1. Introduction
Cattle welfare and comfort are important considerations when selecting housing and bedding type, particularly in the Upper Midwest of the USA, where cold and snowy winters are the norm. Deep bedded compost barns are a housing system that has been increasing in popularity. Dairy producers reported using these barns for increased cow comfort and longevity [
1]. However, high building costs and environmental concerns have resulted in many farmers investigating options to keep cattle outdoors throughout winter [
2,
3,
4]. In Ireland, cows housed outdoors on outwintering pads through winter showed no negative effect on production [
5] or udder health [
6].
Stable flies are economically significant pests of dairy cattle and other livestock during summer, with national losses to the cattle industry exceeding USD 2 billion annually [
7]. Adult flies are obligate blood feeders, whose painful bites annoy and stress cattle. Stress from biting flies negatively affects weight gain and milk production. These flies develop in decaying organic matter, including soiled animal bedding. Previous studies have shown that soiled straw calf bedding yielded significantly higher numbers of stable flies and house flies than wood chips or sawdust bedding [
8]. Leftover hay from bale feeders is also a significant substrate for developing maggots [
9,
10]. With several types of potential breeding sites, the control of stable flies through source reduction may be difficult.
There are many successful housing systems for dairy cows, and all are designed with cow comfort in mind. Comfortable resting facilities are important for optimum production. Designing adequate housing helps to alleviate stress related to poor cow comfort [
11]. Providing adequate resting time is an important aspect of dairy management, both for production and welfare [
12]. Dairy cows in confinement should lie down for approximately 12 h/day [
13]; however, if facilities are not sufficiently clean or comfortable, cows will often remain standing [
14,
15].
An alternative confinement housing option is the compost bedded pack, which is more commonly used in smaller dairies [
16]. Producers using compost barns have reported increased cow comfort [
17]. Cows may be kept in compost packs during the winter months or throughout the year. These barns are free of stalls and other barriers, allowing unrestricted movement. A barn consists of enough space where each cow can simultaneously lie down with space remaining to allow a walking cow to pass through [
17]. Sawdust is the most frequently used bedding substrate, although alternate materials are acceptable, provided that there is adequate management [
18]. Packs should begin with 30–50 cm of material [
17]. Properly managed compost packs may vary in depth, but should have core temperatures exceeding 40 °C with moisture in the range of 50–60% [
19]. If dry bedding is not available, cows will spend significantly less time lying down [
20].
Faced with potentially high building costs and environmental concerns, many producers are investigating options to keep cattle outdoors throughout winter [
2]. In some countries, beef cattle are often kept outdoors through winter without adverse effects to welfare or growth [
21]. In northern climates, housing in winter must protect cattle from cold weather to maintain production. Cows are cold-hardy and thrive in cooler temperatures. However, protection from the elements should be provided [
22], especially for younger animals that are more susceptible to the cold. Wind and excess moisture are stressors to cows exposed to inclement weather [
23]. Dairy cows exposed to winter temperatures below zero Fahrenheit usually opt to spend time indoors [
24]. According to Tucker et al. [
25], higher body condition scores helped to insulate cows against winter effects and cows position themselves to minimize the surface area exposed to harsh weather. In a controlled research study in Minnesota, cows housed on outdoor straw packs had lower hygiene scores, improved udder health, and greater rumination time compared with cows housed indoors [
3]. Furthermore, outwintered cows were not different regarding production or somatic cell count from the cows housed indoors, and outwintered cows had more net return per day compared to cows housed indoors [
4].
There is extensive literature detailing the effects of the weather on the development, fecundity, population dynamics, and activity of flies [
26,
27]. The weather is an important factor to the activity and feeding behaviors of flies, although results describing how the weather affects activity vary. The temperature is consistently an important variable to fly activity. The feeding activity of stable flies ceases at temperatures below 15 °C [
28]. Smith and Hansens [
29] fed stable flies at varying combinations of temperature and humidity levels in the lab and found the highest percentage of stable flies when feeding at 32 °C with a relative humidity below 43%. Additionally, the lowest percentage of flies was found when feeding at 23 °C with a relative humidity above 75%. Berry and Campbell [
30] noted that, while stable fly feeding behavior was influenced by varying weather effects, feeding was partially dictated by time, regardless of the weather.
Adult stable flies closely resemble house flies but are easily distinguished by piercing mouthparts that protrude from under the head. These flies are obligate blood feeders and are often found on the legs of cows. According to Aguiar-Valgode [
31], the ideal temperature for developing stable fly maggots is 25 °C, and temperatures exceeding 35 °C are harmful to development. While heat does speed development time, moisture is another critical factor [
32]. Substrates that are too dry or too wet are unsuitable for developing maggots. Development is also dependent on live bacterial colonies within the substrate [
33]. More eggs are laid in hay and manure mixtures with live microbial activity as opposed to a sterile substrate of the same mixture [
34]. Maggots can develop in an array of decaying organic material and environmental conditions [
35]. Known breeding sites on a dairy farm include straw bedding in calf hutches [
8], bale hay feeding sites [
10], and accumulated silage [
36]. Adults have been considered pests of cattle kept indoors due to the abundance of soiled bedding, an ideal breeding site for stable flies. However, with the increased use of round bale hay feeding sites in pasture, at present, stable flies are considered important pests of pastured cattle as well [
10].
The control of adult stable flies is difficult [
37]. Knowledge of breeding sites will influence management strategies on a dairy farm, particularly given the range of suitable habitats. Some areas, such as drainage ditches, are more difficult to manage due to limited accessibility. However, facility modifications with fly control in mind, including the use of bedding unsuitable for fly development, can be quite useful to efforts in managing adult populations. Schmidtmann [
38] found significantly reduced densities of stable flies in calf hutches with sawdust bedding as opposed to straw bedding. Removing accumulated waste feed eliminates a prolific breeding site for stable flies. Broce et al. [
9] found that a wide range of stable flies emerged from hay waste at feeding sites, with some sites producing over 3000 flies/m
2.
The present study is part of a larger study to develop sustainable and profitable management strategies of dairy cattle, including winter housing and fly management [
3,
4,
39], which compared organic dairy cows housed in outdoor straw packs and covered sawdust compost barns. The objectives of the study are to characterize ambient stable fly populations in summer around the remaining debris from straw bedding piles and compost bedding piles and to characterize the stable fly populations that emerge from bedding debris piles. If the piles are equally attractive, the expectation is to observe similar numbers of ambient adults surrounding the piles. Comparing the numbers and sizes of flies emerging from the piles provides an understanding of substrate attractivity and quality. The hypothesis is that, when compared with compost piles, straw piles produce higher numbers of larger flies.
2. Materials and Methods
The study was conducted using the certified organic dairy herd at the West Central Research and Outreach Center (WCROC) in Morris, MN. Surrounding the WCROC are organic pastures for grazing and field crops. This site also includes a conventional dairy herd independently managed from the organic herd. Conventional cows were housed in a compost barn adjacent to the study barn for most of the year and transferred to a dry lot for part of each summer. The research herd comprised Holstein and crossbred dairy cows. Daily temperatures and precipitation were recorded at the onsite weather station from November 2012 to October 2014 and were used to characterize the weather from both winters and the following summer grazing seasons.
Results from studies with the same herd and timeframe reported that dairy cows housed on straw packs housed outdoors had lower hygiene scores, less clinical mastitis, and improved udder health compared to cows housed in compost barns [
3]. There were no differences in milk production or somatic cell score between cows housed on straw packs and compost barns; however, the cows housed on straw packs had greater profitability than the cows housed in compost barns [
4].
2.1. Experimental Design
The compared winter housing systems were indoor compost bedding packs and outdoor straw bedding packs. In November of 2012 and 2013, the sites for both treatments were cleaned out and prepared to house the cattle for the upcoming winter. Each treatment had two replicates in both years, which are referred to as “Straw 1, Straw 2” and “Compost 1, Compost 2”. The two replicates were housed in identical straw packs and compost barns (
Figure 1). A group of 22 cows, balanced by breed, parity, and calving date, was randomly assigned to one of the four replicates in December of both years. Each group remained in their assigned housing, except for daily milking twice, and was fed a total mixed ration consisting of organic corn silage, alfalfa silage, organic dry alfalfa hay, organic expelled soybean meal, organic corn, and vitamins and minerals throughout winter. The average temperature of the research dairy herd ranged from −14 to 5 °C [
4]. The same weather environmental conditions affected the cows in both the straw packs and compost barns. The compost barn was a 3-sided barn without insulation and an open side where the cows had to eat outside, which had similar temperature conditions to the cows in the straw packs [
4].
The replicate straw piles were located 100 m apart, with each pile being 100 m away from the milking parlor. Roughly 1500 kg of organic wheat straw was laid to create a pile measuring 12 × 21 m. New straw was added on top of the old straw as needed to maintain a dry lying surface. Cows also had a loafing area to access feed and water. To the north of each straw lot was a natural tree windbreak (
Figure 1).
One existing open front compost barn that was 385 m from the milking parlor was divided in half to create two replicate barns with independently managed groups on each side (
Figure 1). Each barn measured 9 × 12 m. Immediately outside was a concrete floored loafing area where cows had access to feed and water. In December of each year, 1600 kg of new organic sawdust was laid to create a deep bedding pack. Compost was tilled twice each day during milking using a skid loader, and new sawdust was added as needed to maintain a dry lying surface. In February of each year, the compost piles became too high to effectively till using the skid loader, necessitating the removal of 12,700 kg of compost from the barns. The removed material from each compost replicate was piled separately nearby and probed for maggots throughout summer. After the cows were turned out to pasture on 28 May 2013 and 4 June 2014, the remaining straw piles were flagged and measured to calculate the area and create a scaled map of each pile.
2.2. Ambient Adult Fly Population
After the cows vacated their assigned housing, the Olson biting fly traps (
Figure 2A) were placed adjacent to each pile to measure the ambient adult density and test the hypothesis that the two bedding pile types attracted different numbers of adult stable flies. A sheet of Alsynite fiberglass (66 × 30.5 cm) was bent into a cylinder with a surface area of 3000 cm
2 and secured 0.5 m above ground with a wooden stake. Each cylinder was wrapped with a replaceable sticky sleeve, which was secured in place with a binder clip. The Olson traps were placed at the straw pile edges in all cardinal directions and diagonally from the compost barn. The locations of the Olson traps in relation to the debris piles are shown in
Figure 1.
In 2013, the traps on one straw pile and both compost piles were assembled on 22 May after the cows were removed. The Olson traps were assembled on the second straw pile on June 6, after the cows were transferred to pasture. Each pile had four traps placed on compass points for a total of 12 traps. In summer 2014, the traps were assembled on 24 April to document when the flies first became active. An additional four traps were placed 500 to 1000 m away from the leftover piles to evaluate the fly density at distant locations from potential breeding sites. Two of the field traps were placed to the south of the piles, and the remaining two were placed to the north. All sleeves were inspected on a regular basis and replaced as needed, once or twice weekly. Full sleeves were frozen until counted. In both years, the sticky traps were maintained until the second week of October.
To test the hypothesis that ambient adult females around debris piles differ in size and gonotrophic age, the sampling of female flies from the Olson traps began in summer 2014. The Olson traps in all locations were inspected daily throughout the week until 30 adult females were sampled from all locations. In weeks when 30 females could not be sampled due to low catches, the males were collected until head widths of 30 flies were measured. The sampled flies were transferred to the lab for measurement and dissection. The heads were removed, lined up occiput down on a Petri dish, and measured at the widest point using a dissecting microscope at 12×. The bodies of the flies were saved for dissection. The females were dissected on the day of collection in a 0.7% saline solution using the methods outlined by Scholl [
40]. Abdominal terga were gently pried apart using forceps to expose the ovaries. The ovaries were then inspected and scored as being previtellogenic (scored as 0 or 1) or vitellogenic (scored as 2 and above) based on the presence of a visible yolk in the developing follicles. This only occurred in 2014.
2.3. Stable Flies Emerging from the Piles
Funnel emergence traps (
Figure 2B) were placed on the piles to quantify the numbers of stable flies emerging from the remaining debris piles and to test the hypothesis that the two bedding types differ in the numbers and sizes of the flies that emerged. Thirty traps were assembled in each pile after the cows vacated their assigned housing in both years. In both years, random sections of the piles were probed to search for developing maggots with a trowel. The hauled compost debris was also probed for maggots throughout the summer. In 2014, the head widths of emerged flies were measured to examine the substrate quality.
Each emergence trap was a 7.57 L plastic bucket (Leaktite, Leominster, MA, USA) with the bottom removed. A screen-sided 1.89 L funnel (Midwest Can, Melrose Park, IL, USA) was inserted midway into the plastic bucket and capped with a 0.95 L plastic deli container (Delitainer, Lake Forest, IL, USA) to collect the trapped flies (
Figure 2C). A plastic 37 mL portion cup (Solo, Lake Forest, IL, USA) with its bottom removed was glued inside a hole on the bottom of each deli container to help to secure the container to the funnel and to facilitate quick removal and replacement. Each container included a laminated note card coated in a 50% Stikem Special, spread to aid the trapping of the flies. Wooden stakes anchored the bucket traps to the straw piles in the case of high winds (
Figure 2E). Traps in the compost barns were sheltered by walls; as such, wooden stakes were not needed (
Figure 2D). Total possible trap locations within each pile were used to extrapolate the cumulative fly emergence from the entire pile each summer.
Emergence traps were randomly assigned locations on all piles in both years. All the traps were inspected, emptied, and assigned a new random location each week. Containers with flies were retrieved and frozen until the flies were counted. The emergence traps were initially assembled on 14 June 2013, using stratified random sampling. Each pile contained 10 traps at the edge and 20 in the center to determine if more flies emerged from the pile’s edge or center.
In summer 2014, all the emergence traps were assembled using unrestricted random sampling (n = 90). Thirty emergence traps were assembled in each straw pile on 13 May, shortly after the first observation of stable flies on sticky traps. Each compost barn contained 15 emergence traps in 2014, which were set on 28 May after the cows were removed to pasture. The traps could not be assembled until after the cows were removed from their assigned housing. Emergence traps from the compost barns were dismantled in the second week of August in both years after consecutive weeks of not trapping any stable flies. Straw pile traps were maintained until the second week of September of both years as stable flies continued to emerge.
A vial of approximately 30 lab-reared house fly pupae was also planted inside the emergence traps in both summer periods to measure trap efficiency. These marker houseflies were easily distinguished from wild-type flies. Sand was added to the vials with a hole punched in the bottom in 2014 to protect the pupae from weather and predators. Each week, when the emergence traps were reassembled, adult marker flies captured in deli containers and empty puparia inside the vials were counted. These counts were used to calculate the percent of emerged flies that were captured in overhead emergence traps.
To estimate the egg to adult development time for stable flies developing in the debris pile, degree-day models based on the pile temperature were used. The temperatures in 1 of each kind of debris pile were recorded every 30 min in summer 2014 using 4 probe HOBO® temperature loggers (Onset Computer Corp., Bourne, MA, USA) installed on 29 May. Each probe was designated different locations within the pile. In straw, all 4 probes were assembled in the pile’s center at varying depths ranging from 2.5 to 100 cm below the surface. In the compost, 2 probes were placed at the pile’s edge and the other 2 probes at the center to assess possible temperature differences. At both the edge and center positions, the probes were positioned 75 and 2.5 cm below the surface. The HOBO®s were maintained on the pile until the emergence traps were dismantled.
To analyze the differences in the ambient adult population of flies surrounding the piles, a repeated measures nonlinear mixed effects analysis was used on log transformed data and tested for an interaction of treatment and date. Similarly, proportion vitellogenic and mean head widths of flies sampled in 2014 were compared by date and location to assess any differences between bedding types and if flies captured away from piles were significantly different than flies captured next to piles.
In 2013, emergence traps were set using stratified random sampling. Raw catch rates reflected weekly catches, from which log transformed daily catch rates were determined by taking the log of weekly catch rates divided by the number of days that traps were assembled. Summer emergence was calculated using pooled means of catch rates from edge traps and center traps. After determining emergence was not significantly different between edge and center traps, emergence traps were set using unrestricted random sampling in summer 2014.
In both years, analysis was with repeated-measures nonlinear mixed effects on the log transformed daily catch rate to assess differences in the numbers of stable flies emerging by date and pile type. The size of emerged flies was compared by date and location using the same methods previously described for the size of ambient adult flies.
Marker flies were analyzed with a repeated-measures model with the fixed effects of date and treatment and random effects of pile and trap within the pile. To calculate the trap efficiency value, the number of marker flies captured in the deli container were divided by the total emerged flies. After determining the trap efficiency result, stable fly catches were adjusted accordingly by dividing the raw daily catch rates by matching estimates of the trap’s efficiency. The total numbers of stable flies emerging from each pile were extrapolated by multiplying the mean daily catch rate by the number of possible trap locations within a pile. These numbers were used to estimate the cumulative emergence value over the summer and the proportion of the total that emerged at any given time.
Degree-day models were used to compare the timing of emergence and capture of ambient adults on Olson traps by pile type in 2014. Degree days were calculated from the temperatures recorded every 30 min in bedding piles. In both types of piles, one probe was selected for these calculations, based on where stable fly maggots were predicted to develop. In the compost, the center probe located 2.5 cm below the surface was chosen. In the straw, the temperatures from the probe located 2.5 cm below the surface were used until mid-July, when a 2nd shallowest probe was positioned 40 cm below the surface. This switch was performed due to observations that the top layer of the straw was drying out, which would have forced maggots to develop deeper within the pile. One generation was defined as 327.1 degree days, with 7 °C as the lower developmental threshold and 34 °C as the upper developmental threshold. When HOBO® loggers were not in place, the pile temperatures were estimated based on the records obtained from the weather station.
2.4. Statistical Analysis
All analyses began with a full repeated-measures model with fixed effects of date and treatment and random effects of pile and trap within pile. Random effects accounting for less than 15% of the total variation were removed and the analyses proceeded with only the fixed effects. Log transformations were used as needed to satisfy the analytical assumptions of the normal distribution and equal variance. Residual plots were graphed and inspected to confirm these assumptions. Insignificant effects were removed to create a minimally sufficient model. All statistical analyses were conducted using R version 3.0.1, using packages “nlme” and “lme4” (R Foundation for Statistical Computing, Vienna, Austria).
3. Results
3.1. Study Site and Weather
Removed compost debris was piled within 50 m of the compost barn and regularly probed for maggots in the spring and summer of both years. Maggots were never found in the removed debris in either year. The cows were removed from straw bedding packs prior to pasture turnout due to muddy conditions from rainy weather. In 2013, the cows from one replicate straw pile were removed on 22 April, approximately one month prior to the scheduled pasture transfer. In 2014, all the cows in straw piles were removed on 23 April, approximately one month before turning the cows out to pasture. Due to limited housing in both years, the cows remained in compost until being turned out to pasture. Throughout both summers, the conventional dairy herd was independently housed in a compost barn less than 15 m from the study barn. In August 2014, young cows were housed in the compost barns after the emergence traps were dismantled due to limited housing space. Average monthly temperatures during winter (2012–2013) were close to normal averages. However, with snow remaining until the second week of May and temperatures being colder than average temperatures, the growing season was delayed. After spring, the weather was warm through September. Two strong storms passed through, including strong winds and tennis-ball-sized hail. Several emergence traps placed in both pile types were displaced or damaged from these storms, but were repaired and replaced in less than 24 h. In early November 2013, winter conditions returned with permanent snow cover on 4 December. Most snowfall during winter 2013–2014 occurred in December 2013. Mean winter temperature onsite values ranged from −14.4 °C to 5 °C. Summer temperatures in 2014 were cooler than on average. In both years, the precipitation levels were comparable to long-term averages. Humidity ranged from 69% in May 2013 to 74% in September 2013, and 66% in May 2014 to 77% and 78% in August and September 2014, respectively.
3.2. Pile Characteristics
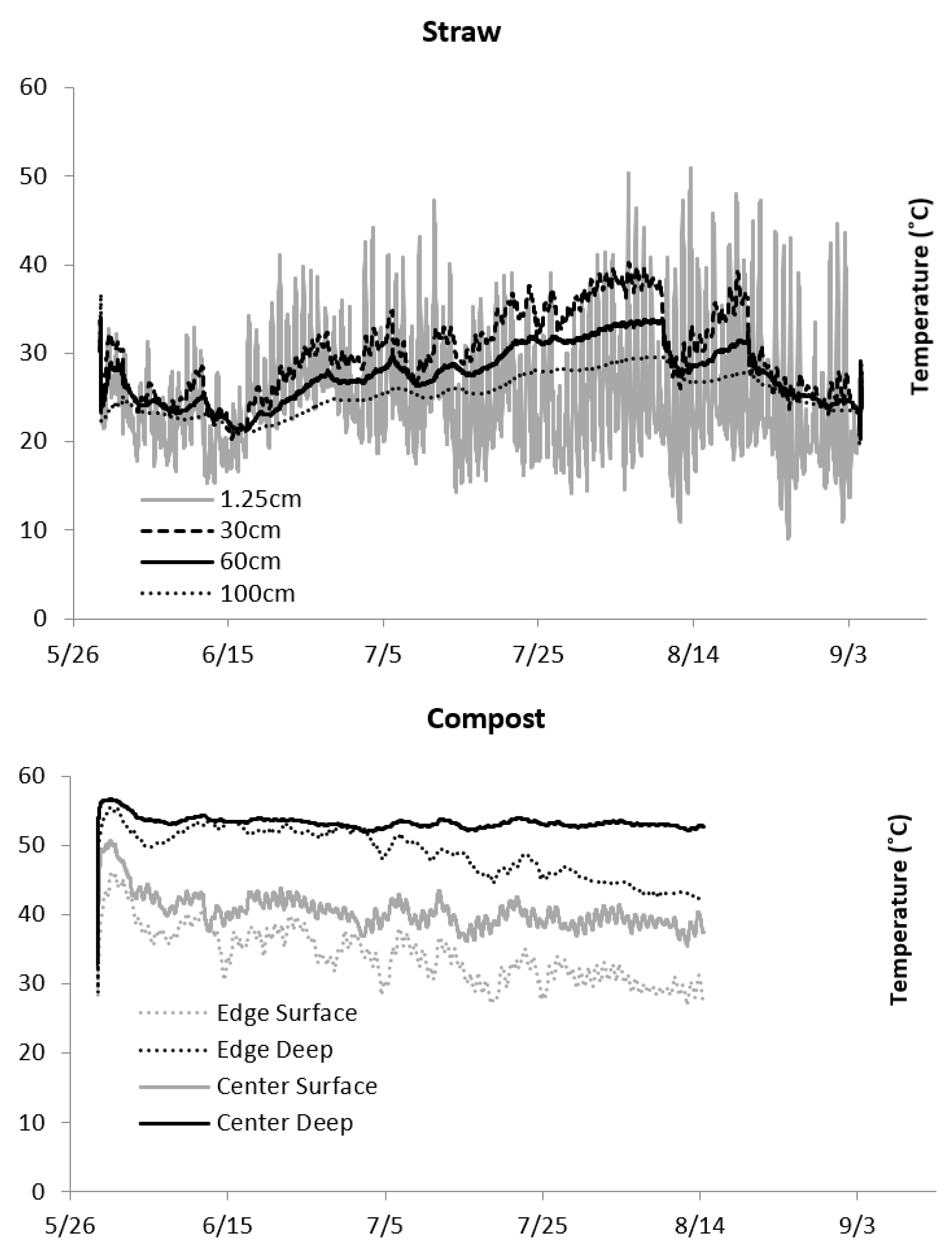
In both years, both straw piles produced flies in September when the emergence traps were dismantled. In contrast, the compost piles were relatively dry. Few maggots were found during troweling and emerging stable fly adults were infrequently captured in the emergence traps. In the straw, the pile surface temperature fluctuated with ambient air temperatures, ranging from 15 to 37 °C. At lower depths, the temperatures were more consistent at 20 °C, but steadily increased above 30 °C as summer progressed (
Figure 3). Compost temperatures remained high throughout summer, ranging from 29 °C on the surface of the pile’s edge up to 53 °C deeper into the pile’s center (
Figure 3). The compost on the surface had more weather exposure, with a greater variance in temperature than the probes buried 30 cm below the surface.
3.3. Ambient Fly Populations
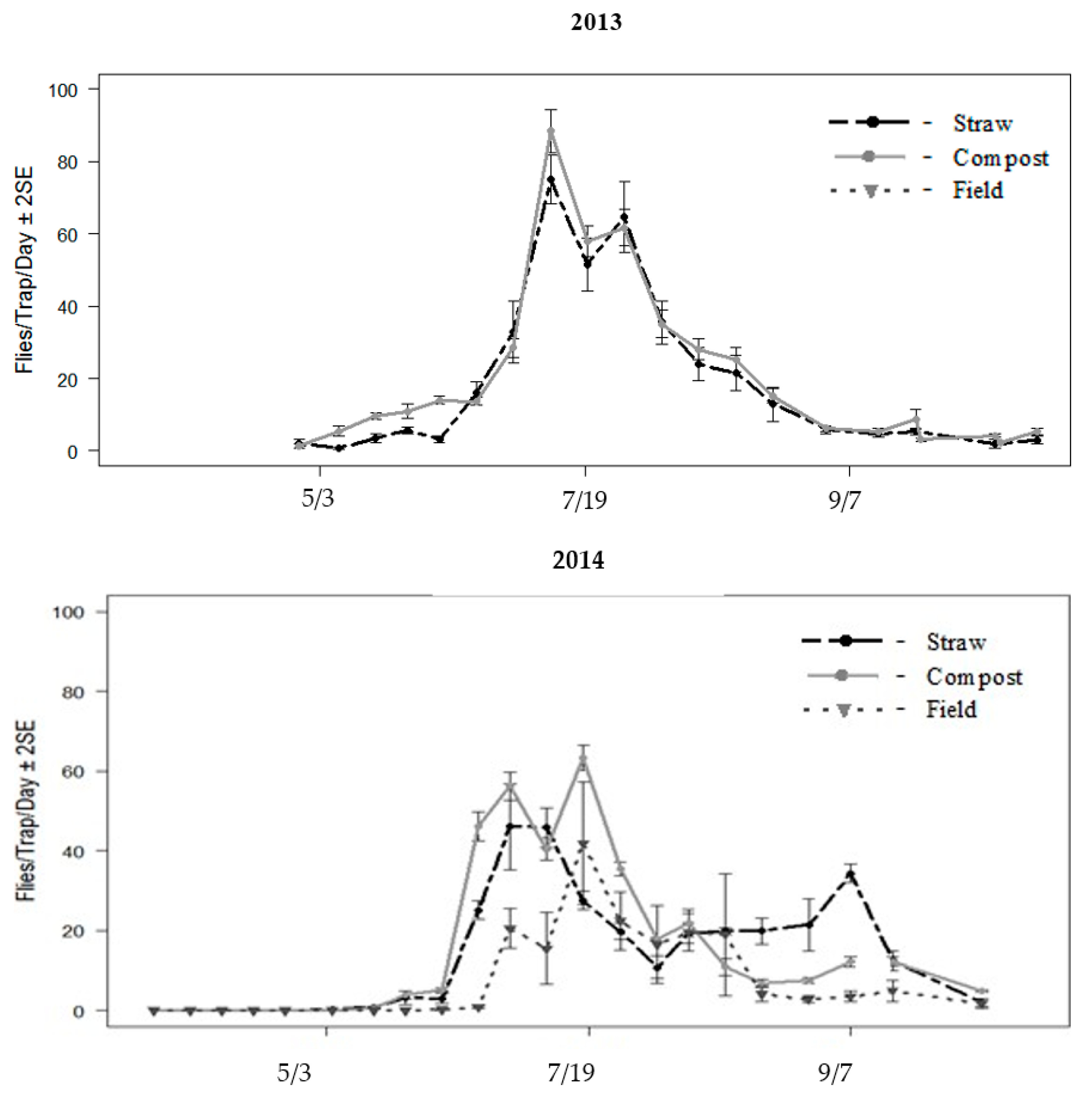
Stable flies became abundant at all locations where Olson traps were set. Flies were already active when the traps were set on 22 May 2013, with an average of 13 flies per trap per day (
Figure 4). The catch rates were lower in the compost than straw for the first five weeks of the study, and then became equal to the straw result after 25 June. The numbers between the compost and straw were similar, although there were significant differences over time. The catch rates peaked in mid-July, with 500 stable flies per trap before the catches tapered off in August.
Traps in 2014 were set before the stable flies became active. The first stable flies were captured in the third week, between 8 to 13 May. No flies were caught in the field traps until mid-June, between 18 and 25 June. Numbers caught in field traps remained low until the first week of July (
Figure 4). The catch rates peaked in the week of 15 July, with 450 flies per straw trap and 300 flies per field trap. The captured flies in compost traps peaked the week before, with 300 flies per trap. All numbers tapered off in July and August. In the week of 25 August, the catch rates in the compost piles increased to 200 flies per trap, while straw traps and those in the field captured 100 flies (
Figure 4). Field trap numbers were consistently lower than traps placed near breeding sources, except for late July through early August when the numbers at all piles were more equal. Sticky sleeves were not changed as frequently in October 2014 as in 2013.
Numbers in the straw traps were higher than the compost at the beginning of both years. More flies were captured throughout 2013 but the catch rates in September and October 2014 were higher than the months in 2013. All Olson traps were dismantled in mid-October in both years when the fly numbers were declining, but still active. In both years, the random effects of the pile and trap in pile accounted for less than 15% of the total variation and were consequently removed from the model. There was a significant interaction between the treatment and time in both years, indicating that different numbers of flies were captured in the pile types with differing patterns during the summer (
Figure 4). These differences were most apparent at the beginning and end of the season.
The random effects of pile and trap in the pile accounted for less than 15% of the total variation in head widths, and so these effects were removed, and analyses proceeded with the fixed effects only. There was no significant interaction between treatment and week (
p = 0.42), and so the interaction was also removed from the model. Flies significantly differed in size over time (
p < 0.001). The head widths of flies sampled at the end of June from both straw and compost piles averaged 2.35 mm in late June and decreased to 2.22 mm in August (
Figure 5). Mean head widths ranged between 2.46 and 2.22 mm (
Figure 5), with significant differences between treatments (
p = 0.001). However, when analyzing a subset of data with head widths of flies sampled from only the compost and straw piles, there were no significant differences in size (
p = 0.62). Flies sampled from field traps were larger than flies sampled from straw or compost piles (
Figure 5).
Of females dissected through summer, over 60% were previtellogenic, indicating recent emergence (
Figure 6). More vitellogenic females were captured in field traps than straw or compost piles.
3.4. Stable Flies Emerging from Piles
In 2013, 120 emergence traps were assembled on leftover debris piles for 12 weeks on straw (14 June to 6 September) and 10 weeks on compost (14 June to 22 August). In 2014, 90 traps were set for 16 weeks on straw (13 May to 4 September) and 11 weeks on compost (29 May to 14 August). Emergence did not cease in the straw piles in early September, but only 2 flies per pile were being captured when the traps were dismantled. Stable flies were last captured in compost traps between 2 and 9 August in 2013 and between 10 and 17 July in 2014.
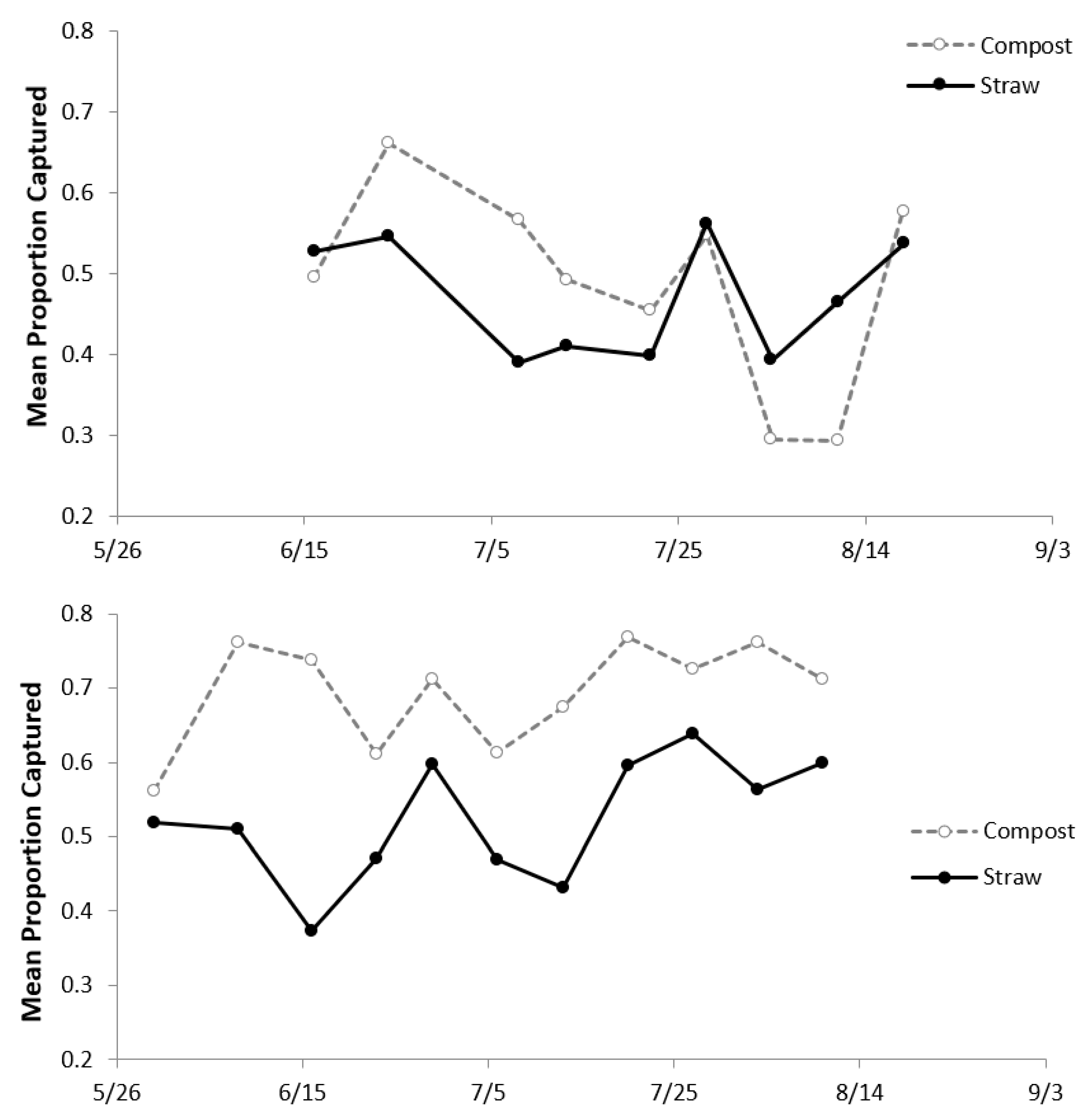
Marker flies were not present in the last week of June in 2013, and so were not set in the emergence traps (
Figure 7). Trap efficiency was highly variable in both types of piles, with the proportion captured ranging from 0.39–0.56 in straw piles and 0.30–0.66 in compost piles. In summer 2013, many marker flies either died inside the vials or failed to emerge due to water accumulation in the vials. To eliminate these problems, each vial was filled with sand in 2014. After adding the sand, virtually no marker flies died inside the vials. The mean proportion of marker flies captured in the compost piles presented a notable increase from 0.48 in 2013 to 0.69 in 2014. In 2014, the proportion captured ranged from 0.37 to 0.64 in the straw piles and 0.56 to 0.77 in compost piles.
The analyses of marker flies were conducted using a subset of data when the traps were assembled in both the straw and compost piles in order to compare the catch rates between the two treatments. Random effects accounted for <15% of the total variation in both years, so these effects were removed from the model and the analyses proceeded with the fixed effects only. The analyses began with a full model, including treatment, time, and interaction. In 2013, there was no interaction between the treatment and time (p = 0.96), and so the interaction was removed from the model. There was no significant difference in the proportion of marker flies captured in the straw and compost piles (p = 0.8) nor any differences in the proportion captured over time (p = 0.65). On average, the trap efficiency was 48%.
As in 2013, there was no significant interaction between the treatment and time in 2014 (
p = 0.24). Consequently, this interaction was removed from the model. The proportion of marker flies captured differed significantly between the straw and compost piles (
p < 0.001). Only 52% of the marker flies were captured in the collection containers of the emergence traps set in the straw piles, and 69% were captured in collection containers in compost piles. There was a significant change in the proportion captured over time (
p = 0.03). More marker flies were captured at both pile types at the end of the season than the beginning (
Figure 7).
Stable fly counts were adjusted based on the results of the marker fly catch rates. Counts were adjusted upwards by dividing the stable flies captured by the proportion of marker flies captured. In 2013, there was no significant interaction between the treatment and time in the proportion of marker flies captured, and the same proportion, 0.48, was used to adjust the stable fly counts. However, in 2014, there was a significant interaction between the treatment and time, and so the proportion used to adjust the stable fly numbers was calculated separately for each week. All the results regarding stable fly emergence numbers from the piles are adjusted catch rates from this analysis.
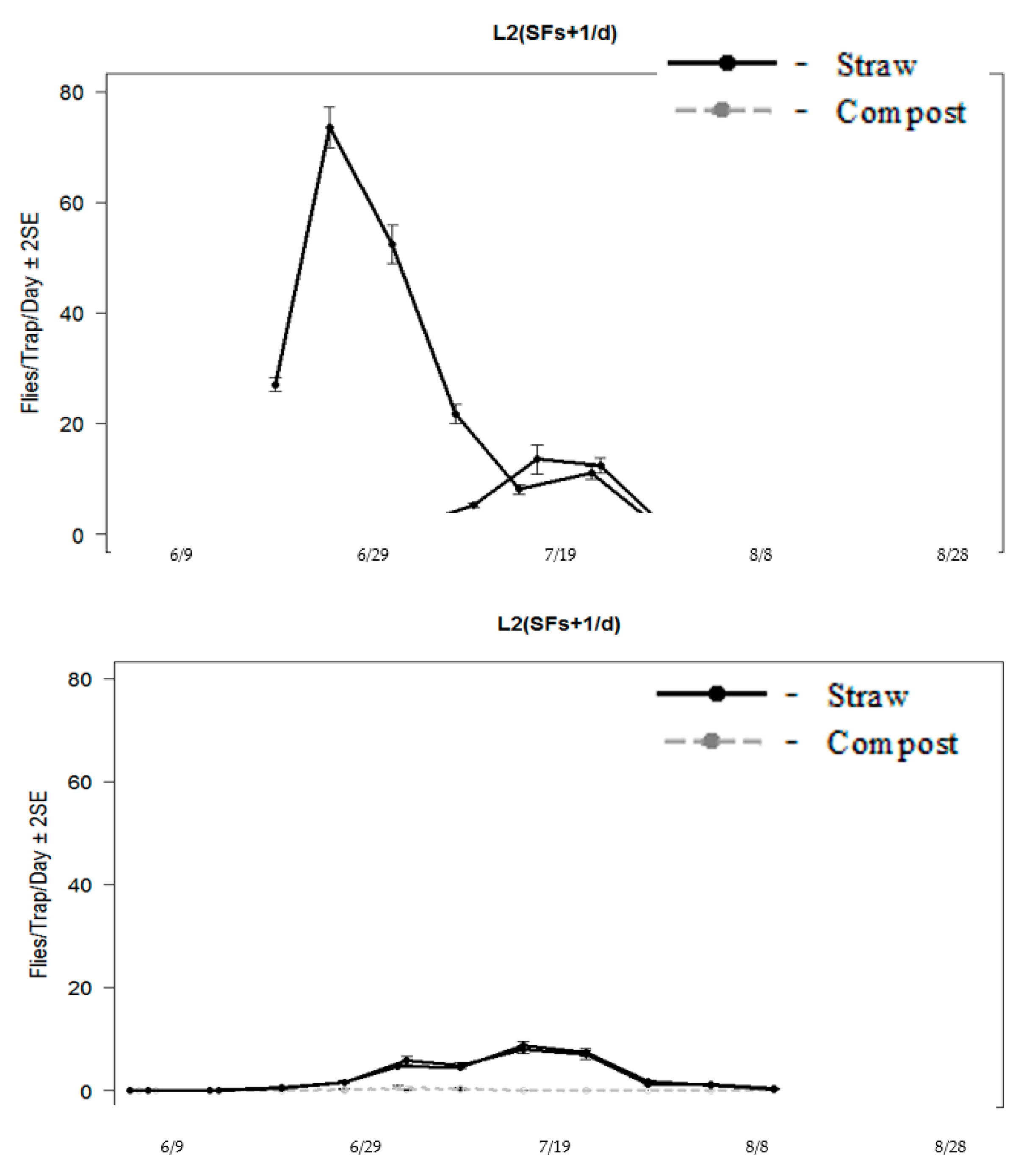
Stable flies were already emerging when the emergence traps were placed in 2013, with a peak emergence at Straw 1 in the week of 22 June (
Figure 8). Weekly catches were used to calculate the mean daily catch rate per emergence trap. Both straw piles produced higher numbers of stable flies than compost barns from late June to early August. Straw 1 was highly productive from mid-June to mid-July, with mean daily catch rates of 20 to 70 flies per trap (
Figure 8). Straw 2 did not peak until the week of 21 July, with approximately 11 flies per day. Compost piles were much lower in production, averaging approximately 1 to 2 flies per day from 26 June to 4 July. In the week of 4 August, the mean daily catch rate in the compost peaked again, averaging 2.5 flies. Stable flies that were captured from the compost piles were typically from traps set at the front of the barn. In this area, the substrate was cooler and wetter due to accumulated rainwater. However, the compost barn area was quite small, and it was difficult to determine how much weather conditions actually affected the fly production numbers. The conditions could also be related to lighting conditions as well as environmental ones. There are many factors that can influence fly production rates. The front part of the compost barn was only 60 cm, which could have influenced fly production, which was quite a small area.
In 2014, no flies were captured in the emergence traps until the week of 13 June (
Figure 8). Both straw piles were very comparable for emergence rates, peaking in the week of 10 July, with approximately 10 flies captured per trap per day and decreasing through August (
Figure 8). The daily catch rates in the compost piles were slightly lower than in 2013, with fewer than 1 fly captured per day consistently through summer.
No significant differences in emergence values were found between the edge and center traps in either pile type when the sampling was stratified in 2013 (p = 0.07), and so the methods were simplified to unrestricted random sampling in 2014. When comparing stable fly emergence rates between the compost and straw piles in 2013, the random effect of the pile accounted for 22% of the total variation, so this effect was retained in the model. There was a significant interaction between the treatment and time, indicating that the emergence patterns over time differed between the two treatments. In 2014, the random effects of the pile and the trap with the pile were removed from the model after accounting for <1% of the total variation. Again, in 2014, there was no significant interaction between the treatment and time.
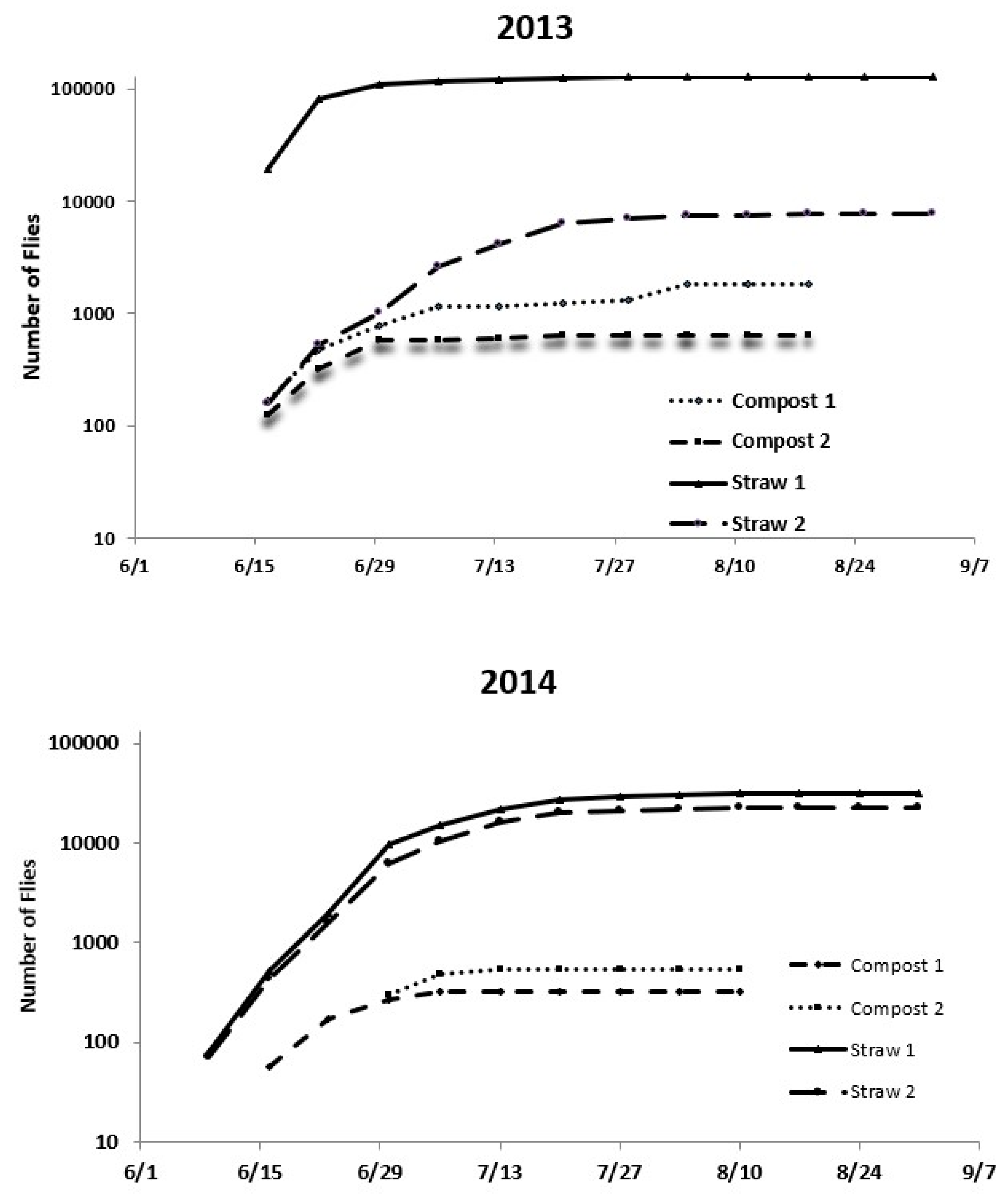
Cumulative emergence numbers from the total straw and compost piles in both years were predicted using extrapolated numbers calculated from the daily catch rates for the emergence traps (
Figure 9). Based on the daily catch rates, Straw 1 alone produced over 120,000 stable flies in 2013 (
Figure 9). Seasonal totals averaged approximately 69,000 flies per straw pile, about 46-times more than the average compost pile for the same number of cow months the previous winter. Although the emergence rate was overall lower in 2014 (
Figure 8), there was a considerable difference in cumulative emergence values between the compost and straw piles. Seasonal totals averaged approximately 45,000 flies per straw pile, over 70-times more than the average compost pile (
Figure 9).
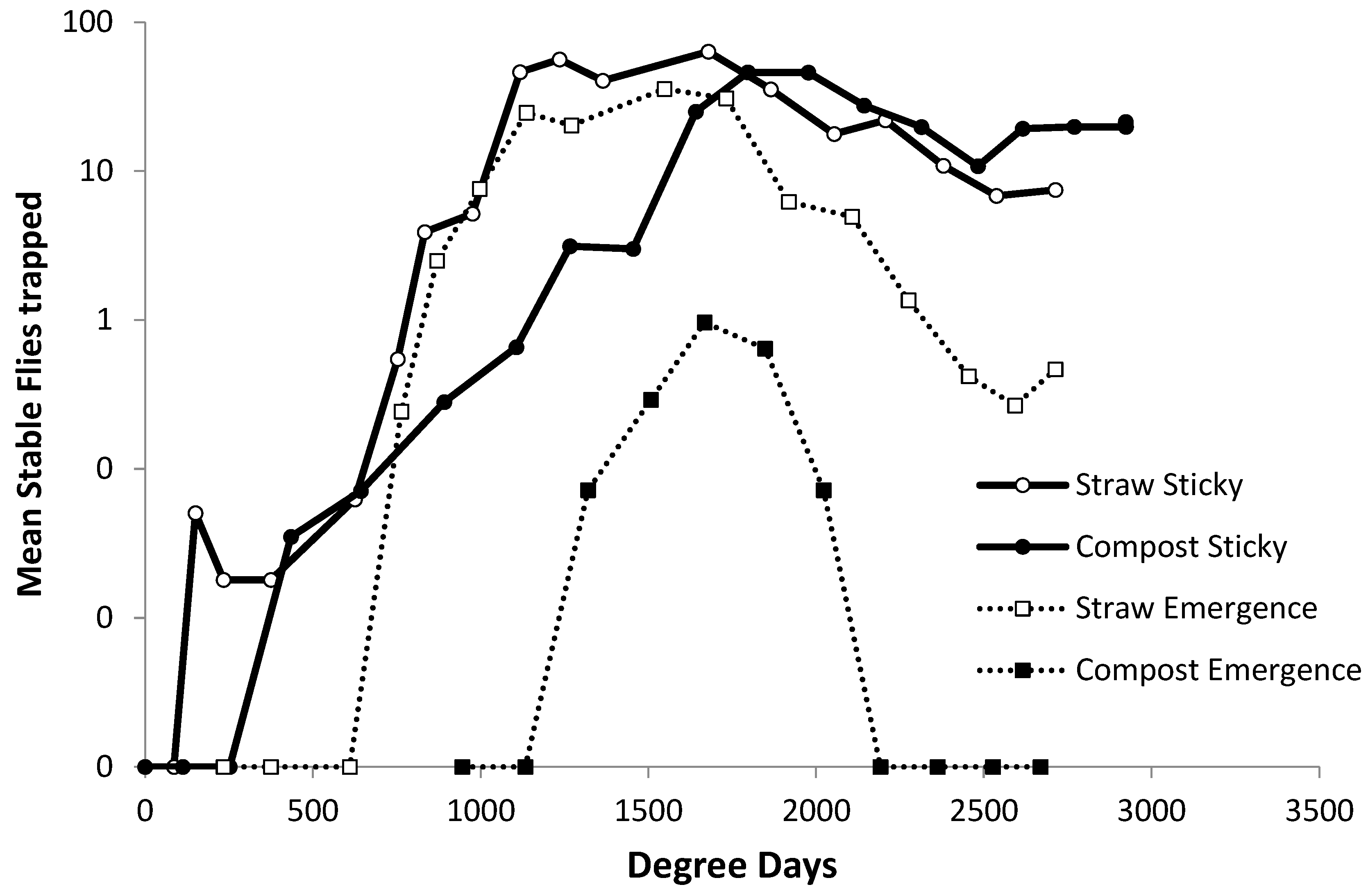
To compare the seasonal pattern of abundance in ambient populations of adults and local emergence from the debris piles, the catch rates from Olson traps in summer 2014 were superposed onto catch rates from emergence traps on a degree-day scale (
Figure 10). The first stable flies were captured in Olson traps placed around straw piles in the second week of May, but no flies were captured in emergence traps on debris piles until the second week of June, approximately one month after initial stable fly activity. Approximately 600 degree days passed between the first flies captured in the Olson traps and the first flies captured in the emergence traps set on straw piles. On compost, the first flies were captured in emergence traps 885 degree days after the first flies were captured in the Olson traps.
4. Discussion
Ambient populations of adult stable flies were overall higher in 2013 than in 2014, though at different times. Olson trap catch rates in October 2014 were higher than the catch rates in October 2013. When left unattended, sleeves can become clogged with insects, dust, and other debris, and lose effectiveness when trapping stable flies. It was possible that the flies declined at a similar rate in both years, but the sleeves were not changed frequently enough to observe the decrease. The decrease in the head widths of the flies sampled from Olson traps could indicate that the substrate quality of developing maggots was not as good at the end of summer as at the beginning of summer. Extrapolating trap efficiency between fly species assumed that the flies would display similar behaviors in the traps [
41], which may not always be the case.
Stable flies prefer to remain in the vicinity of hosts [
42], but are strong fliers capable of flying several kilometers [
43]. Predictably, when cows are moved, stable flies will follow. Semelbauer et al. [
44] reported that stable fly counts were the highest when there was a higher concentration of cattle, as well as more light conditions.
In 2014, the numbers of flies captured on Olson traps decreased through July and increased again in August, a pattern not observed for the Olson traps set near straw or in the field, nor in the catch rates in 2013. In early August, just prior to the observed increase, the conventional dairy herd was transferred from their summer dry lot back to a deep bedded compost barn adjacent to the study barn. With a herd of cows < 10 m away from the set Olson traps, an increased catch of flies was predicted. Therefore, it was possible that the flies were not equally attracted to all piles and Olson trap catches appeared similar between the piles because the cows were housed close to compost debris. In future studies, ensuring that no cows are housed close to any leftover debris can help clarify if flies are equally attracted to both pile types for oviposition. Had no cows been present at the compost site, it is possible that the Olson trap counts would have been lower.
It is possible for developing stable flies to overwinter in piled silage or manure [
45] as a pile’s internal temperature gradient can protect immatures from freezing [
46]. Furthermore, stable fly development was greater in hay mixed with manure, which was similar to the straw and manure in the current study [
33] To ensure that previous piles were not an overwintering source for stable flies, all winter bedding packs were hauled away in fall and new packs were laid. As such, any flies emerging from the piles had to have developed from eggs laid that spring by ambient females. Significantly higher numbers of flies emerging from straw than compost piles were observed, yet the ambient adult density value was similar between types of piles. These results indicate that all piles are equally attractive to adult flies. In 2014, flies captured in Olson traps at the beginning and end of the season were flies from elsewhere that were drawn to the piles. The first stable flies in 2014 occurred on Olson traps approximately one month before any flies were captured in the emergence traps on debris piles, which was consistent with the stable fly development time. However, the number of degree days that passed after the flies were first captured in Olson traps was approximately two-times longer than the generation definition of 327.1 degree days. These results imply that, if the first flies on site oviposit on debris piles, maggots take longer than expected to develop. However, females were either not ovipositing in compost piles or maggots did not survive to reach adulthood. As summer progressed, the catch rates in Olson traps remained higher as emergence trap rates decreased. The differences in the seasonal patterns of the catch rates between sticky and emergence traps imply that stable flies have other breeding sites. Decreasing emergence rates from straw toward the end of summer indicated that straw piles became less suitable for developing maggots as summer progressed.
Differences in emergence rates between straw piles were especially apparent at the beginning of summer 2013. Heavy rain and muddy conditions necessitated the removal of cows from Straw 2, one month before the cows from Straw 1. As a result, Straw 1 had an additional months’ worth of accumulated soiled debris, which could have increased pile attractiveness to ovipositing females. Additionally, cows were still being housed in Straw 1 when stable flies first became active. Flies could have initially been drawn to the pile in search of hosts and aggregated due to the proximity of hosts, mates, and a suitable oviposition site. To ensure the equal treatment of piles, all cows were removed from both straw piles at the same time the following year. Emergence rates between straw piles were much more comparable in summer 2014 and similar to the emergence rates from Straw 2 in 2013, supporting the hypotheses previously described.
At the start of 25 June 2013, a herd of heifers were housed in a concrete loafing area outside the compost barn, while emergence traps were set with a fence blocking the entrance into the barn. However, on 1 July, the herd broke into the compost barn during the night and was not discovered until the next morning. All the emergence traps were destroyed, and significant amounts of fresh manure were added to both piles. Then, in the first week of August, more stable flies were captured than what was trapped in the previous month. The average development time from egg to adult is approximately one month, indicating that females oviposit shortly after the break in, and maggots are able to survive to adulthood. No cows breached the compost barns when the emergence traps were assembled in 2014, and there was no increase in the catch rate in August 2014 as seen in 2013. As such, the addition of fresh urine and manure onto the compost at that time either supported maggots to survive into adulthood or attracted additional females to oviposit. Based on the factors described above, the weather environmental conditions and loss of data from the emergence traps may have contributed to the imperfect experimental data.
Flies became abundant at all trap locations as summer progressed. Catch rates in the field traps in 2014 were lower than compost or straw locations until the first week of July. Fly numbers decreased when the Olson traps were dismantled in both years, however the flies were still active. The catch rates differed significantly over time between locations in both years, despite similar numbers in mid-summer. Flies were less abundant in compost piles than straw piles at the beginning and end of the trapping sessions. Flies trapped at the straw and compost piles were not different in size, whereas flies captured in field traps were significantly larger. The emergence rates were significantly higher in straw packs than compost packs between June and August in both years. In the straw piles, the emergence rate peaked in late June to early July, which was similar to the results found by Taylor [
47], who indicated stable fly populations increased during June and early July in Nebraska. However, the current study reported a decreased stable fly population from August to September. Stable fly catches per trap in compost piles were consistently close to zero during both summers. The head widths of flies emerging from straw piles were larger than flies emerging from compost piles, but these differences were not significant. When catches from the Olson traps were superposed onto catches from emergence traps, there was a difference of approximately 1 month, or degree days between first activity in Olson traps and catches in emergence traps, consistent with the stable fly development time.
The overall differences in the emergence rates between straw and compost locations likely occurred due to several factors. Our results are comparable to the results from Schmidtmann [
8], which demonstrate that straw bedding in calf hutches produces significantly more stable flies than wood chip bedding, which can be attributed to the microbial content, temperature, and moisture content. Little research has been conducted on stable fly oviposition behavior, including the microbial composition in larval development substrates [
34], and maggot development is dependent on live bacterial colonies in the substrate [
33]. Compost piles in Morris were consistently above 40 °C below the surface, temperatures too hot to support developing maggots. In a recent study exploring stable flies and weather conditions, the researchers reported that high temperatures ≤ 30 °C correlated with higher stable fly trap counts [
48]. Furthermore, dairies with feed rations that contained straw had cows with lower fly counts compared to dairies that did not provide straw in rations [
48], which was similar to the current study that found straw bedding had more total flies compared to composted wood shaving bedding.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









