


Effect of the Neck Linker on Processive Stepping of Kinesin Motor


Abstract
:1. Introduction
2. Methods
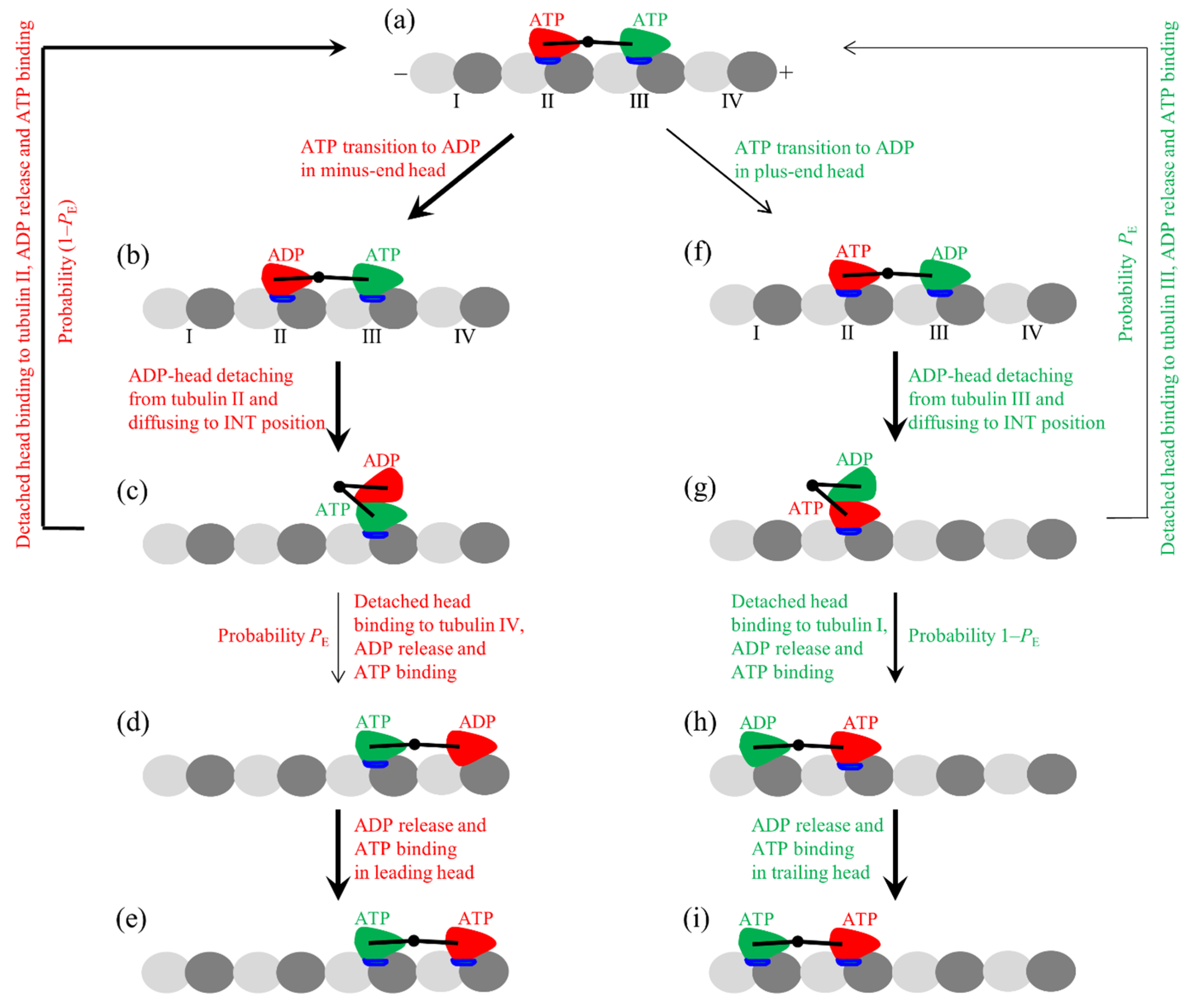
2.1. Model
- (i)
- The kinesin head in the nucleotide-free (ϕ), ATP, and ADP.Pi states has a high binding energy to MT [11,45,46], with the strong interaction between them inducing large conformational modulations of the local tubulin [47,48,49], while in adenosine diphosphate (ADP) state has a low binding energy to MTs [11,45,46], with the weak interaction between them inducing little conformational modulations of the tubulin [48,49]. The binding energy of the ADP-head to the tubulin with the large conformational modulations, which is denoted by Ew1, is much smaller than the binding energy of the ADP-head to the tubulin without the conformational modulations, which is denoted by Ew2 [48,49]. This implies that after ATP transition to ADP, the head has a very weak binding energy Ew1 to the local tubulin for a short time tr (in the order of 10 μs), and with the local tubulin returning elastically to the normally unmodulated one in time tr, the binding energy of the ADP-head to the local tubulin transits to Ew2 [48,49,50].
- (ii)
- The kinesin head in the ADP and ϕ states has an open nucleotide-binding pocket (NBP), while in the ATP and ADP.Pi states, it has a closed NBP and a large conformational change compared to that in the ADP/ϕ states [51,52,53]. However, for the ATP- or ADP.Pi-head with its NL in the backward or minus-ended orientation (e.g., for the leading or plus-ended head of the dimer in the two-heads-bound state), the NBP closing and large conformational change are prohibited [53]. The head without the large conformational change has an undocked NL [53,54,55,56] and a high binding energy to the partner ADP-head [57], while the head with the large conformational change has a docked NL [53,54,55,56] and a very low binding energy to the partner ADP-head [57].
- (iii)
- The kinesin head with the NL in the forward or plus-ended orientation has a much larger rate of ATP transition to ADP than the head with the NL not in the forward orientation, as explained as follows. Structural studies indicated that the NL of the kinesin head in the forward orientation clashes with a nucleotide-binding motif (a P-loop subdomain) in the ϕ orientation, resulting in the P-loop subdomain in the ATP-like orientation [10]. Thus, it is expected that the rate of ATP transition to ADP of the head with the NL in the forward orientation is significantly increased compared to that with the NL not in the forward NL orientation. This is consistent with the prior biochemical data showing that the deletion of the NL in the kinesin-1 head and that in the kinesin-3/KIF1a head significantly reduced the ATPase rate but had no effect on the rate of ADP release [10,58].
2.2. General Equations for Motor Dynamics
3. Results
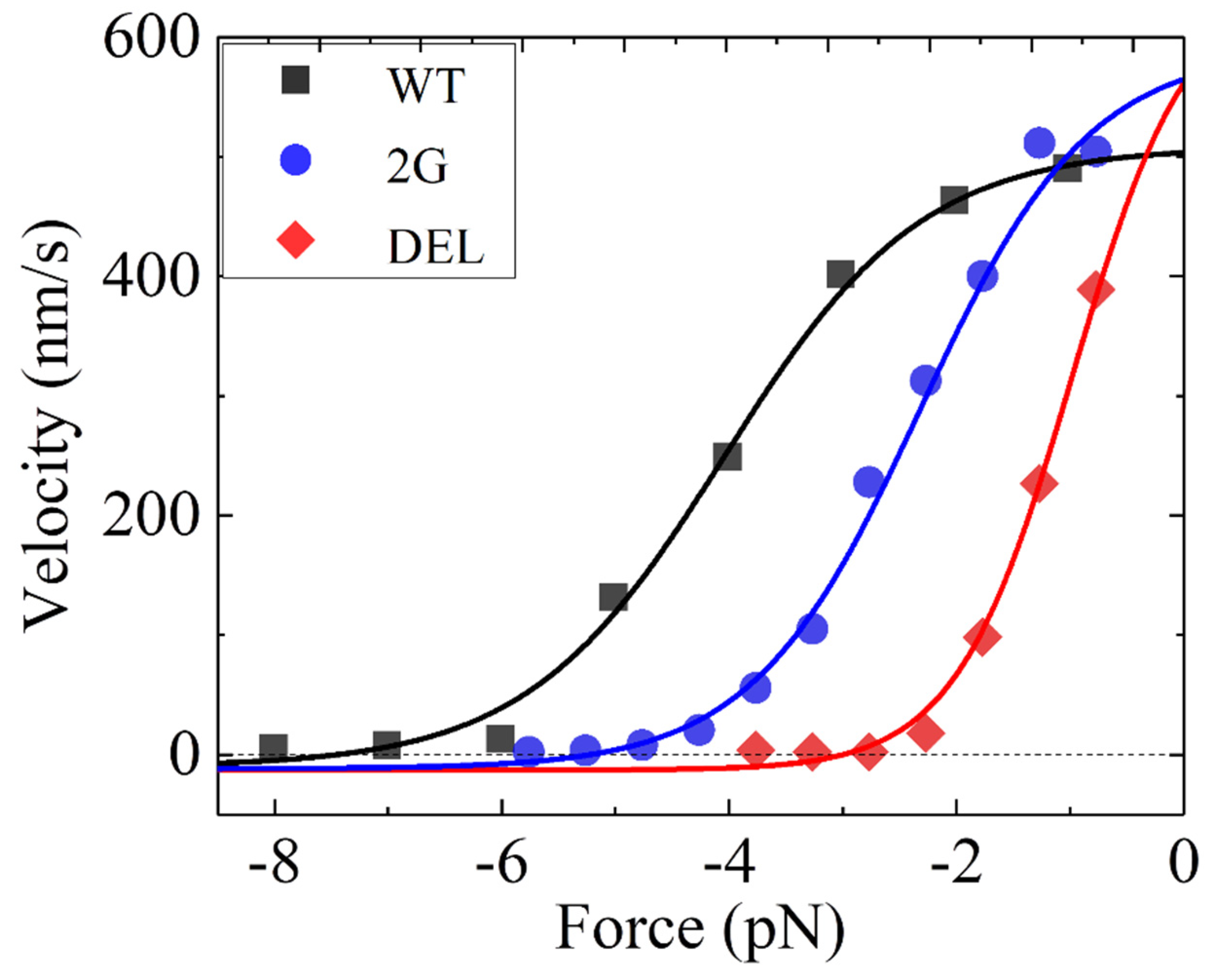
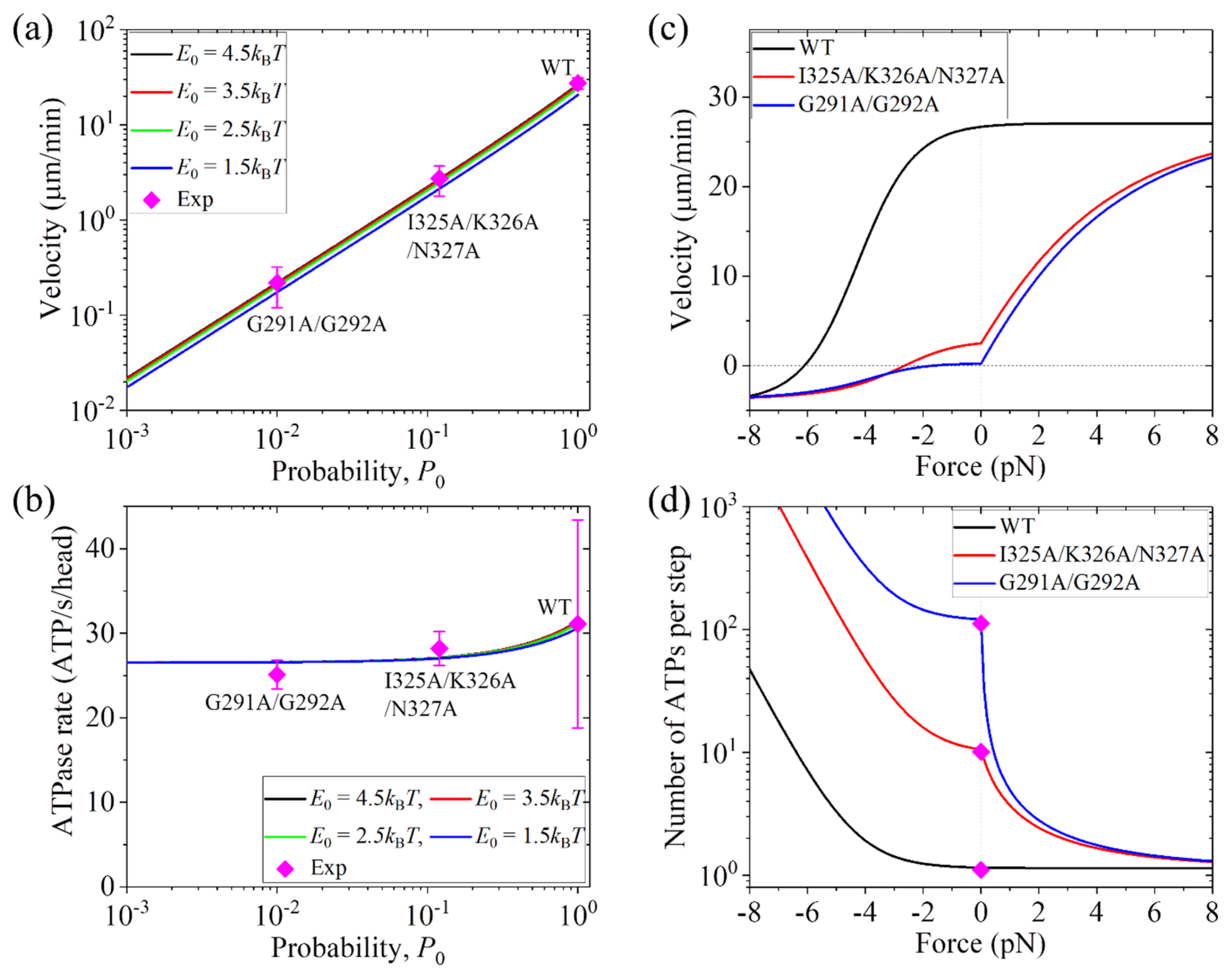
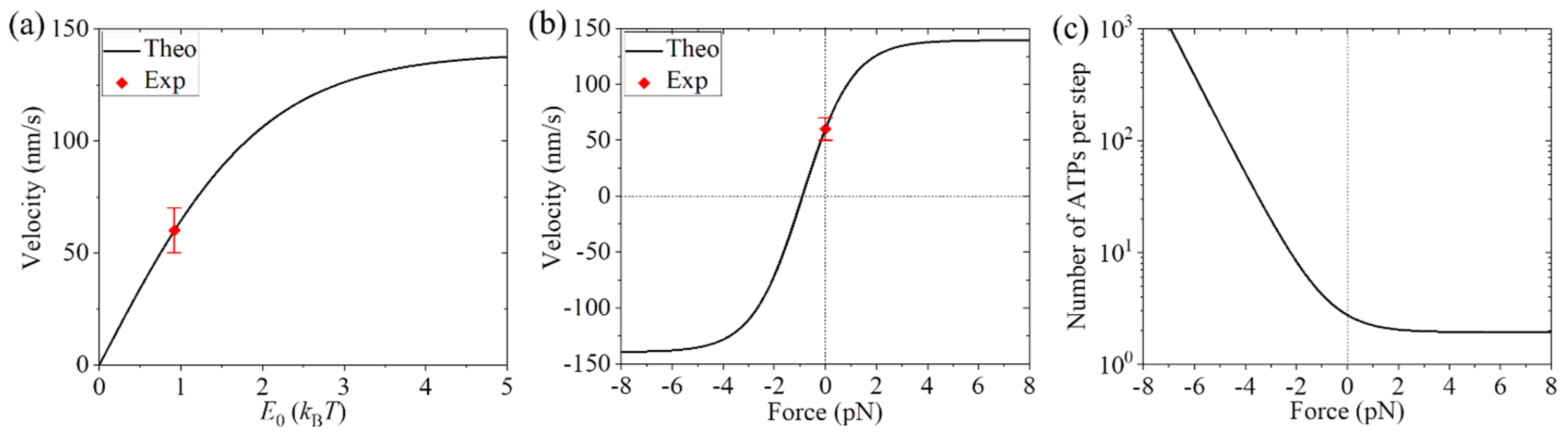
3.1. Dynamics of Kinesin-1 with Mutation or Deletion of Cover Strand Contributing to NL Docking
3.2. Dynamics of Kinesin-1 with NL Extension
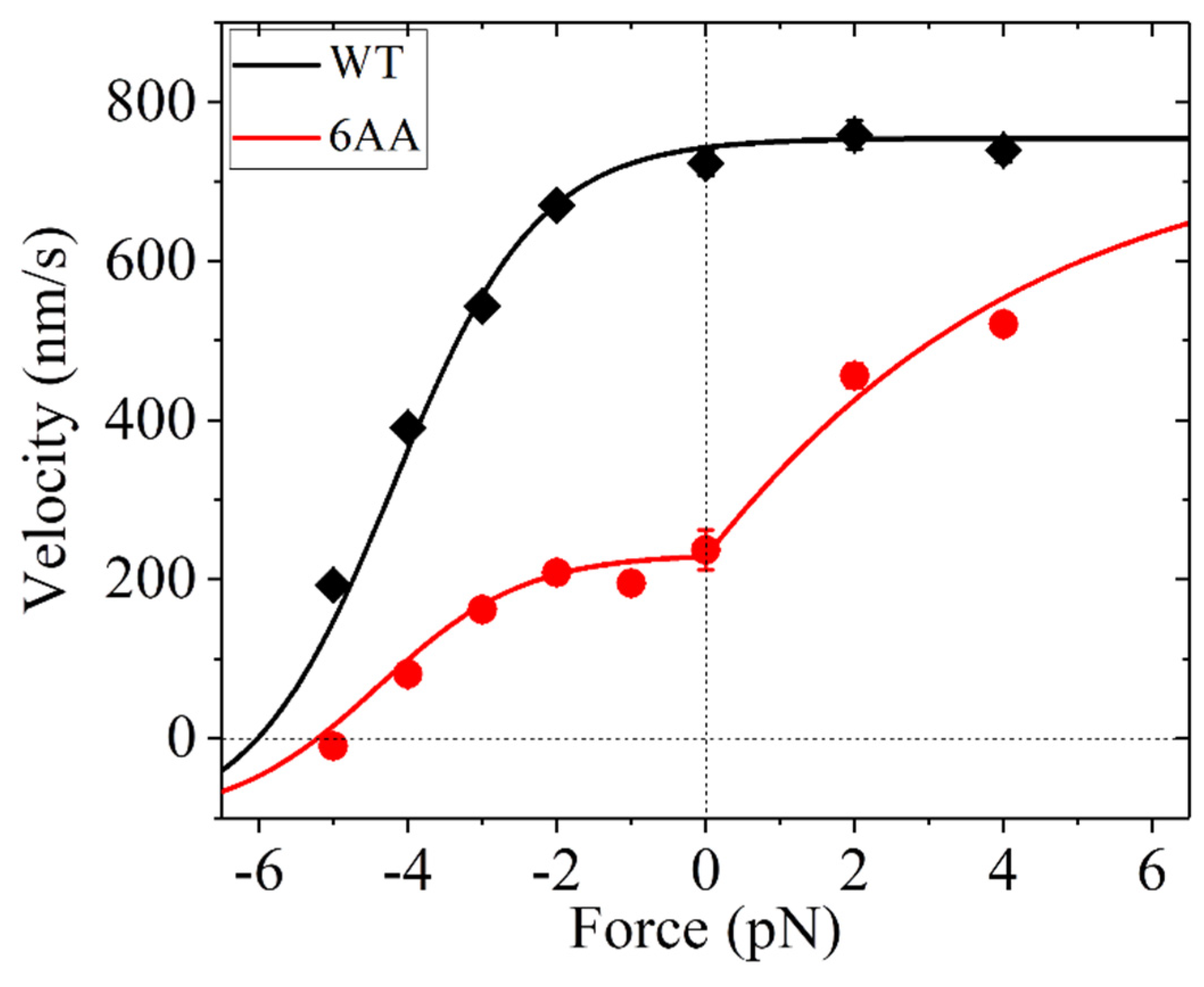
3.3. Dynamics of Kinesin-1 with NL Mutation
3.4. Dynamics of Kinesin-2 with NL Swapping
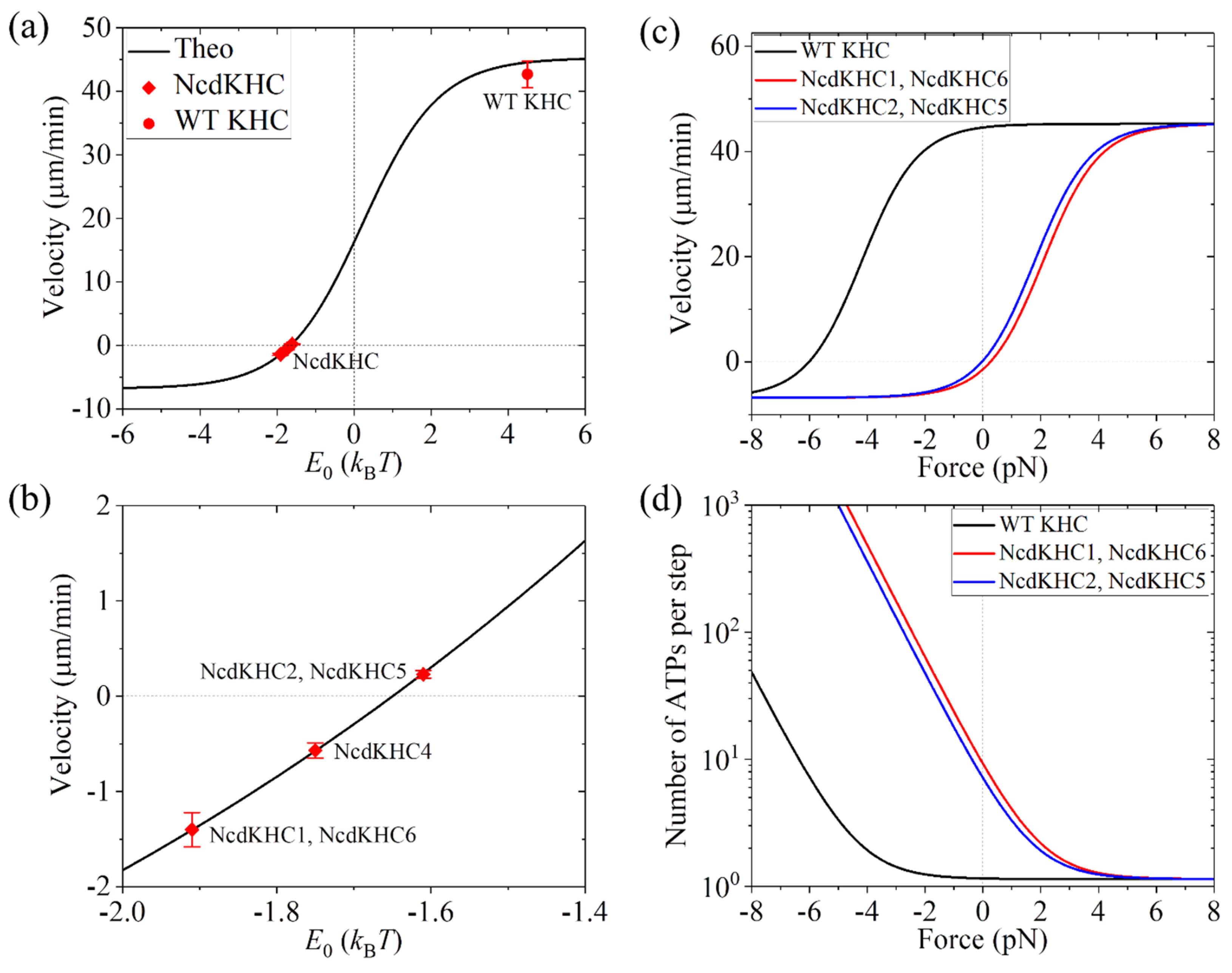
3.5. Dynamics of Motor with Ncd Stalk and Neck Joined to the Kinesin-1 Motor Domain
3.6. Dynamics of Motor with Kinesin-1 Neck and NL Joined to the Ncd Motor Domain
4. Discussion
4.1. Unidirectional Motility of Kinesin-6 MKLP2
4.2. Effect of NL Mutation on Motility of bi-Directional Kinesin-5 Cin8
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vale, R.D.; Reese, T.S.; Sheetz, M.P. Identification of a novel force-generating protein, kinesin, involved in microtubule-based motility. Cell 1985, 42, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Kozielski, F.; Sack, S.; Marx, A.; Thormahlen, M.; Schonbrunn, E.; Biou, V.; Thompson, A.; Mandelkow, E.-M.; Mandelkow, E. The crystal structure of dimeric kinesin and implications for microtubule-dependent motility. Cell 1997, 91, 985–994. [Google Scholar] [CrossRef]
- Lawrence, C.J.; Dawe, R.K.; Christie, K.R.; Cleveland, D.W.; Dawson, S.C.; Endow, S.A.; Wordeman, L. A standardized kinesin nomenclature. J. Cell Boil. 2004, 167, 19–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, J. The movement of kinesin along microtubules. Annu. Rev. Physiol. 1996, 58, 703–729. [Google Scholar] [CrossRef] [PubMed]
- Cross, R.A. The kinetic mechanism of kinesin. Trends Biochem. Sci. 2004, 29, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Asbury, C.L. Kinesin: World’s tiniest biped. Curr. Opin. Cell Biol. 2005, 17, 89–97. [Google Scholar] [CrossRef]
- Block, S.M. Kinesin motor mechanics: Binding, stepping, tracking, gating, and limping. Biophys. J. 2007, 92, 2986–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, P. Insight into the chemomechanical coupling mechanism of kinesin molecular motors. Commun. Theor. Phys. 2021, 73, 057601. [Google Scholar] [CrossRef]
- Gigant, B.; Wang, W.; Dreier, B.; Jiang, Q.; Pecqueur, L.; Plückthun, A.; Wang, C.; Knossow, M. Structure of a kinesin-tubulin complex and implications for kinesin motility. Nat. Struct. Mol. Biol. 2013, 20, 1001–1007. [Google Scholar] [CrossRef]
- Cao, L.; Wang, W.; Jiang, Q.; Wang, C.; Knossow, M.; Gigant, B. The structure of apo-kinesin bound to tubulin links the nucleotide cycle to movement. Nat. Commun. 2014, 5, 5364. [Google Scholar] [CrossRef]
- Cross, R.A. Mechanochemistry of the kinesin-1 ATPase. Biopolymers 2016, 105, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visscher, K.; Schnitzer, M.J.; Block, S.M. Single Kinesin Molecules Studied with a Molecular Force Clamp. Nature 1999, 400, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Coppin, C.M.; Pierce, D.W.; Hsu, L.; Vale, R.D. The load dependence of kinesin’s mechanical cycle. Proc. Natl. Acad. Sci. USA 1997, 94, 8539–8544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, M.; Higuchi, H.; Yanagida, T. Chemomechanical Coupling of the Forward and Backward Steps of Single Kinesin Molecules. Nat. Cell Biol. 2002, 4, 790–797. [Google Scholar] [CrossRef]
- Carter, N.J.; Cross, R.A. Mechanics of the Kinesin Step. Nature 2005, 435, 308–312. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Nishiyama, M.; Ishii, Y.; Yanagida, T. Entropy rectifies the Brownian steps of kinesin. Nat. Chem. Biol. 2005, 1, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.E.; Kolomeisky, A.B. Simple mechanochemistry describes the dynamics of kinesin molecules. Proc. Natl. Acad. Sci. USA 2001, 98, 7748–7753. [Google Scholar] [CrossRef] [Green Version]
- Liepelt, S.; Lipowsky, R. Kinesin’s network of chemomechanical motor cycles. Phys. Rev. Lett. 2007, 98, 258102. [Google Scholar] [CrossRef] [Green Version]
- Hyeon, C.; Onuchic, J.N. Internal strain regulates the nucleotide binding site of the kinesin leading head. Proc. Natl. Acad. Sci. USA 2007, 104, 2175–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumi, T. Design principles governing chemomechanical coupling of kinesin. Sci. Rep. 2017, 7, 1163. [Google Scholar] [CrossRef] [PubMed]
- Hyeon, C.; Klumpp, S.; Onuchic, J.N. Kinesin’s backsteps under mechanical load. Phys. Chem. Chem. Phys. 2009, 11, 4899–4910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Kaya, M.; Higuchi, H. A unified walking model for dimeric motor proteins. Biophys. J. 2018, 115, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, P.; Guo, S.-K.; Chen, H. ATP-concentration- and force-dependent chemomechanical coupling of kinesin molecular motors. J. Chem. Inf. Model. 2019, 59, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Mugnai, M.L.; Hyeon, C.; Hinczewski, M.; Thirumalai, D. Theoretical perspectives on biological machines. Rev. Mod. Phys. 2020, 92, 025001. [Google Scholar] [CrossRef] [Green Version]
- Vale, R.D.; Milligan, R.A. The way things move: Looking under the hood of molecular motor proteins. Science 2000, 288, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endow, S.A.; Barker, D.S. Processive and nonprocessive models of kinesin movement. Annu. Rev. Physiol. 2003, 65, 161–175. [Google Scholar] [CrossRef]
- Klumpp, L.M.; Hoenger, A.; Gilbert, S.P. Kinesin’s second step. Proc. Natl. Acad. Sci. USA 2004, 101, 3444–3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, S.S.; Fordyce, P.M.; Jefferson, G.M.; King, P.H.; Block, S.M. Stepping and stretching: How kinesin uses internal strain to walk processively. J. Biol. Chem. 2003, 278, 18550–18556. [Google Scholar] [CrossRef] [Green Version]
- Schief, W.R.; Howard, J. Conformational changes during kinesin motility. Curr. Opin. Cell Biol. 2001, 13, 19–28. [Google Scholar] [CrossRef]
- Mori, T.; Vale, R.D.; Tomishige, M. How kinesin waits between steps. Nature 2007, 450, 750–754. [Google Scholar] [CrossRef]
- Sack, S.; Muller, J.; Marx, A.; Thormahlen, M.; Mandelkow, E.-M.; Brady, S.T.; Mandelkow, E. X-ray structure of motor and neck domains from rat brain kinesin. Biochemistry 1997, 36, 16155–16165. [Google Scholar] [CrossRef] [PubMed]
- Sindelar, C.V.; Budny, M.J.; Rice, S.; Naber, N.; Fletterick, R.; Cooke, R. Two conformations in the human kinesin power stroke defined by X-ray crystallography and EPR spectroscopy. Nat. Struct. Biol. 2002, 9, 844–848. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.; Lang, M.J.; Karplus, M. Force generation in kinesin hinges on cover-neck bundle formation. Structure 2008, 16, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, A.S.; Appleyard, D.C.; Labno, A.K.; Georges, A.; Karplus, M.; Belcher, A.M.; Hwang, W.; Lang, M.J. Kinesin’s cover-neck bundle folds forward to generate force. Proc. Natl. Acad. Sci. USA 2008, 105, 19247–19252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budaitis, B.G.; Jariwala, S.; Reinemann, D.N.; Schimert, K.I.; Scarabelli, G.; Grant, B.J.; Sept, D.; Lang, M.J.; Verhey, K.J. Neck linker docking is critical for Kinesin-1 force generation in cells but at a cost to motor speed and processivity. eLife 2019, 8, e44146. [Google Scholar] [CrossRef] [PubMed]
- Clancy, B.E.; Behnke-Parks, W.M.; Andreasson, J.O.L.; Rosenfeld, S.S.; Block, S.M. A universal pathway for kinesin stepping. Nat. Struct. Mol. Biol. 2011, 18, 1020–1027. [Google Scholar] [CrossRef] [Green Version]
- Andreasson, J.O.L.; Milic, B.; Chen, G.-Y.; Guydosh, N.R.; Hancock, W.O.; Block, S.M. Examining kinesin processivity within a general gating framework. eLife 2015, 4, e07403. [Google Scholar] [CrossRef]
- Case, R.B.; Rice, S.; Hart, C.L.; Ly, B.; Vale, R.D. Role of the kinesin neck linker and catalytic core in microtubule-based motility. Curr. Biol. 2000, 10, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Andreasson, J.O.L.; Shastry, S.; Hancock, W.O.; Block, S.M. The mechanochemical cycle of mammalian kinesin-2 KIF3A/B under load. Curr. Biol. 2015, 25, 1166–1175. [Google Scholar] [CrossRef] [Green Version]
- Endow, S.A.; Waligora, K.W. Determinants of kinesin motor polarity. Science 1998, 281, 1200–1202. [Google Scholar] [CrossRef]
- Case, R.B.; Pierce, D.W.; Hom-Booher, N.; Hart, C.L.; Vale, R.D. The directional preference of kinesin motors is specified by an element outside of the motor catalytic domain. Cell 1997, 90, 959–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henningsen, U.; Schliwa, M. Reversal in the direction of movement of a molecular motor. Nature 1997, 389, 93–96. [Google Scholar] [CrossRef]
- Xie, P.; Guo, S.-K.; Chen, H. A generalized kinetic model for coupling between stepping and ATP hydrolysis of kinesin molecular motors. Int. J. Mol. Sci. 2019, 20, 4911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, P. Dynamics of kinesin motor proteins under longitudinal and sideways loads. J. Theor. Biol. 2021, 530, 110879. [Google Scholar] [CrossRef]
- Crevel, I.M.T.C.; Lockhart, A.; Cross, R.A. Weak and strong states of kinesin and Ncd. J. Mol. Biol. 1996, 257, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Sosa, H.; Peterman, E.J.G.; Moerner, W.E.; Goldstein, L.S.B. ADP-induced rocking of the kinesin motor domain revealed by single-molecule fluorescence polarization microscopy. Nat. Struct. Biol. 2001, 8, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Yajima, H.; Nitta, R.; Inoue, S.; Ogura, T.; Sato, C.; Hirokawa, N. X-ray and Cryo-EM structures reveal mutual conformational changes of kinesin and GTP-state microtubules upon binding. EMBO J. 2015, 34, 1270–1286. [Google Scholar] [CrossRef]
- Shi, X.-X.; Fu, Y.B.; Guo, S.K.; Wang, P.Y.; Chen, H.; Xie, P. Investigating role of conformational changes of microtubule in regulating its binding affinity to kinesin by all-atom molecular dynamics simulation. Proteins 2018, 86, 1127–1139. [Google Scholar] [CrossRef]
- Shi, X.-X.; Wang, P.-Y.; Chen, H.; Xie, P. Studies of conformational changes of tubulin induced by interaction with kinesin using atomistic molecular dynamics simulations. Int. J. Mol. Sci. 2021, 22, 6709. [Google Scholar] [CrossRef]
- Xie, P. Modeling processive motion of kinesin-13 MCAK and kinesin-14 Cik1-Kar3 molecular motors. Protein Sci. 2021, 30, 2092–2105. [Google Scholar] [CrossRef]
- Shang, Z.; Zhou, K.; Xu, C.; Csencsits, R.; Cochran, J.C.; Sindelar, C.V. High-resolution structures of kinesin on microtubules provide a basis for nucleotide-gated force-generation. eLife 2014, 3, e04686. [Google Scholar] [CrossRef] [PubMed]
- Kikkawa, M.; Hirokawa, N. High-resolution cryo-EM maps show the nucleotide binding pocket of KIF1A in open and closed conformations. EMBO J. 2006, 25, 4187–4194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benoit, M.P.M.H.; Asenjo, A.B.; Paydar, M.; Dhakal, S.; Kwok, B.H.; Sosa, H. Structural basis of mechano-chemical coupling by the mitotic kinesin KIF14. Nat. Commun. 2021, 12, 3637. [Google Scholar] [CrossRef] [PubMed]
- Rice, S.; Lin, A.W.; Safer, D.; Hart, C.L.; Naber, N.; Carragher, B.O.; Cain, S.M.; Pechatnikova, E.; Wilson-Kubalek, E.M.; Whittaker, M.; et al. A structural change in the kinesin motor protein that drives motility. Nature 1999, 402, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Tomishige, M.; Vale, R.D. Controlling kinesin by reversible disulfide crosslinking. Identifying the motility-producing conformational change. J. Cell Biol. 2000, 151, 1081–1092. [Google Scholar] [CrossRef] [Green Version]
- Asenjo, A.B.; Weinberg, Y.; Sosa, H. Nucleotide binding and hydrolysis induces a disorder-order transition in the kinesin neck-linker region. Nat. Struc. Mol. Biol. 2006, 13, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.-X.; Guo, S.-K.; Wang, P.-Y.; Chen, H.; Xie, P. All-atom molecular dynamics simulations reveal how kinesin transits from one-head-bound to two-heads-bound state. Proteins 2020, 88, 545–557. [Google Scholar] [CrossRef]
- Nitta, R.; Okada, Y.; Hirokawa, N. Structural model for strain-dependent microtubule activation of Mg-ADP release from kinesin. Nat. Struct. Mol. Biol. 2008, 15, 1067–1075. [Google Scholar] [CrossRef]
- Moyer, M.L.; Gilbert, S.P.; Johnson, K.A. Pathway of ATP hydrolysis by monomeric and dimeric kinesin. Biochemistry 1998, 37, 800–813. [Google Scholar] [CrossRef]
- Okada, Y.; Hirokawa, N. A processive single-headed motor: Kinesin superfamily protein KIF1A. Science 1999, 283, 1152–1157. [Google Scholar] [CrossRef]
- Yildiz, A.; Tomishige, M.; Gennerich, A.; Vale, R.D. Intramolecular strain coordinates kinesin stepping behavior along microtubules. Cell 2008, 134, 1030–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, S.S.; Jefferson, G.M.; King, P.H. ATP reorients the neck linker of kinesin in two sequential steps. J. Biol. Chem. 2001, 276, 40167–40174. [Google Scholar] [CrossRef] [Green Version]
- Xie, P. Molecular mechanism of processive stepping of kinesin motors. Symmetry 2021, 13, 1799. [Google Scholar] [CrossRef]
- Guo, S.-K.; Wang, P.-Y.; Xie, P. A model of processive movement of dimeric kinesin. J. Theor. Biol. 2017, 414, 62–75. [Google Scholar] [CrossRef]
- Rice, S.; Cui, Y.; Sindelar, C.; Naber, N.; Matuska, M.; Vale, R.; Cooke, R. Thermodynamic properties of the kinesin neck region docking to the catalytic core. Biophys. J. 2003, 84, 1844–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, W.; Lang, M.J.; Karplus, M. Kinesin motility is driven by subdomain dynamics. eLife 2017, 6, e28948. [Google Scholar] [CrossRef]
- Xie, P. Non-tight and tight chemomechanical couplings of biomolecular motors under hindering loads. J. Theor. Biol. 2020, 490, 110173. [Google Scholar] [CrossRef]
- Togashi, Y.; Flechsig, H. Coarse-Grained Protein Dynamics Studies Using Elastic Network Models. Int. J. Mol. Sci. 2018, 19, 3899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loutchko, D.; Flechsig, H. Allosteric communication inmolecular machines via information exchange: What can be learned from dynamical modeling. Biophys. Rev. 2020, 12, 443–452. [Google Scholar] [CrossRef]
- Endres, N.F.; Yoshioka, C.; Milligan, R.A.; Vale, R.D. A lever-arm rotation drives motility of the minus-end-directed kinesin Ncd. Nature 2006, 439, 875–878. [Google Scholar] [CrossRef]
- Gruneberg, U.; Neef, R.; Honda, R.; Nigg, E.A.; Barr, F.A. Relocation of aurora B from centromeres to the central spindle at the metaphase to anaphase transition requires MKlp2. J. Cell Biol. 2004, 166, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesario, J.M.; Jang, J.K.; Redding, B.; Shah, N.; Rahman, T.; McKim, K.S. Kinesin 6 family member Subito participates in mitotic spindle assembly and interacts with mitotic regulators. J. Cell Sci. 2006, 119, 4770–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummer, S.; Mayer, T.U. Cdk1 negatively regulates midzone localization of the mitotic kinesin Mklp2 and the chromosomal passenger complex. Curr. Biol. 2009, 19, 607–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, M.; Fung, S.Y.S.; Onishi, N.; Saya, H.; Lee, S.H. Targeting aurora B to the equatorial cortex by MKlp2 is required for cytokinesis. PLoS ONE 2013, 8, e64826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landino, J.; Norris, S.R.; Li, M.; Ballister, E.R.; Lampson, M.A.; Ohi, R. Two mechanisms coordinate the recruitment of the chromosomal passenger complex to the plane of cell division. Mol. Biol. Cell 2017, 28, 3634–3646. [Google Scholar] [CrossRef]
- Atherton, J.; Yu, I.-M.; Cook, A.; Muretta, J.M.; Joseph, A.; Major, J.; Sourigues, Y.; Clause, J.; Topf, M.; Rosenfeld, S.S.; et al. The divergent mitotic kinesin MKLP2 exhibits atypical structure and mechanochemistry. eLife 2017, 6, e27793. [Google Scholar] [CrossRef] [Green Version]
- Adriaans, I.E.; Hooikaas, P.J.; Aher, A.; Vromans, M.J.M.; van Es, R.M.; Grigoriev, I.; Akhmanova, A.; Lens, S.M.A. MKLP2 Is a Motile Kinesin that Transports the Chromosomal Passenger Complex during Anaphase. Curr. Biol. 2020, 30, 2628–2637. [Google Scholar] [CrossRef]
- Kapitein, L.C.; Peterman, E.J.; Kwok, B.H.; Kim, J.H.; Kapoor, T.M.; Schmidt, C.F. The bipolar mitotic kinesin Eg5 moves on both microtubules that it crosslinks. Nature 2005, 435, 114–118. [Google Scholar] [CrossRef]
- Van den Wildenberg, S.M.; Tao, L.; Kapitein, L.C.; Schmidt, C.F.; Scholey, J.M.; Peterman, E.J. The homotetrameric kinesin-5 KLP61F preferentially crosslinks microtubules into antiparallel orientations. Curr. Biol. 2008, 18, 1860–1864. [Google Scholar] [CrossRef] [Green Version]
- Roostalu, J.; Hentrich, C.; Bieling, P.; Telley, I.A.; Schiebel, E.; Surrey, T. Directional switching of the kinesin cin8 through motor coupling. Science 2011, 332, 94–99. [Google Scholar] [CrossRef]
- Gerson-Gurwitz, A.; Thiede, C.; Movshovich, N.; Fridman, V.; Podolskaya, M.; Danieli, T.; Lakamper, S.; Klopfenstein, D.R.; Schmidt, C.F.; Gheber, L. Directionality of individual kinesin-5 Cin8 motors is modulated by loop 8, ionic strength and microtubule geometry. EMBO J. 2011, 30, 4942–4954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridman, V.; Gerson-Gurwitz, A.; Shapira, O.; Movshovich, N.; Lakamper, S.; Schmidt, C.F.; Gheber, L. Kinesin-5 Kip1 is a bi-directional motor that stabilizes microtubules and tracks their plus-ends in vivo. J. Cell Sci. 2013, 126, 4147–4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edamatsu, M. Bidirectional motility of the fission yeast kinesin-5, Cut7. Biochem. Biophys. Res. Commun. 2014, 446, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Pandey, H.; Al-Bassam, J.; Gheber, L. Bidirectional motility of kinesin-5 motor proteins: Structural determinants, cumulative functions and physiological roles. Cell. Mol. Life Sci. 2018, 75, 1757–1771. [Google Scholar] [CrossRef] [PubMed]
- Britto, M.; Goulet, A.; Rizvi, S.; von Loeffelholz, O.; Moores, C.A.; Cross, R.A. Schizosaccharomyces pombe kinesin-5 switches direction using a steric blocking mechanism. Proc. Natl Acad. Sci. USA 2016, 113, E7483–E7489. [Google Scholar] [CrossRef] [Green Version]
- Goldstein-Levitin, A.; Pandey, H.; Allhuzaeel, K.; Kass, I.; Gheber, L. Intracellular functions and motile properties of bi-directional kinesin-5 Cin8 are regulated by neck linker docking. eLife 2021, 10, e71036. [Google Scholar] [CrossRef] [PubMed]
- Xie, P. A common ATP-dependent stepping model for kinesin-5 and kinesin-1: Mechanism of bi-directionality of kinesin-5. Biophys. Chem. 2021, 271, 106548. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Wang, P.; Xie, P. Effect of kinesin-5 tail domain on motor dynamics for antiparallel microtubule sliding. Int. J. Mol. Sci. 2021, 22, 7857. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | ||
|---|---|---|---|
| WT | 2G | DEL | |
| kD () | 250 | 250 | 250 |
| k(+) () | 62 * | 72 * | 79 * |
| k(−) | 0.01k(+) | 0.01k(+) | 0.01k(+) |
| E0 (kBT) | 4.8 * | 3.3 * | 2.1 * |
| (nm) | 4.7 * | 5.5 * | 7.9 * |
| Parameter | Value |
|---|---|
| kD () | 250 |
| k(+) () | 92 * |
| k(−) | 0.15k(+) |
| E0 (kBT) | 4.5 * |
| (nm) | 4.2 * |
| F0 (pN) | 3.8 * |
| 1 (for K1-WT) | |
| 0.37 * (for K1-6AA) |
| Parameter | Value |
|---|---|
| kD () | 250 |
| k(+) () | 88.5 * |
| k(−) | 0.15k(+) |
| E0 (kBT) | 3.05 * (for WT) 3.05/2 (for K2-KHC-1 and K2-KHC-2) |
| (nm) | 2 * |
| F0 (pN) | 4.5 * |
| 0.82 (for WT) * 0.82 (for K2-KHC-1) 1 (for K2-KHC-2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, P. Effect of the Neck Linker on Processive Stepping of Kinesin Motor. Biophysica 2023, 3, 46-68. https://doi.org/10.3390/biophysica3010004
Xie P. Effect of the Neck Linker on Processive Stepping of Kinesin Motor. Biophysica. 2023; 3(1):46-68. https://doi.org/10.3390/biophysica3010004
Chicago/Turabian StyleXie, Ping. 2023. "Effect of the Neck Linker on Processive Stepping of Kinesin Motor" Biophysica 3, no. 1: 46-68. https://doi.org/10.3390/biophysica3010004
APA StyleXie, P. (2023). Effect of the Neck Linker on Processive Stepping of Kinesin Motor. Biophysica, 3(1), 46-68. https://doi.org/10.3390/biophysica3010004