Prey-Related Asphyxiation in Harbor Porpoises (Phocoena phocoena) along the U.S. West Coast: Importance of American Shad (Alosa sapidissima) on Adult Female Harbor Porpoise Mortality


Abstract
:1. Introduction
2. Materials and Methods
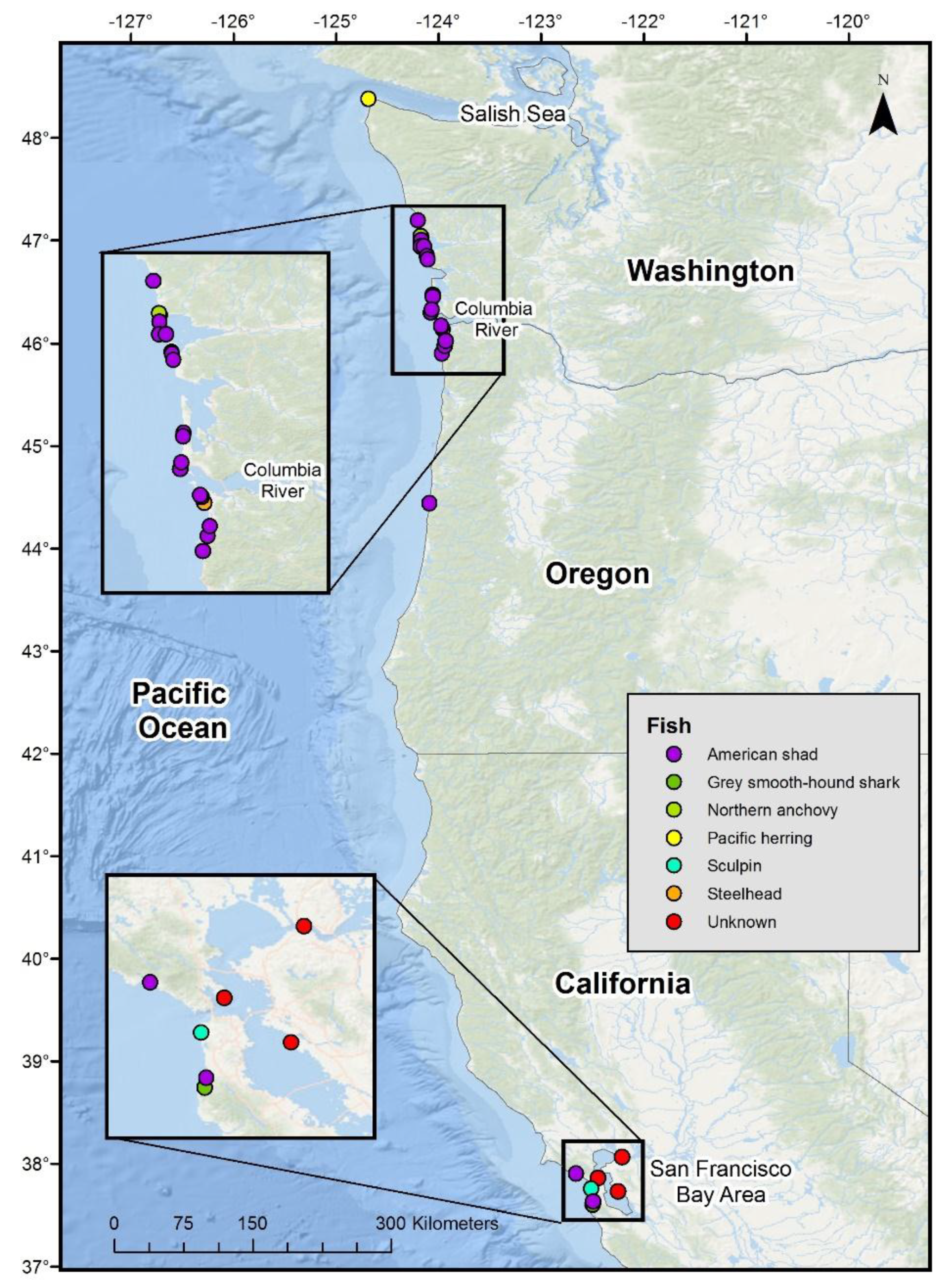
3. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Date (Year/Month/Day) | State | Contributor | HP Sex | HP Age | Length (cm) | Carcass Condition | ♀ Reproductive Status | Fish Species | Fish Length (cm) | Fish Weight (g) | Details |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | 1937/3/9 | CA | 28 * | F | SubA | 101.6 | Fresh | Unknown | Grey smooth-hound shark | 55.9 | NA | Tail of the shark was sticking out of mouth, head was partly in stomach. Froth in lung indicated suffocation. |
| B | 1948 | WA | 30 * | F | A | Unknown | Unknown | Unknown | American shad | 37.5 | NA | American shad stuck head first in throat. |
| C | 1952/6/8 | WA | 31 * | F | A | 147 | Fresh | Not visibly pregnant | American shad | 34.2 | 680 | American shad stuck head first in throat. |
| D | 1979/12/19 | WA | 29 * | F | A | 137 | Fresh | Pregnant | Pacific herring | 44.5 | NA | Fish firmly stuck in throat, pregnant with 20.3 cm well developed fetus. |
| E | 2008/6/30 | CA | 32 * | M | SubA | 132 | Fresh | NA | Sculpin sp. | ~13 | NA | Sculpin lodged in nasal passages, completely obstructing it. |
| 1 | 1983/5/9 | WA | WDFW | F | A | 150 | Moderate | Unknown | American shad | NA | NA | American shad stuck in throat with tail sticking out of the mouth. |
| 2 | 1992/9/5 | CA | CAS/MVZ | F | A | 153 | Moderate | Unknown | American shad | NA | NA | American shad stuck in throat with tail sticking out of the mouth; A pilchard and several squid beaks in stomach. |
| 3 | 2005/6/19 | CA | TMMC | F | A | 165 | Fair | Recently pregnant, endometrial hemosiderosis | American shad | 41 | NA | American shad wedged in esophagus, tail was sticking out of the mouth, goosebeak was dislocated. |
| 4 | 2006/1/12 | WA | CRC | F | A | 161 | Fresh | Pregnant | American shad | NA | NA | American shad stuck in throat head first, goosebeak dislocated. |
| 5 | 2007/5/5 | WA | CRC | U | A | 150 (est.) | Fresh | Unknown | Unknown large fish | NA | NA | Large fish was lodged in mouth, only the tail was visible. Reported by reliable source, carcass not collected. |
| 6 | 2008/5/21 | OR | PSU | F | A | 162.5 | Moderate | Pregnant | American shad | 47 | 943 | American shad stuck head first in throat, goosebeak dislocated. |
| 7 | 2009/3/6 | CA | CAS | M | SubA | 130 | Moderate | NA | Unknown large fish | 35 | NA | Fish lodged in esophagus with tail extending into the oral cavity, goosebeak dislocated and esophagus was distended. |
| 8 | 2009/3/14 | OR | PSU | F | A | 151.4 | Moderate | Pregnant | American shad | 43 | 852 | American shad stuck head first in throat, goosebeak dislocated. |
| 9 | 2010/6/1 | CA | CSP | U | SubA | 122 | Fresh | Unknown | Unknown large fish | NA | NA | Fish stuck in throat, carcass not collected. |
| 10 | 2011/9/13 | WA | PSU | M | SubA | 121.7 | Moderate | NA | American shad | 33 | 398 | American shad stuck head first in throat, goosebeak dislocated, fishery interaction. |
| 11 | 2013/4/29 | OR | OSU | F | A | 163 | Fresh | Pregnant | American shad | 38 | NA | American shad stuck in throat head first obstructing the trachea and bruising the goosebeak. |
| 12 | 2014/2/1 | WA | PSU | F | A | 162.5 | Moderate | Pregnant | American shad | 42.5 | 755 | American shad stuck head first in throat, goosebeak dislocated. |
| 13 | 2014/3/20 | WA | CRC | F | A | 162 | Fresh | Pregnant | American shad | 35.5 | NA | American shad stuck head first in throat, goosebeak dislocated. Head of fish was down level with the lungs, some regurgitated food in the esophagus. Squid beaks and fresh anchovy in stomach. Figure 2. |
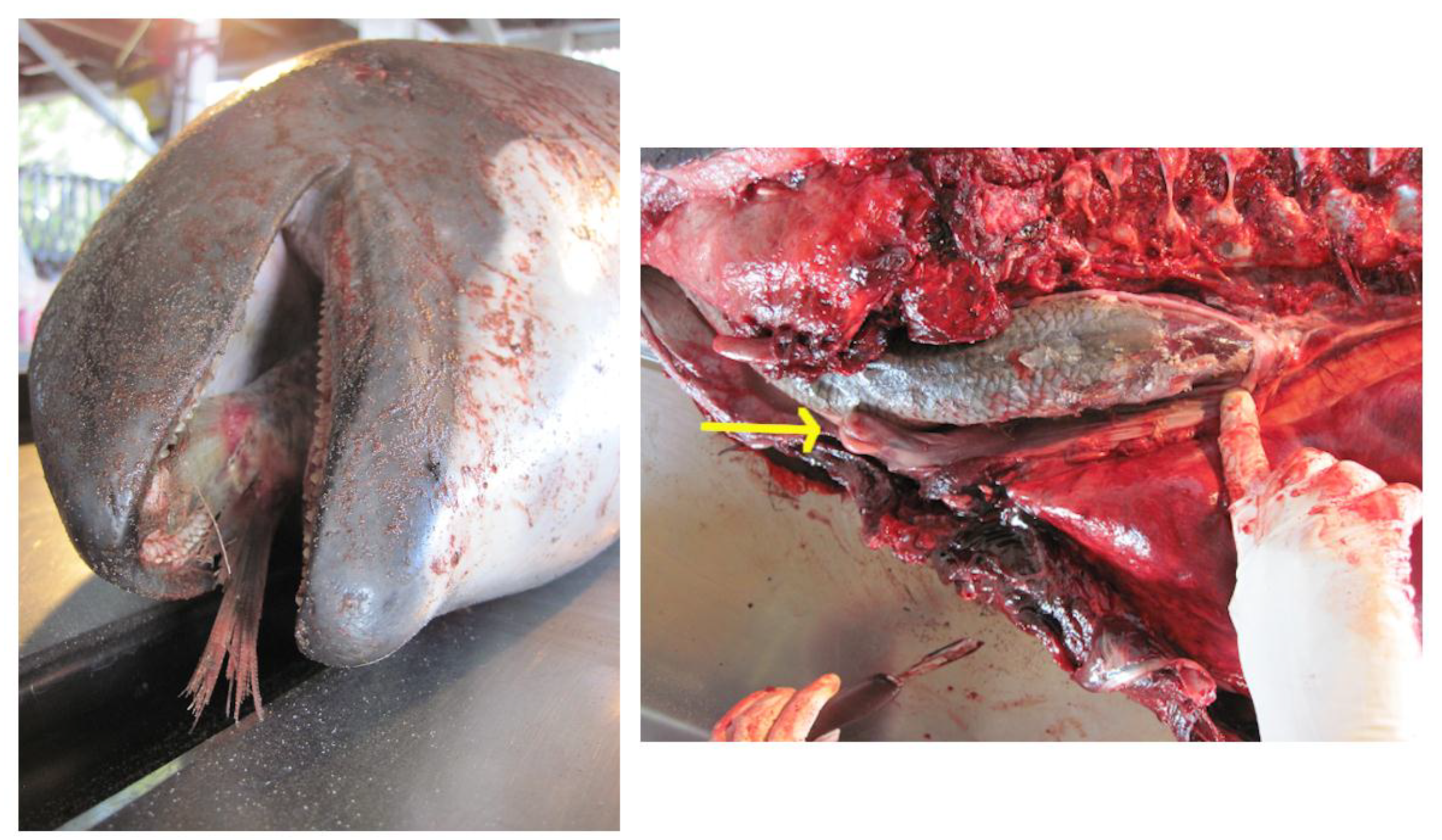
| 14 | 2014/5/9 | WA | CRC | F | A | 181 | Moderate/Advanced | Not pregnant | American shad | 40 * | NA | American shad stuck head first in throat, goosebeak dislocated. Minimal exam due to carcass condition. * scavenged, estimated length. Figure 3. |
| 15 | 2016/5/23 | OR | PSU | F | A | 168 | Moderate | Pregnant | American shad | 46.7 | 870 | American shad stuck head first in throat, goosebeak dislocated. Fishery interaction. |
| 16 | 2016/7/3 | OR | PSU | F | A | 163 | Fresh | Pregnant | Steelhead | 70 | 3200 | Steelhead stuck head first in throat, goosebeak dislocated. Fishery interaction. A full-term fetus was pressing on the caudal end of the forestomach, preventing full expansion of the stomach by 2.5 to 5 cm and thereby preventing the fish from being completely swallowed. |
| 17 | 2016/7/16 | OR | PSU | F | A | 169.2 | Advanced | Recently pregnant, distended uterus | American shad | 49 | 837 | American shad stuck head first in throat, goosebeak dislocated. Fishery interaction. Recently pregnant with distended uterus. |
| 18 | 2017/4/28 | WA | PSU | F | A | 174.5 | Moderate | Pregnant | American shad | 30.9 | 870 | 3/4 of American shad stuck head first in throat, goosebeak dislocated. Fishery interaction; 649 g intact American shad also in forestomach. |
| 19 | 2018/5/31 | WA | PSU | F | A | 166 | Fresh | Pregnant | American shad | 41.1 | 1269 | American shad stuck head first in throat, goosebeak dislocated. |
| 20 | 2018/8/15 | CA | CAS | F | A | 152.5 | Advanced | Unknown | Unknown large fish | ~30 | NA | Fish stuck head first in throat, tail sticking out of the mouth, goosebeak dislocated. |
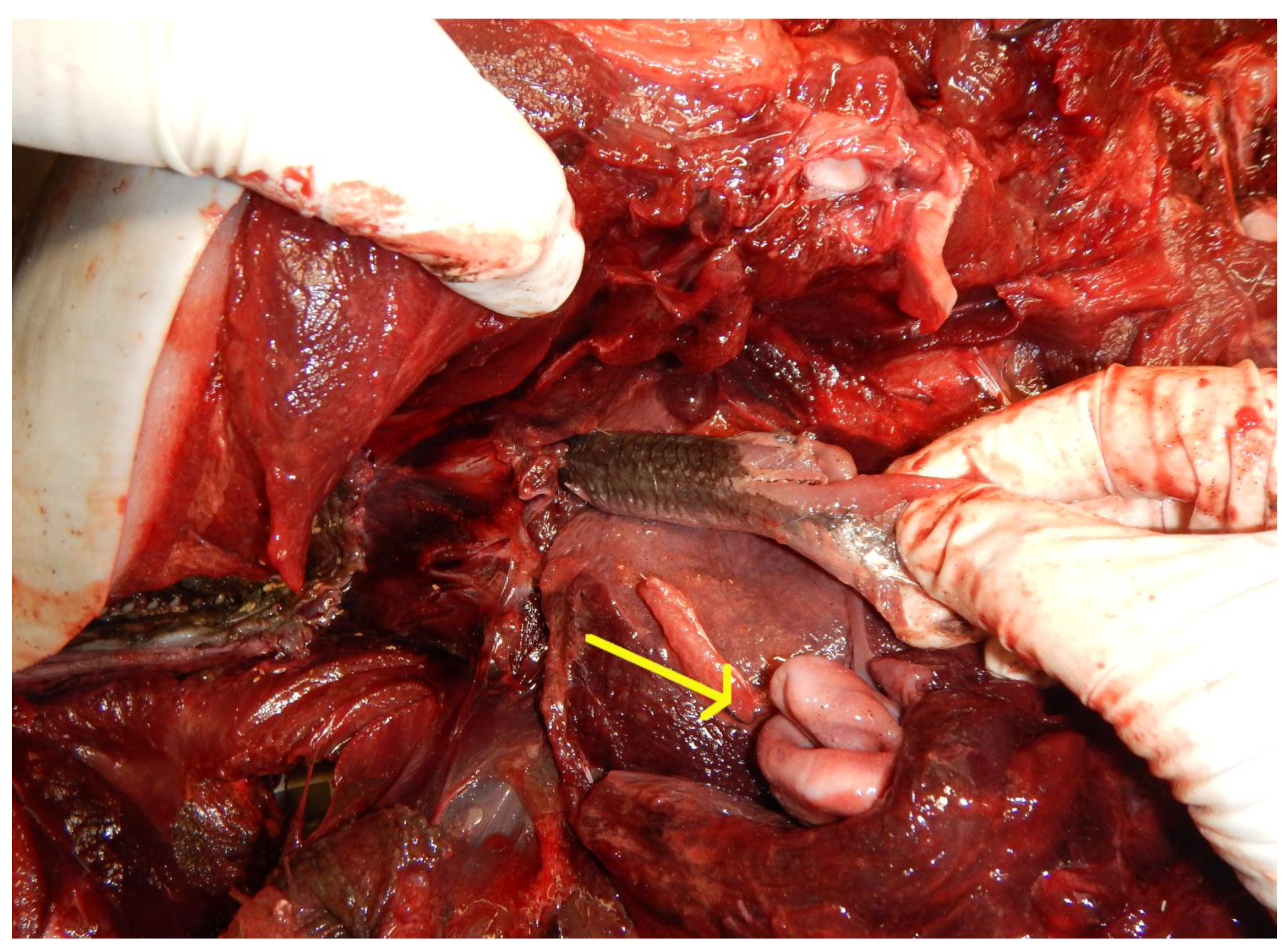
| 21 | 2018/10/3 | WA | CRC | F | A | 184 | Fresh | Unable to determine | Northern anchovy | 7.6 | NA | Fish lodged in blowhole. It was suspected that the harbor porpoise was feeding on the anchovy when attacked. The goosebeak was dislocated and there was bruising and edema around the blowhole at the top of the head and bruising in the back of the throat (Figure 4). There were pre-mortem bite marks from a broadnose seven gill shark. Reproductive status of the harbor porpoises was unknown because the shark had bitten out that area of the abdomen. Fresh and partially digested anchovy in stomach. |
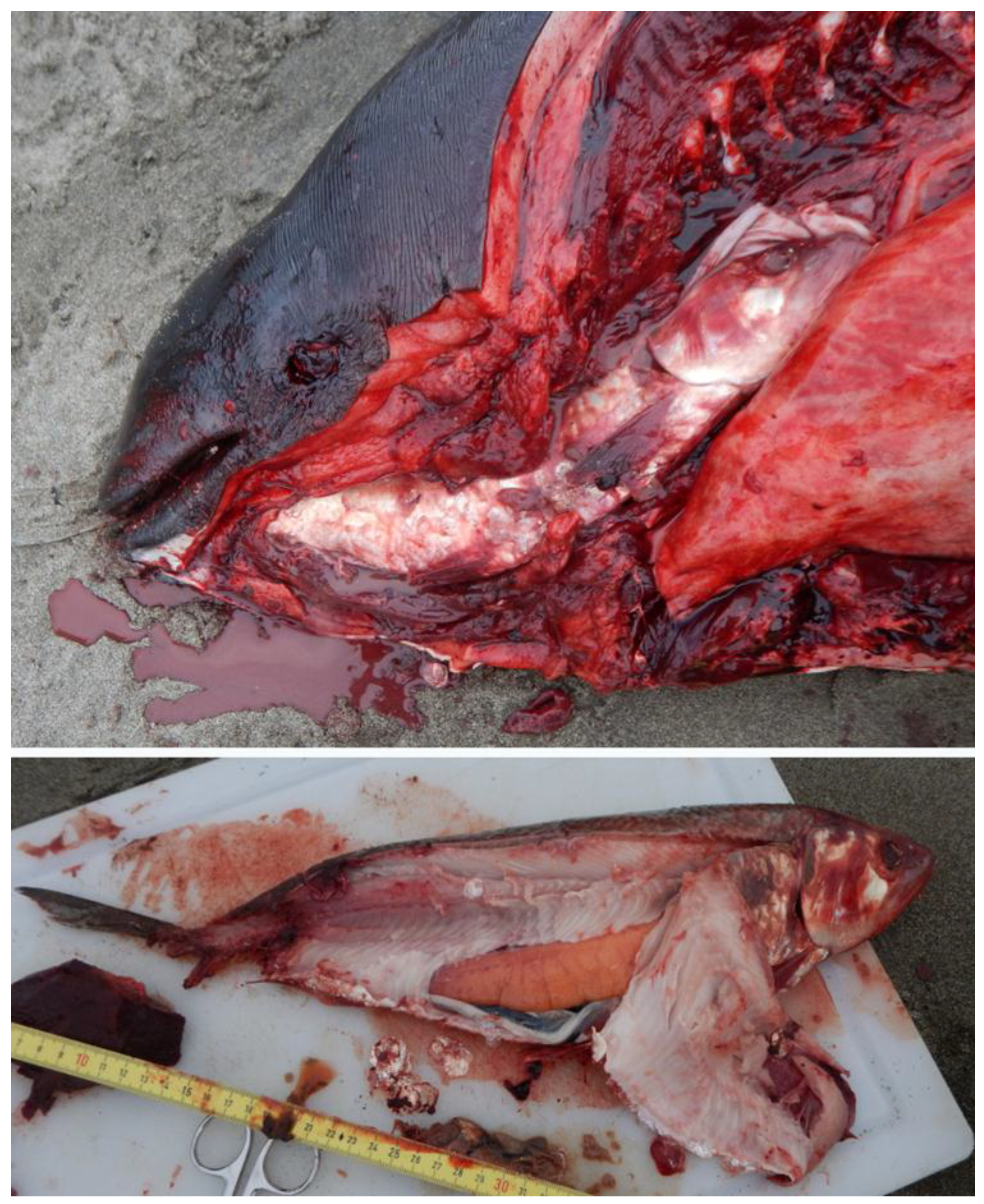
| 22 | 2019/6/28 | WA | CRC | F | A | 174.5 | Fresh | Possibly post-partum | American shad | 49 | NA | American shad stuck head first in throat, goosebeak dislocated and the fish was down the esophagus past the heart (Figure 5). Shad was a gravid female. |
| 23 | 2019/9/19 | OR | PSU | F | A | 167.5 | Moderate | Not pregnant, recent corpus luteum | American shad | 46.5 | 686 | American shad stuck head first in throat, goosebeak dislocated. Not pregnant or lactating, but had recent corpus luteum. |
| 24 | 2020/4/25 | WA | CRC | F | A | 165 | Fresh | Pregnant | American shad | 49 | NA | American shad (gravid female) stuck head first in throat, goosebeak dislocated (Figure 6). Stomach full of partially digested anchovy. Third-trimester fetus. |
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Osmek, S.; Calambokidis, J.; Laake, J.; Gearin, P.; Delong, R.; Scordino, J.; Jeffries, S.; Brown, R. Assessment of the Status of Harbor Porpoise (Phocoena phocoena) in Oregon and Washington Waters; NOAA Technical Memorandum NMFS-AFSC-76; U.S. Department of Commerce: Washington, DC, USA, 1996.
- Barlow, J. Harbor porpoise, Phocoena phocoena, abundance estimation for California, Oregon, and Washington I. Ship surveys. Fish. Bull. 1988, 83, 417–432. [Google Scholar]
- Forney, K.A.; Carretta, J.V.; Benson, S.R. Preliminary Estimates of Harbor Porpoise Abundance in Pacific Coast Waters of California, Oregon, and Washington, 2007–2012; NOAA Technical Memorandum NMFS-SWFSC-537; U.S. Department of Commerce: Washington, DC, USA, 2014.
- Encyclopedia Britannica. Available online: https://www.britannica.com/science/continental-shelf (accessed on 1 June 2020).
- Nielsen, N.H.; Teilmann, J.; Sveegaard, S.; Hansen, R.G.; Sinding, M.H.S.; Dietz, R.; Heide-Jørgensen, M.P. Oceanic movements, site fidelity and deep diving in harbour porpoises from Greenland show limited similarities to animals from the North Sea. Mar Ecol. Prog. Ser. 2018, 597, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.E. Food habits of smaller marine mammals from northern California. Proc. Calif. Acad. Sci. 1981, 42, 409–433. [Google Scholar]
- Dorfman, E.J. Distribution, Behavior, and Food Habits of Harbor Porpoises (Phocoena phocoena) in Monterey Bay. Master’s Thesis, San Jose State University, San Jose, CA, USA, 1990. [Google Scholar]
- Gearin, P.J.; Melin, S.R.; Delong, R.L.; Kajimura, H.; Johnson, M.A. Harbor porpoise interactions with a chinook salmon set-net fishery in Washington State. Rep. Int. Whal. Commn. Special 1994, 427–438. [Google Scholar]
- Walker, W.A.; Hanson, M.B.; Baird, R.W.; Guenther, T.J. Food habits of the harbor porpoise, Phocoena phocoena, and Dall’s porpoise, Phocoenoides dalli, in the inland waters of British Columbia and Washington. Alask. Fish. Sci. Center Process. Rep. 1998, 98-10, 63–75. [Google Scholar]
- Toperoff, A.K. Examination of Diet of Harbor Porpoise (Phocoena phocoena) from Central California Using Stomach Content and Stable Isotope Analysis from Multiple Tissues. Master’s Thesis, San Jose State University, San Jose, CA, USA, 2002. [Google Scholar]
- Nichol, L.M.; Hall, A.M.; Ellis, G.M.; Stredulinsky, E.; Boogaards, M.; Ford, J.K. Dietary overlap and niche partitioning of sympatric harbour porpoises and Dall’s porpoises in the Salish Sea. Prog. Oceanogr. 2013, 115, 202–210. [Google Scholar] [CrossRef]
- Oliaro, J.F. Identification of Harbor Porpoise (Phocoena phocoena) Prey Species Using PCR Based Molecular Technique. Master’s Thesis, San Francisco University, San Francisco, CA, USA, 2013. [Google Scholar]
- Smith, G.J.D.; Gaskin, D.E. The diet of harbor porpoises (Phocoena phocoena (L.)) in coastal water of Eastern Canada, with special reference to the Bay of Fundy. Can. J. Zool. 1974, 52, 777–782. [Google Scholar] [CrossRef]
- Recchia, C.A.; Read, A.J. Stomach contents of harbour porpoises, Phocoena phocoena (L.), from the Bay of Fundy. Can. J. Zool. 1989, 67, 2140–2146. [Google Scholar] [CrossRef]
- Smith, R.J.; Read, A.J. Consumption of euphausiids by harbour porpoise (Phocoena phocoena) calves in the Bay of Fundy. Can. J. Zool. 1992, 70, 1629–1632. [Google Scholar] [CrossRef]
- Fontaine, P.M.; Hammill, M.O.; Barrette, C.; Kingsley, M.C. Summer diet of the harbour porpoise (Phocoena phocoena) in the estuary and the northern Gulf of St. Lawrence. Can. J. Fish. 1994, 51, 172–178. [Google Scholar] [CrossRef]
- Aarefjord, H.; Bjørge, A.J.; Kinze, C.C.; Lindstedt, I. Diet of the harbour porpoise (Phocoena phocoena) in Scandinavian waters. Oceanogr. Lit. Rev. 1996, 10, 1041. [Google Scholar]
- Martin, A.R. The Diet of Harbour Porpoises (Phocoena phocoena) in British Waters; SC/47/SM48; International Whaling Commission: Cambridge, UK, 1996. [Google Scholar]
- Benke, H.; Siebert, U.; Lick, R.; Bandomir, B.; Weiss, R. The current status of harbor porpoises (Phocoena phocoena) in German waters. Arch. Fish. Mar. Res./Arch. Fisch. Meeresforsch. 1998, 46, 97–123. [Google Scholar]
- Gannon, D.P.; Craddock, J.E.; Read, A.J. Autumn food habits of harbor porpoises, Phocoena phocoena, in the Gulf of Maine. Fish. Bull. 1998, 96, 428–437. [Google Scholar]
- Víkingsson, G.A.; Ólafsdóttir, D.; Sigurjónsson, J. Geographical, and seasonal variation in the diet of harbour porpoises (Phocoena phocoena) in Icelandic coastal waters. NAMMCO Sci. Publ. 2003, 5, 243–270. [Google Scholar] [CrossRef]
- Sveegaard, S.; Andreasen, H.; Mouritsen, K.N.; Jeppesen, J.P.; Teilmann, J.; Kinze, C.C. Correlation between the seasonal distribution of harbour porpoises and their prey in the Sound, Baltic Sea. Mar. Biol. 2012, 159, 1029–1037. [Google Scholar] [CrossRef] [Green Version]
- Wisniewska, D.M.; Johnson, M.; Teilmann, J.; Rojano-Doñate, L.; Shearer, J.; Sveegaard, S.; Madsen, P.T. Ultra-high foraging rates of harbor porpoises make them vulnerable to anthropogenic disturbance. Curr. Biol. 2016, 26, 1441–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreasen, H.; Ross, S.D.; Siebert, U.; Andersen, N.G.; Ronnenberg, K.; Gilles, A. Diet composition and food consumption rate of harbor porpoises (Phocoena phocoena) in the western Baltic Sea. Mar. Mammal Sci. 2017, 33, 1053–1079. [Google Scholar] [CrossRef]
- Elliser, C.R.; Hessing, S.; MacIver, K.H.; Webber, M.A.; Keener, W. Harbor porpoises (Phocoena phocoena vomerina) catching and handling large fish on the U.S. West Coast. Aquat. Mamm. 2020, 46, 191–199. [Google Scholar] [CrossRef]
- D’Alessandro, D.N.; Duffield, D.A. Salmonid passive integrated transponder tags and coded wire tags found in the forestomach of a harbor porpoise (Phocoena phocoena) in southwestern Washington. Fish. Bull. 2019, 117, 303–307. [Google Scholar] [CrossRef]
- Treacy, S.D. Feeding habits of marine mammals from Grays Harbor, Washington to Netarts Bay Oregon. Marine mammals and their interactions with fisheries of the Columbia River and adjacent waters, 1980–1982. NOAA: Northwest Alaska Fish. Sci. Cent. NWAFC Processed Report 85-04. Natl. Mar. Fish. Serv. 1985, 149–198. Available online: https://archive.fisheries.noaa.gov/afsc/Publications/ProcRpt/NWAFCPR85-04.pdf (accessed on 2 May 2020).
- Orr, R.T. A porpoise chokes on a shark. J. Mammal. 1937, 18, 370. [Google Scholar] [CrossRef]
- Hult, R.W.; Dupey, S.E.; Badley, R.W. Mortalities associated with prey ingestion by small cetaceans. Cetology 1980, 38, 1–2. [Google Scholar]
- Scheffer, V.B.; Slipp, J.W. The whales and dolphins of Washington State with a key to the cetaceans of the West Coast of North America. Am. Midl. Nat. 1948, 39, 257–337. [Google Scholar] [CrossRef]
- Scheffer, V.B. Measurements and stomach contents of eleven delphinids from Northeast Pacific. Murrelet 1953, 34, 27–30. [Google Scholar] [CrossRef]
- Wilkin, S.M.; Cordaro, J.; Gulland, F.M.D.; Wheeler, E.; Dunkin, R.; Sigler, T.; Casper, D.; Berman, M.; Flannery, M.; Fire, S.; et al. An unusual mortality event of harbor porpoises (Phocoena phocoena) off central California: Increase in blunt trauma rather than an epizootic. Aquat. Mamm. 2012, 38, 301–310. [Google Scholar] [CrossRef]
- MacLeod, C.D.; Reidenberg, J.S.; Weller, M.; Santos, M.B.; Herman, J.; Goold, J.; Pierce, G.J. Breaking symmetry: The marine environment, prey size, and the evolution of asymmetry in cetacean skulls. Anat. Rec. 2007, 290, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Siebert, U.; Wünschmann, A.; Weiss, R.; Frank, H.; Benke, H.; Frese, K. Post-mortem findings in harbour porpoises (Phocoena phocoena) from the German North and Baltic Seas. J. Comp. Path. 2001, 124, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.; Bolin, V. Stranded harbour porpoise (Phocoena phocoena (Grey)) with throat obstructed by a scad (Trachurus trachurus (L.)). Ir. Nat. J. 2014, 33, 155. [Google Scholar]
- Roller, M.; Gross, S.; Reckendorf, A.; Andreasen, H.; Wohlsein, P.; Siebert, U. Fatal asphyxiation with laryngeal displacement caused by platvissen in harbour porpoises. In Proceedings of the 31st Annual Conference of the European Cetacean Society, Middelfart, Denmark, 1–3 May 2017. [Google Scholar]
- Sveegaard, S. Personal Communication; Aarhus University: Aarhus, Denmark, 2020. [Google Scholar]
- Byard, R.W.; Gilbert, J.D.; Gibbs, S.E.; Kemper, C.M. Cetacean Café Coronary. J. Clin. Forensic Med. 2003, 10, 85–88. [Google Scholar] [CrossRef]
- Byard, R.W.; Tomo, I.; Kemper, C.M.; Gibbs, S.E.; Bossley, M.; Machado, A.; Hill, M. Unusual causes of fatal upper aerodigestive tract obstruction in wild bottlenose dolphins (Tursiops aduncus). Forensic Sci. Med. Pathol. 2010, 6, 207–210. [Google Scholar] [CrossRef]
- Watson, A.; Gee, L.E. Laryngeal displacement and asphyxiation by a beheaded sheepshead (Archosargus probatocephalus) in a bottlenose dolphin (Tursiops truncatus). Aquat. Mamm. 2005, 31, 447–452. [Google Scholar] [CrossRef]
- Mignucci-Giannoni, A.A.; Rosario-Delestre, R.J.; Alsina-Guerrero, M.M.; Falcon-Matos, L.; Guzmán-Ramírez, L.; Williams, E.H.; Bossart, G.D.; Reidenberg, J. Asphyxiation in a bottlenose dolphin (Tursiops truncatus) from Puerto Rico due to choking on a black margate (Anisotremus surinamensis). Aquat. Mamm. 2009, 35, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Stolen, M.; St. Leger, S.; Noke Durden, W.; Mazza, T.; Nilson, E. Fatal asphyxiation in bottlenose dolphins (Tursiops trunactus) from the Indian River Lagoon. PLoS ONE 2013, 8, e66828. [Google Scholar] [CrossRef] [PubMed]
- IJsseldijk, L.L.; Leopold, M.F.; Bravo Rebolledo, E.L.; Deaville, R.; Haelters, J.; IJzer, J.; Jepson, P.D.; Gröne, A. Fatal asphyxiation in two long-finned pilot whales (Globicephala melas) caused by common soles (Solea solea). PLoS ONE 2015, 10, e0141951. [Google Scholar] [CrossRef] [PubMed]
- Mariani, D.B.; Guimarães, J.P.; Batista, R.G.; Brum, A.; Groch, K.R.; Díaz-Delgado, J.; Parente, J.E.V. Fatal asphyxia due to laryngeal displacement by large-sized prey in a Guiana dolphin (Sotalia guianensis), Brazil. Cienc. Rural 2020, 50, e20190068. [Google Scholar] [CrossRef]
- Rouse, N.; Burek-Huntington, K.A.; Shelden, K.E.W. Asphyxiation of an endangered Cook Inlet beluga whale, Delphinapterus leucas. Mar. Fish. Rev. 2017, 79, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Huggins, J.L.; Raverty, S.A.; Norman, S.A.; Calambokidis, J.; Gaydos, J.K.; Duffield, D.A.; Lambourn, D.M.; Rice, J.M.; Hanson, B.; Wilkinson, K.; et al. Increased harbor porpoise mortality in the Pacific Northwest, USA: Understanding when higher levels may be normal. Dis. Aquat. Organ. 2015, 115, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Pearcy, W.G.; Fisher, P. Ocean distribution of the American shad (Alosa sapidissma) along the Pacific coast of North America. Fish. Bull. 2011, 109, 440–453. [Google Scholar]
- Pugliares-Bonner, K.; Bogomolni, A.; Touhey, K.; Herzig, S.; Harry, C.; Moore, M. Marine Mammal Necropsy: An Introductory Guide for Stranding Responders and Field Biologists. Woods Hole Oceanog. Inst. Tech. Rept. 2007. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.; Gero, S.; Bejder, L.; Calambokidis, J.; Kraus, S.D.; Lusseau, D.; Read, A.J.; Robbins, J. Underestimating the damage: Interpreting cetacean carcass recoveries in the context of the Deepwater Horizon/BP incident. Conserv. Lett. 2011, 4, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.J.; Mitchell, G.H.; Rowles, T.K.; Early, G. Dead cetacean? Beach, bloat, float, sink. Front. Mar. Sci. 2020, 7, 333. [Google Scholar] [CrossRef]
- Hasselman, D.J.; Hinrichsen, R.A.; Shields, B.A.; Ebbesmeyer, C.C. The rapid establishment, dispersal and increased abundance of invasive American shad in the Pacific Northwest. Fisheries 2012, 37, 103–114. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Available online: https://www.fws.gov/fisheries/freshwater-fish-of-america/american_shad.html (accessed on 1 June 2020).
- Lodi, L.; Hetzel, B. Rough-toothed dolphin, Steno bredanensis, feeding behavior in Ilha Grande Bay, Brazil. Biociéncias 1999, 7, 29–42. [Google Scholar]
- Best, R.C.; da Silva, V.M.F. Inia geoffrensis de Blainville, 1817. Mamm. Species 1993, 426, 1–8. [Google Scholar]
- Ronje, E.I.; Barry, K.P.; Sinclair, C.; Grace, M.A.; Barros, N.; Allen, J.; Balmer, B.; Panike, A.; Toms, C.; Mullin, K.D.; et al. A common bottlenose dolphin (Tursiops truncatus) prey handling technique for marine catfish (Ariidae) in the northern Gulf of Mexico. PLoS ONE 2017, 12, e0181179. [Google Scholar] [CrossRef] [Green Version]
- Ralls, K.; Mesnick, S.L. Sexual dimorphism. In Encyclopedia of Marine Mammals; Perrin, W.F., Wursig, B., Thewissen, J.G.M., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 1071–1078. [Google Scholar]
- Hohn, A.A.; Brownell, J.R.L., Jr. Harbor Porpoise in Central California Waters: Life History and Incidental Catches; IWC Report SC/42/SM47; International Whaling Commission: Cambridge, UK, 1990. [Google Scholar]
- Norman, S.A.; Hanson, M.B.; Huggins, J.; Lambourn, D.; Calambokidis, J.; Cottrell, P.; Greene, A.; Raverty, S.; Berta, S.; Dubpernell, S.; et al. Conception, fetal growth, and calving seasonality of harbor porpoise (Phocoena phocoena) in the Salish Sea waters of Washington, USA and southern British Columbia, Canada. Can. J. Zool. 2018, 96, 566–575. [Google Scholar] [CrossRef] [Green Version]
- Read, A.J.; Hohn, A.A. Life in the fast lane: The life history of harbor porpoises from the Gulf of Maine. Mar. Mammal Sci. 1995, 11, 423–440. [Google Scholar] [CrossRef]
- Lockyer, C.; Kinze, C. Status, ecology and life history of harbour porpoise (Phocoena phocoena), in Danish waters. NAMMCO Sci. Publ. 2003, 5, 143–175. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, C.A.; Stern, S.J.; Hines, E. The metabolic cost of swimming and reproduction in harbor porpoises (Phocoena phocoena) as predicted by a bioenergetic model. Mar. Mammal Sci. 2018, 34, 875–900. [Google Scholar] [CrossRef]
- Yasui, W.Y.; Gaskin, D.E. Energy budget of a small cetacean, the harbour porpoise, Phocoena phocoena (L.). Ophelia 1986, 25, 183–197. [Google Scholar] [CrossRef]
- Gaskin, D.E.; Watson, A.P. The harbor porpoise, Phocoena phocoena, in Fish Harbour, New Brunswick, Canada: Occupancy, distribution, and movements. Fish. Bull. 1985, 83, 427–442. [Google Scholar]
- Smith, G.J.D.; Gaskin, D.E. An environmental index for habitat utilization by female harbour porpoises with calves near Deer Island, Bay of Fundy. Ophelia 1983, 22, 1–13. [Google Scholar] [CrossRef]
- Christiansen, F.; Víkingsson, G.A.; Rasmussen, M.H.; Lusseau, D. Female body condition affects foetal growth in a capital breeding mysticete. Funct. Ecol. 2014, 28, 579–588. [Google Scholar] [CrossRef]
- Mukherjee, S.; Heithaus, M.R. Dangerous prey and daring predators: A review. Biol. Rev. 2013, 88, 550–563. [Google Scholar] [CrossRef] [PubMed]
- McGovern, B.; Gridley, T.; James, B.S.; Elwen, S. Risky business? A note on repeated live strandings of common bottlenose dolphins (Tursiops truncatus) while foraging in a shallow water environment. Mar. Mammal Sci. 2019, 36, 305–314. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elliser, C.R.; Calambokidis, J.; D’Alessandro, D.N.; Duffield, D.A.; Huggins, J.L.; Rice, J.; Szczepaniak, I.; Webber, M. Prey-Related Asphyxiation in Harbor Porpoises (Phocoena phocoena) along the U.S. West Coast: Importance of American Shad (Alosa sapidissima) on Adult Female Harbor Porpoise Mortality. Oceans 2020, 1, 94-108. https://doi.org/10.3390/oceans1030008
Elliser CR, Calambokidis J, D’Alessandro DN, Duffield DA, Huggins JL, Rice J, Szczepaniak I, Webber M. Prey-Related Asphyxiation in Harbor Porpoises (Phocoena phocoena) along the U.S. West Coast: Importance of American Shad (Alosa sapidissima) on Adult Female Harbor Porpoise Mortality. Oceans. 2020; 1(3):94-108. https://doi.org/10.3390/oceans1030008
Chicago/Turabian StyleElliser, Cindy R., John Calambokidis, Dalin N. D’Alessandro, Deborah A. Duffield, Jessica L. Huggins, James Rice, Isidore Szczepaniak, and Marc Webber. 2020. "Prey-Related Asphyxiation in Harbor Porpoises (Phocoena phocoena) along the U.S. West Coast: Importance of American Shad (Alosa sapidissima) on Adult Female Harbor Porpoise Mortality" Oceans 1, no. 3: 94-108. https://doi.org/10.3390/oceans1030008
APA StyleElliser, C. R., Calambokidis, J., D’Alessandro, D. N., Duffield, D. A., Huggins, J. L., Rice, J., Szczepaniak, I., & Webber, M. (2020). Prey-Related Asphyxiation in Harbor Porpoises (Phocoena phocoena) along the U.S. West Coast: Importance of American Shad (Alosa sapidissima) on Adult Female Harbor Porpoise Mortality. Oceans, 1(3), 94-108. https://doi.org/10.3390/oceans1030008