L.M.; Lüthi, A. Sleep Spindles: Mechanisms and Functions. Physiol. Rev. 2020, 100, 805–868. [Google Scholar] [CrossRef]
- Guilleminault, C.; Palombini, L.; Pelayo, R.; Chervin, R.D. Sleepwalking and sleep terrors in prepubertal children: What triggers them? Pediatrics 2003, 111, e17–e25. [Google Scholar] [CrossRef] [Green Version]
- Guilleminault, C.; Kirisoglu, C.; Bao, G.; Arias, V.; Chan, A.; Li, K.K. Adult chronic sleepwalking and its treatment based on polysomnography. Brain 2005, 128, 1062–1069. [Google Scholar] [CrossRef] [Green Version]
- Bargiotas, P.; Arnet, I.; Frei, M.; Schindler, K.; Baumann, C.R.; La Bassetti, C. Demographic, Clinical and Polysomnographic Characteristics of Childhood- and Adult-Onset Sleepwalking in Adults. Eur. Neurol. 2017, 78, 307–311. [Google Scholar] [CrossRef] [Green Version]
- Bušková, J.; Piško, J.; Pastorek, L.; Šonka, K. The Course and Character of Sleepwalking in Adulthood: A Clinical and Polysomnographic Study. Behav. Sleep Med. 2014, 13, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Drakatos, P.; Marples, L.; Muza, R.; Higgins, S.; Nesbitt, A.; Dongol, E.M.; Macavei, R.; Gnoni, V.; Carbonell, L.P.; Duncan, I.; et al. Video polysomnographic findings in non-rapid eye movement parasomnia. J. Sleep Res. 2018, 28, e12772. [Google Scholar] [CrossRef] [PubMed]
- Kales, J.D.; Kales, A.; Soldatos, C.R.; Chamberlin, K.; Martin, E.D. Sleepwalking and night terrors related to febrile illness. Am. J. Psychiatry 1979, 136, 1214–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, C.; Dooley, J.M.; Gordon, K. Fever-associated confusional arousal. Eur. J. Nucl. Med. Mol. Imaging 2004, 163, 696–697. [Google Scholar] [CrossRef] [PubMed]
- Lopez, R.; Jaussent, I.; Dauvilliers, Y. Pain in Sleepwalking: A Clinical Enigma. Sleep 2015, 38, 1693–1698. [Google Scholar] [CrossRef] [Green Version]
- Giuliano, L.; Fatuzzo, D.; Mainieri, G.; La Vignera, S.; Sofia, V.; Zappia, M. Adult-Onset Sleepwalking Secondary to Hyperthyroidism: Polygraphic Evidence. J. Clin. Sleep Med. 2018, 14, 285–287. [Google Scholar] [CrossRef] [Green Version]
- Barabas, G.; Ferrari, M.; Matthews, W.S. Childhood migraine and somnambulism. Neurology 1983, 33, 948. [Google Scholar] [CrossRef]
- Juszczak, G.R.; Swiergiel, A.H. Serotonergic hypothesis of sleepwalking. Med. Hypotheses 2005, 64, 28–32. [Google Scholar] [CrossRef]
- Colrain, I.M.; Nicholas, C.L.; Baker, F.C. Alcohol and the sleeping brain. Interv. Neuroradiol. 2014, 125, 415–431. [Google Scholar] [CrossRef] [Green Version]
- Pressman, M.R.; Mahowald, M.W.; Schenck, C.H.; Bornemann, M.C. Alcohol-induced sleepwalking or confusional arousal as a defense to criminal behavior: A review of scientific evidence, methods and forensic considerations. J. Sleep Res. 2007, 16, 198–212. [Google Scholar] [CrossRef]
- Pressman, M.R. The neurophysiological and neurochemical effects of alcohol on the brain are inconsistent with current evidence based models of sleepwalking. Sleep Med. Rev. 2019, 43, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Proserpio, P.; Terzaghi, M.; Manni, R.; Nobili, L. Drugs Used in Parasomnia. Sleep Med. Clin. 2020, 15, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Reis, C.; Pilz, L.K.; Keller, L.K.; Paiva, T.; Roenneberg, T. Social timing influences sleep quality in patients with sleep disorders. Sleep Med. 2020, 71, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, M.; Nagamitsu, S.; Iwasaki, M.; Iemura, A.; Yamashita, Y.; Maeda, M.; Kitani, S.; Kakuma, T.; Uchimura, N.; Matsuishi, T. High incidence of sleep problems in children with developmental disorders: Results of a questionnaire survey in a Japanese elementary school. Brain Dev. 2014, 36, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Durmuş, F.B.; Arman, A.R.; Ayaz, A.B. Chronotype and its relationship with sleep disorders in children with attention deficit hyperactivity disorder. Chrono Int. 2017, 34, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Jafar, N.K.; Tham, E.K.; Eng, D.Z.; Goh, D.Y.; Teoh, O.-H.; Lee, Y.S.; Shek, L.P.-C.; Yap, F.; Chong, Y.S.; Meaney, M.J.; et al. The association between chronotype and sleep problems in preschool children. Sleep Med. 2017, 30, 240–244. [Google Scholar] [CrossRef]
- Eid, B.; Saleh, M.B.; Melki, I.; Torbey, P.-H.; Najem, J.; Saber, M.; El Osta, N.; Khabbaz, L.R. Evaluation of Chronotype Among Children and Associations With BMI, Sleep, Anxiety, and Depression. Front. Neurol. 2020, 11, 416. [Google Scholar] [CrossRef]
- Kayaba, M.; Matsushita, T.; Enomoto, M.; Kanai, C.; Katayama, N.; Inoue, Y.; Sasai-Sakuma, T. Impact of sleep problems on daytime function in school life: A cross-sectional study involving Japanese university students. BMC Public Health 2020, 20, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kerkhof, G.A. Epidemiology of sleep and sleep disorders in The Netherlands. Sleep Med. 2017, 30, 229–239. [Google Scholar] [CrossRef]
- Wei, N.S.; Praharaj, S.K. Chronotypes and its association with psychological morbidity and childhood parasomnias. Indian J. Psychiatry 2020, 61, 598–604. [Google Scholar] [CrossRef]
- Reid, K.J.; Jaksa, A.A.; Eisengart, J.B.; Baron, K.G.; Lu, B.; Kane, P.; Kang, J.; Zee, P.C. Systematic evaluation of Axis-I DSM diagnoses in delayed sleep phase disorder and evening-type circadian preference. Sleep Med. 2012, 13, 1171–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivelä, L.; Papadopoulos, M.R.; Antypa, N. Chronotype and Psychiatric Disorders. Curr. Sleep Med. Rep. 2018, 4, 94–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, M.C.A.; Abreu, R.L.; Neto, V.B.L.; De Bruin, P.F.; De Bruin, V.M. Chronotype and circadian rhythm in bipolar disorder: A systematic review. Sleep Med. Rev. 2017, 34, 46–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passos, G.S.; Santana, M.G.; Poyares, D.; D’Aurea, C.V.; Teixeira, A.A.; Tufik, S.; De Mello, M.T. Chronotype and anxiety are associated in patients with chronic primary insomnia. Rev. Bras. Psiquiatr. 2017, 39, 183–186. [Google Scholar] [CrossRef] [Green Version]
- Lopez, R.; Jaussent, I.; Scholz, S.; Bayard, S.; Montplaisir, J.; Dauvilliers, Y. Functional Impairment in Adult Sleepwalkers: A Case-Control Study. Sleep 2013, 36, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Hartman, D.; Crisp, A.H.; Sedgwick, P.; Borrow, S. Is there a dissociative process in sleepwalking and night terrors? Postgrad. Med. J. 2001, 77, 244–249. [Google Scholar] [CrossRef]
- Stefani, A.; Heidbreder, A.; Brandauer, E.; Guaita, M.; Neier, L.-M.; Mitterling, T.; Santamaria, J.; Iranzo, A.; Videnovic, A.; Trenkwalder, C.; et al. Screening for idiopathic REM sleep behavior disorder: Usefulness of actigraphy. Sleep 2018, 41, zsy053. [Google Scholar] [CrossRef]
- Weissová, K.; Škrabalová, J.; Skálová, K.; Červená, K.; Bendová, Z.; Miletínová, E.; Kopřivová, J.; Šonka, K.; Dudysová, D.; Bartoš, A.; et al. Circadian rhythms of melatonin and peripheral clock gene expression in idiopathic REM sleep behavior disorder. Sleep Med. 2018, 52, 1–6. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Confusional Arousal | Sleep Terror | Sleepwalking | |
|---|---|---|---|
| General features | Individuals may sit up in bed, looking around with a confused gaze | Individuals sit up abruptly in bed, with intense fearful expression, screaming or defensive gestures | Individuals may walk in the bedroom, with unpurposeful searching behaviors or engaging in more complex actions |
| Behaviors | |||
| Routine behaviors that occur at inappropriate times | Eye opening, face rubbing, staring, scratching, yawning, searching behaviors | Inconsolable sobbing, anguished face | Calm, quiet ambulation, getting ready for school, driving, moving furniture |
| Inappropriate or nonsensical behaviors | Manipulating EEG equipment or bed blankets | Recoiling from something, spasmodic searching, evasive behaviors | Walking naked on the roof of the garage or sitting in front of a turned-off TV |
| Dangerous or potentially dangerous behaviors | / | Hitting, kicking, falling from the bed | Climbing on a chair, jumping out from a window |
| Sounds | Mumbling, moaning, single words | Screaming, fearful phrases | Sleeptalking |
| Clinical consequences | Minimal | Possible self injuries | Minimal to severe |
| Forensic implications | + | ++ | +++ |
| Time of the night | Usually first third of the night or throughout the night | First third of the night | First third of the night |
| Sleep Stage | |||
| N3 | +++ | +++ | +++ |
| N2 | + | + | + |
| Sleep features | Increased N3 fragmentation | Increased N3 fragmentation | Increased N3 fragmentation |
| EEG | Co-existence of anterior delta waves and posterior wake rhythms | Mostly masked by movement artifact | Mostly masked by movement artifact |
| Autonomic activation | ++ | +++ | ++ |
| EMG activation | + | +++ | +++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mainieri, G.; Loddo, G.; Provini, F. Disorders of Arousal: A Chronobiological Perspective. Clocks & Sleep 2021, 3, 53-65. https://doi.org/10.3390/clockssleep3010004
Mainieri G, Loddo G, Provini F. Disorders of Arousal: A Chronobiological Perspective. Clocks & Sleep. 2021; 3(1):53-65. https://doi.org/10.3390/clockssleep3010004
Chicago/Turabian StyleMainieri, Greta, Giuseppe Loddo, and Federica Provini. 2021. "Disorders of Arousal: A Chronobiological Perspective" Clocks & Sleep 3, no. 1: 53-65. https://doi.org/10.3390/clockssleep3010004
APA StyleMainieri, G., Loddo, G., & Provini, F. (2021). Disorders of Arousal: A Chronobiological Perspective. Clocks & Sleep, 3(1), 53-65. https://doi.org/10.3390/clockssleep3010004