
Structural Insights into Plasticity and Discovery of Flavonoid Allosteric Inhibitors of Flavivirus NS2B–NS3 Protease

 ,
,  ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
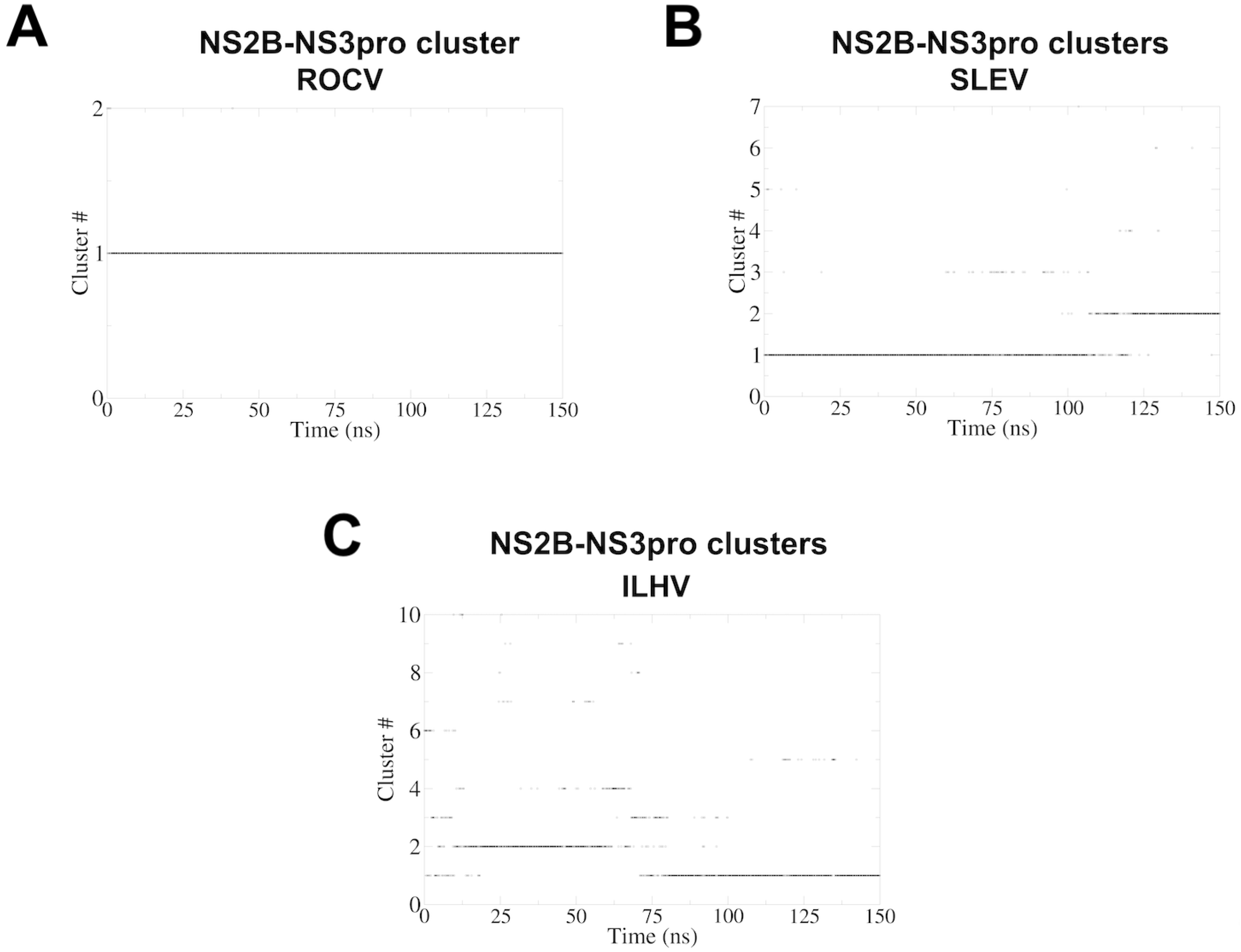
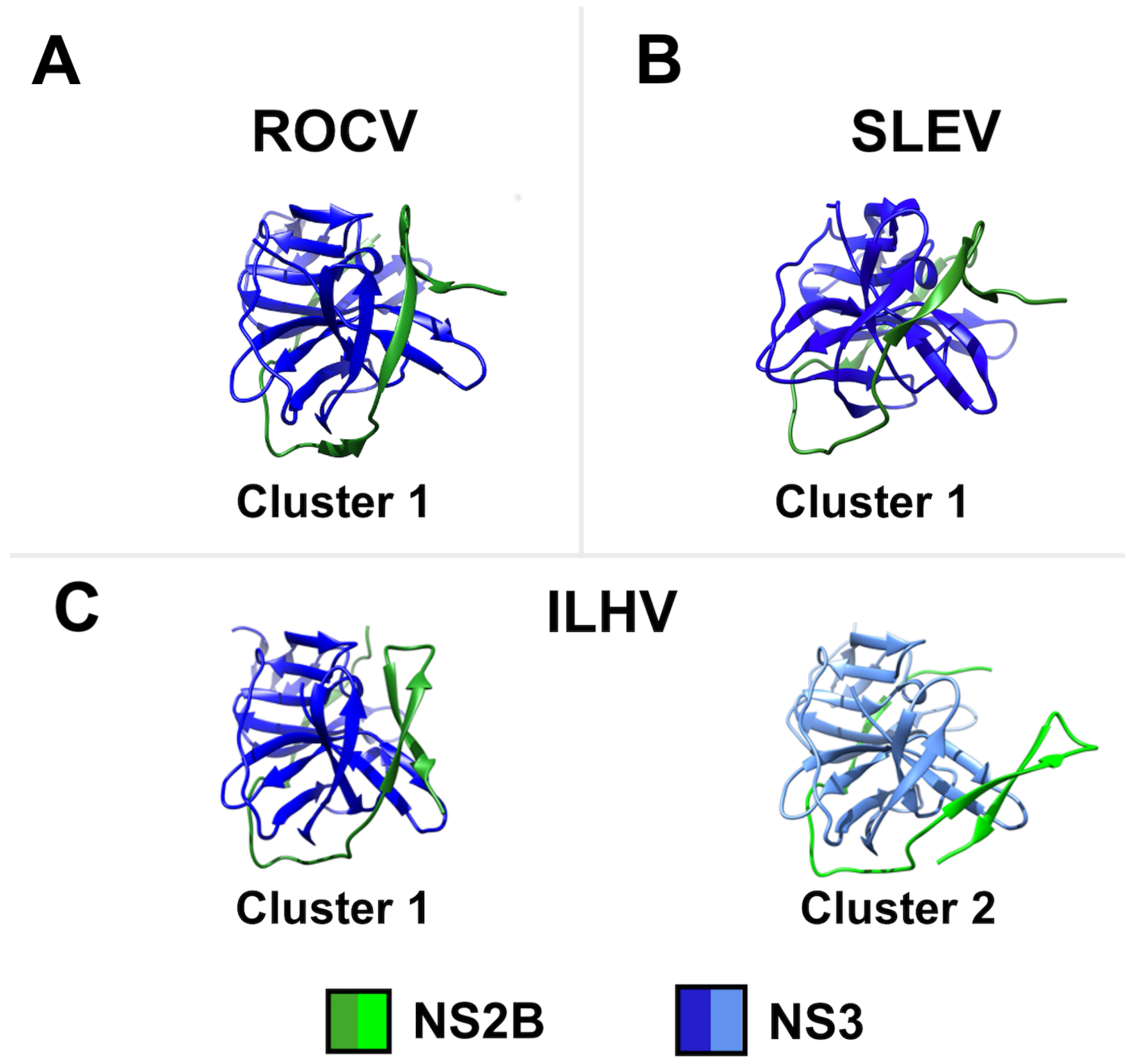
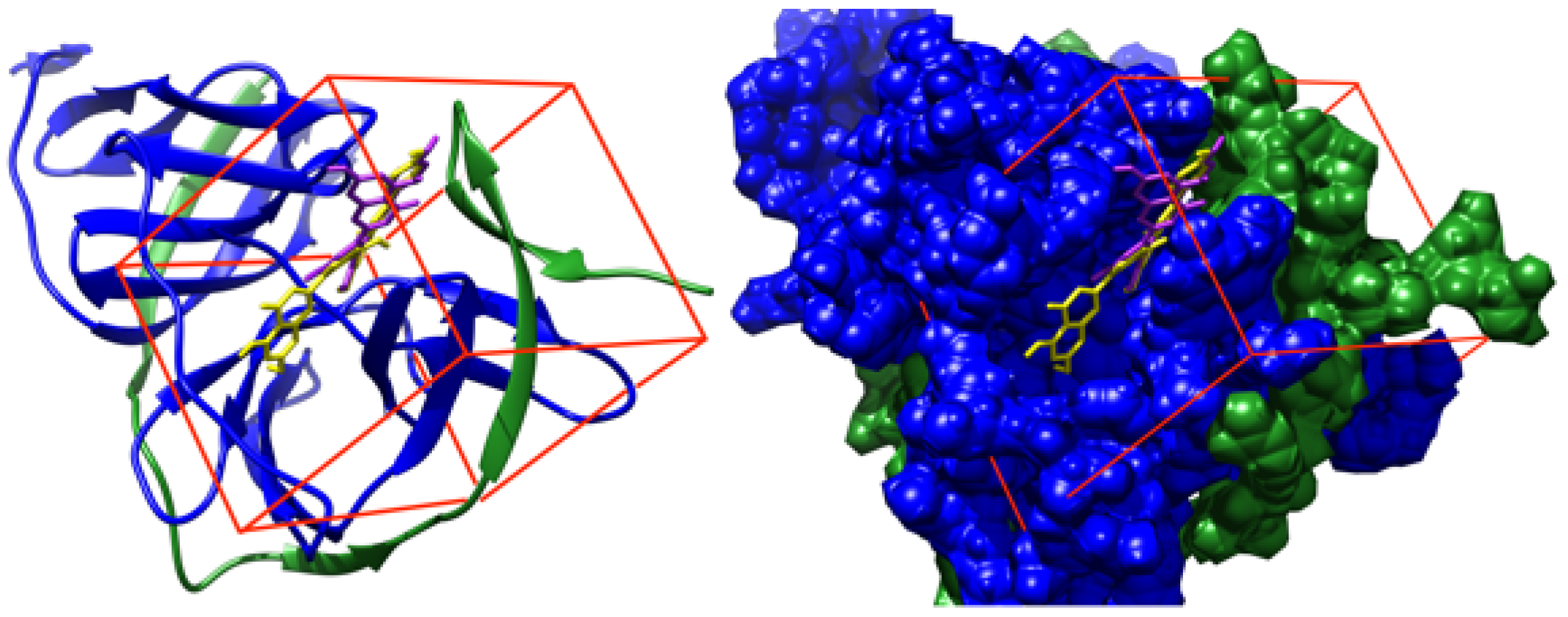
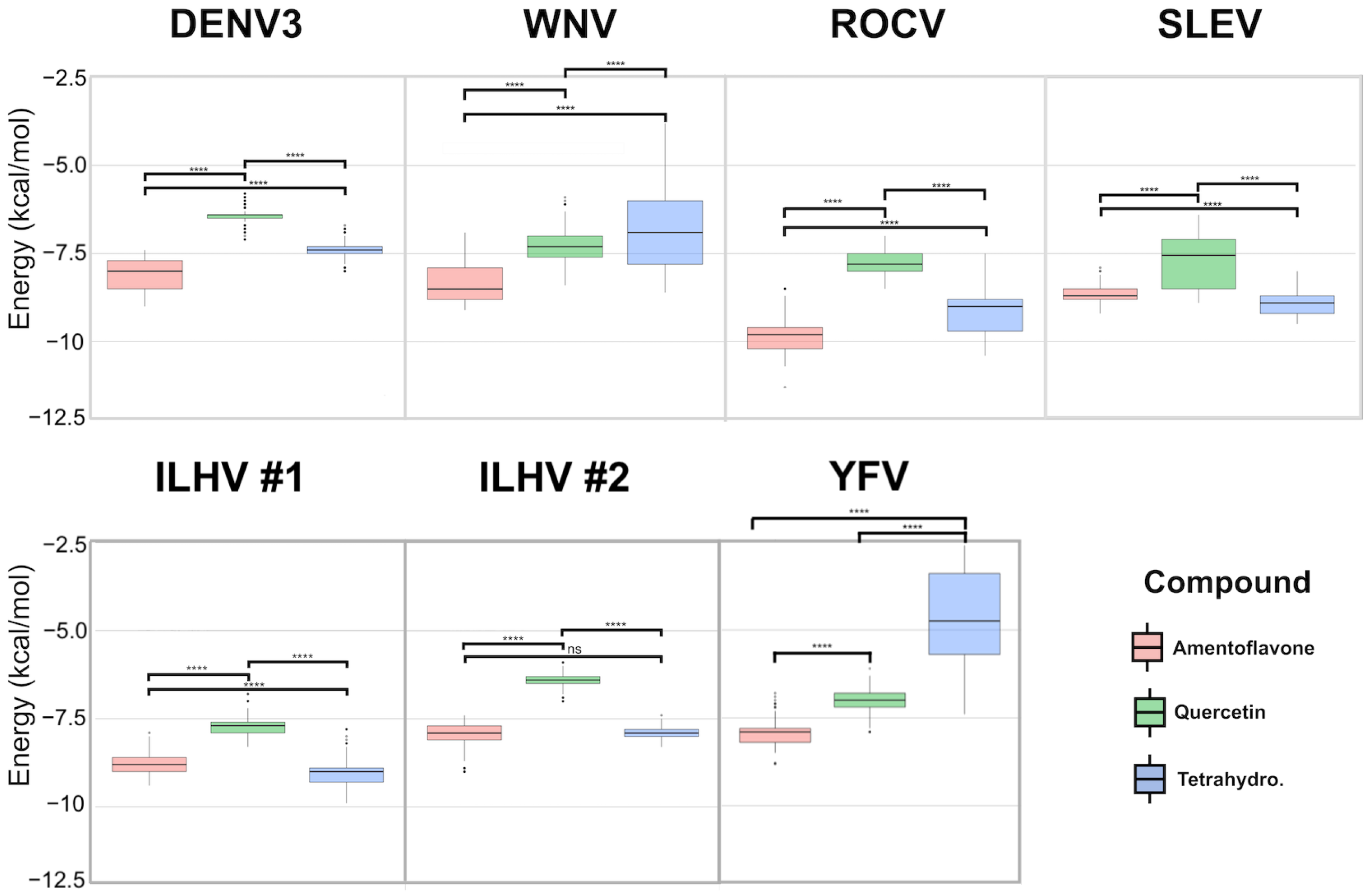
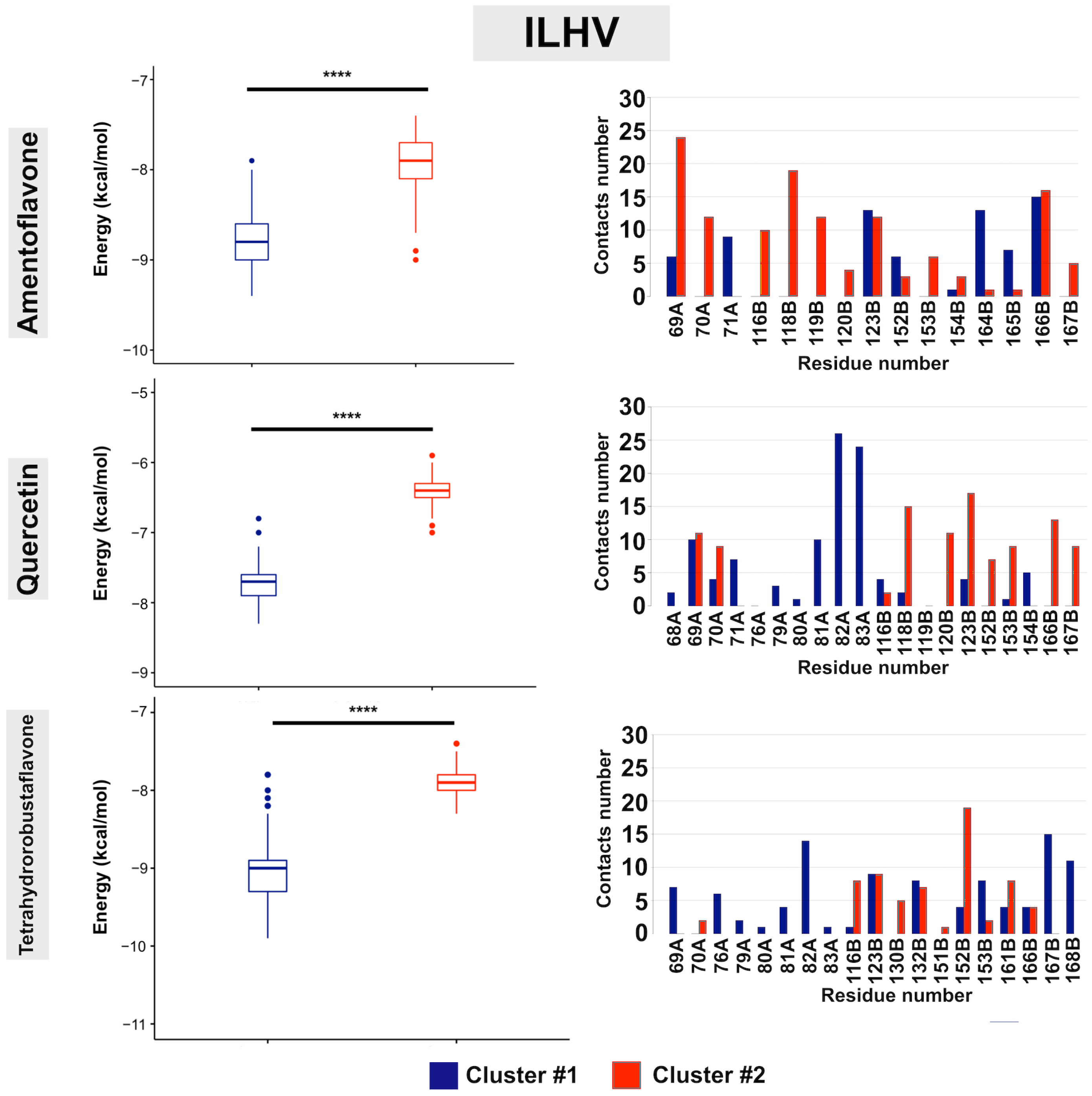
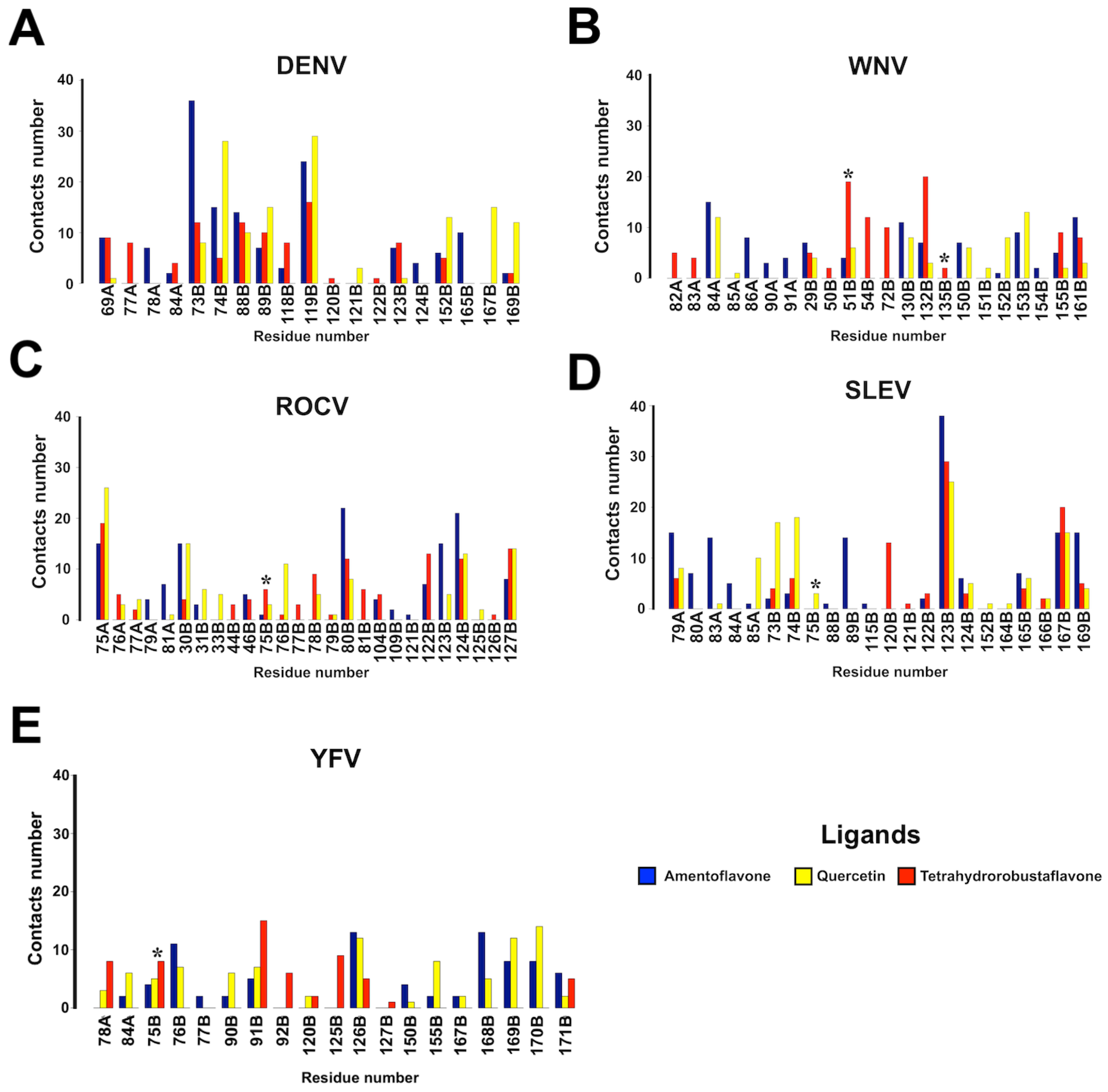
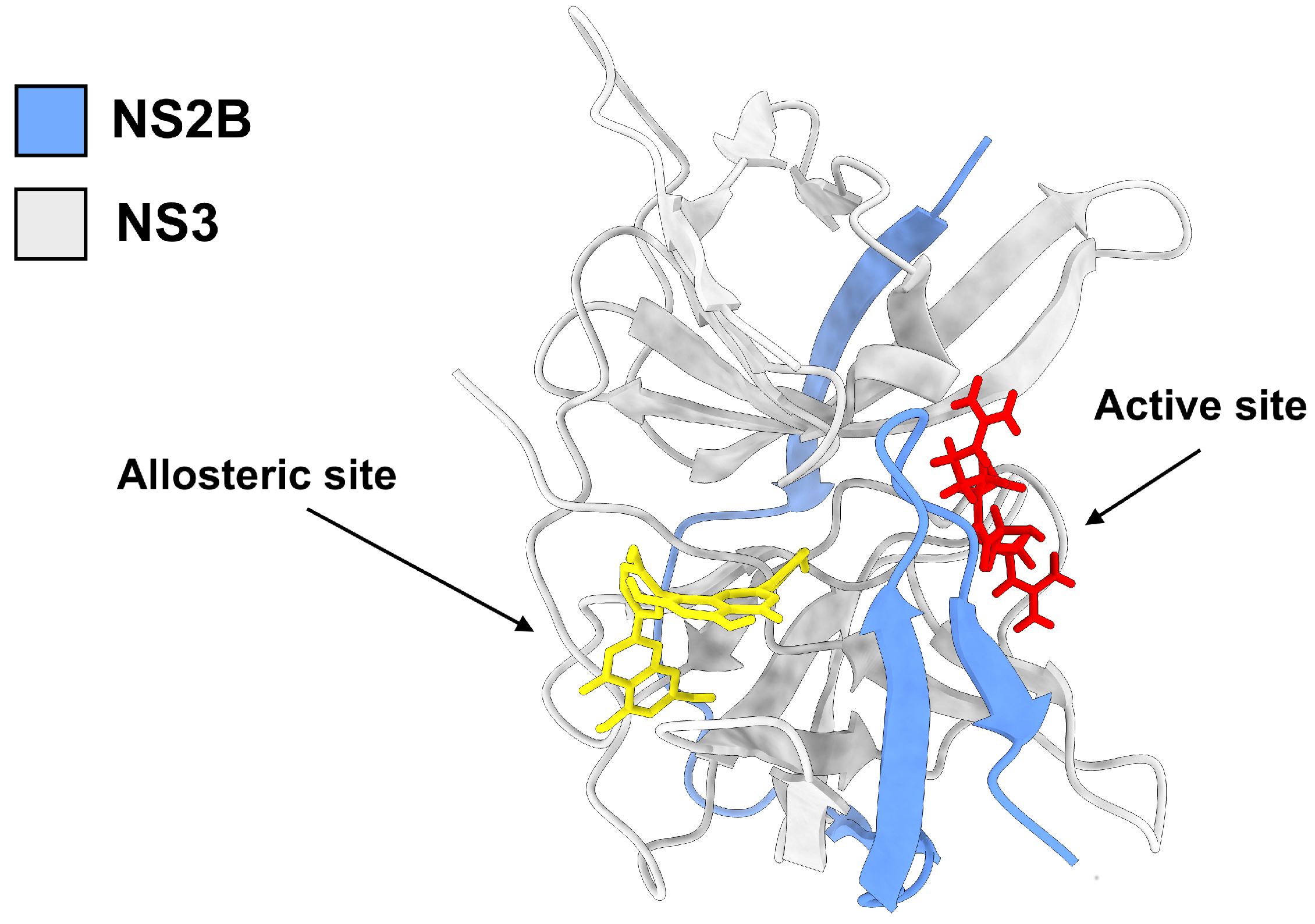
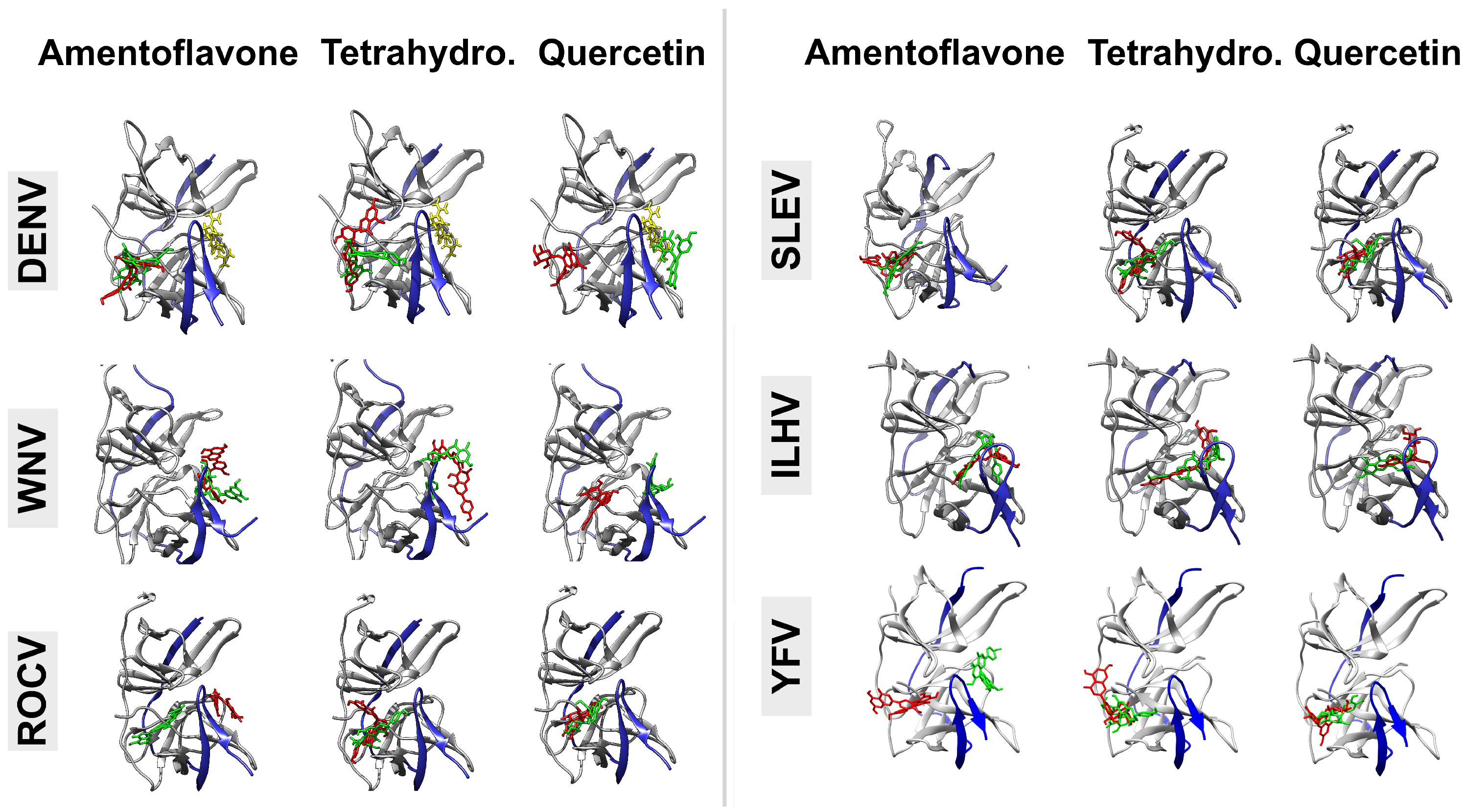
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, M.D.; Mazzon, M.; Jacobs, M.; Amara, A. Pathogenesis of Flavivirus Infections: Using and Abusing the Host Cell. Cell Host Microbe 2009, 5, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Fishburn, A.T.; Pham, O.H.; Kenaston, M.W.; Beesabathuni, N.S.; Shah, P.S. Let’s Get Physical: Flavivirus-Host Protein–Protein Interactions in Replication and Pathogenesis. Front. Microbiol. 2022, 13, 847588. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Phoo, W.W.; Luo, D. Functional interplay among the flavivirus NS3 protease, helicase, and cofactors. Virol. Sin. 2014, 29, 74–85. [Google Scholar] [CrossRef]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus Genome Organization, Expression, and Replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef]
- Li, L.; Lok, S.M.; Yu, I.M.; Zhang, Y.; Kuhn, R.J.; Chen, J.; Rossmann, M.G. The Flavivirus Precursor Membrane-Envelope Protein Complex: Structure and Maturation. Science 2008, 319, 1830–1834. [Google Scholar] [CrossRef]
- Chappell, K.J.; Stoermer, M.J.; Fairlie, D.P.; Young, P.R. Mutagenesis of the West Nile virus NS2B cofactor domain reveals two regions essential for protease activity. J. Gen. Virol. 2008, 89, 1010–1014. [Google Scholar] [CrossRef]
- Falgout, B.; Miller, R.; Lai, C.J. Deletion analysis of dengue virus type 4 nonstructural protein NS2B: Identification of a domain required for NS2B-NS3 protease activity. J. Virol. 1993, 67, 2034–2042. [Google Scholar] [CrossRef]
- Niyomrattanakit, P.; Winoyanuwattikun, P.; Chanprapaph, S.; Angsuthanasombat, C.; Panyim, S.; Katzenmeier, G. Identification of Residues in the Dengue Virus Type 2 NS2B Cofactor That Are Critical for NS3 Protease Activation. J. Virol. 2004, 78, 13708–13716. [Google Scholar] [CrossRef]
- Clum, S.; Ebner, K.E.; Padmanabhan, R. Cotranslational Membrane Insertion of the Serine Proteinase Precursor NS2B-NS3(Pro) of Dengue Virus Type 2 Is Required for Efficient in Vitro Processing and Is Mediated through the Hydrophobic Regions of NS2B. J. Biol. Chem. 1997, 272, 30715–30723. [Google Scholar] [CrossRef] [Green Version]
- Phoo, W.W.; Sahili, A.E.; Zhang, Z.; Chen, M.W.; Liew, C.W.; Lescar, J.; Vasudevan, S.G.; Luo, D. Crystal structures of full length DENV4 NS2B-NS3 reveal the dynamic interaction between NS2B and NS3. Antivir. Res. 2020, 182, 104900. [Google Scholar] [CrossRef]
- Tseng, A.C.; Nerurkar, V.R.; Neupane, K.R.; Kae, H.; Kaufusi, P.H. Potential Dual Role of West Nile Virus NS2B in Orchestrating NS3 Enzymatic Activity in Viral Replication. Viruses 2021, 13, 216. [Google Scholar] [CrossRef]
- Xu, T.; Sampath, A.; Chao, A.; Wen, D.; Nanao, M.; Luo, D.; Chene, P.; Vasudevan, S.G.; Lescar, J. Towards the Design of Flavivirus Helicase/NTPase Inhibitors: Crystallographic and Mutagenesis Studies of the Dengue Virus NS3 Helicase Catalytic Domain. In Novartis Foundation Symposia; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2008; pp. 87–101. [Google Scholar] [CrossRef]
- Saivish, M.V.; da Costa, V.G.; de Lima Menezes, G.; da Silva, R.A.; da Silva, G.C.D.; Moreli, M.L.; Sacchetto, L.; Pacca, C.C.; Vasilakis, N.; Nogueira, M.L. Rocio Virus: An Updated View on an Elusive Flavivirus. Viruses 2021, 13, 2293. [Google Scholar] [CrossRef]
- Saivish, M.V.; da Costa, V.G.; Rodrigues, R.L.; Féres, V.C.; Montoya-Diaz, E.; Moreli, M.L. Detection of Rocio Virus SPH 34675 during Dengue Epidemics, Brazil, 2011–2013. Emerg. Infect. Dis. 2020, 26, 797–799. [Google Scholar] [CrossRef]
- Sagini, J.P.; Arantes, P.R.; Pedebos, C.; Ligabue-Braun, R. Rocio Virus Encephalitis: In Silico Evidence for Drug Repurposing. Macromol 2022, 2, 100–112. [Google Scholar] [CrossRef]
- Milhim, B.H.G.A.; Estofolete, C.F.; da Rocha, L.C.; Liso, E.; Brienze, V.M.S.; Vasilakis, N.; Terzian, A.C.B.; Nogueira, M.L. Fatal Outcome of Ilheus Virus in the Cerebrospinal Fluid of a Patient Diagnosed with Encephalitis. Viruses 2020, 12, 957. [Google Scholar] [CrossRef]
- Diaz, A.; Coffey, L.L.; Burkett-Cadena, N.; Day, J.F. Reemergence of St. Louis Encephalitis Virus in the Americas. Emerg. Infect. Dis. 2018, 24, 2150. [Google Scholar] [CrossRef]
- Swetnam, D.M.; Stuart, J.B.; Young, K.; Maharaj, P.D.; Fang, Y.; Garcia, S.; Barker, C.M.; Smith, K.; Godsey, M.S.; Savage, H.M.; et al. Movement of St. Louis encephalitis virus in the Western United States, 2014–2018. PLoS Neglected Trop. Dis. 2020, 14, e0008343. [Google Scholar] [CrossRef]
- Odhar, H.A.; Ahjel, S.W.; Albeer, A.A.M.A.; Hashim, A.F.; Humadi, S.S. Virtual Screening of FDA Approved Drugs Library to Identify a Potential Inhibitor against NS2B-NS3 Protease of Yellow Fever Virus. J. Pharm. Res. Int. 2021, 33, 177–186. [Google Scholar] [CrossRef]
- Brecher, M.; Li, Z.; Liu, B.; Zhang, J.; Koetzner, C.A.; Alifarag, A.; Jones, S.A.; Lin, Q.; Kramer, L.D.; Li, H. A conformational switch high-throughput screening assay and allosteric inhibition of the flavivirus NS2B-NS3 protease. PLoS Pathog. 2017, 13, e1006411. [Google Scholar] [CrossRef]
- Santos, F.R.S.; Nunes, D.A.F.; Lima, W.G.; Davyt, D.; Santos, L.L.; Taranto, A.G.; Ferreira, J.M.S. Identification of Zika Virus NS2B-NS3 Protease Inhibitors by Structure-Based Virtual Screening and Drug Repurposing Approaches. J. Chem. Inf. Model. 2019, 60, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Virtual Ligand Screening of the National Cancer Institute (NCI) Compound Library Leads to the Allosteric Inhibitory Scaffolds of the West Nile Virus NS3 Proteinase ASSAY and Drug Development Technologies. Available online: https://www.liebertpub.com/doi/abs/10.1089/adt.2010.0309 (accessed on 17 April 2022).
- Shin, H.J.; Kim, M.H.; Lee, J.Y.; Hwang, I.; Yoon, G.Y.; Kim, H.S.; Kwon, Y.C.; Ahn, D.G.; Kim, K.D.; Kim, B.T.; et al. Structure-Based Virtual Screening: Identification of a Novel NS2B-NS3 Protease Inhibitor with Potent Antiviral Activity against Zika and Dengue Viruses. Microorganisms 2021, 9, 545. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shi, D.; Zhou, S.; Liu, H.; Liu, H.; Yao, X. Molecular dynamics simulations and novel drug discovery. Expert Opin. Drug Discov. 2017, 13, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Salo-Ahen, O.M.H.; Alanko, I.; Bhadane, R.; Bonvin, A.M.J.J.; Honorato, R.V.; Hossain, S.; Juffer, A.H.; Kabedev, A.; Lahtela-Kakkonen, M.; Larsen, A.S.; et al. Molecular Dynamics Simulations in Drug Discovery and Pharmaceutical Development. Processes 2020, 9, 71. [Google Scholar] [CrossRef]
- Amaro, R.E.; Baudry, J.; Chodera, J.; Demir, O.; McCammon, J.A.; Miao, Y.; Smith, J.C. Ensemble Docking in Drug Discovery. Biophys. J. 2018, 114, 2271–2278. [Google Scholar] [CrossRef]
- Śledź, P.; Caflisch, A. Protein structure-based drug design: From docking to molecular dynamics. Curr. Opin. Struct. Biol. 2018, 48, 93–102. [Google Scholar] [CrossRef]
- de Lima Menezes, G.; Saivish, M.V.; Nogueira, M.L.; da Silva, R.A. Virtual screening of small natural compounds against NS1 protein of DENV, YFV and ZIKV. J. Biomol. Struct. Dyn. 2022, 1–11. [Google Scholar] [CrossRef]
- de Lima Menezes, G.; da Silva, R.A. Identification of potential drugs against SARS-CoV-2 non-structural protein 1 (nsp1). J. Biomol. Struct. Dyn. 2020, 39, 5657–5667. [Google Scholar] [CrossRef]
- Gonçalves, R.L.; de Lima Menezes, G.; Sussuchi, L.; Moreli, M.L.; Mottin, M.; Andrade, C.H.; Pereira, M.; da Silva, R.A. Dynamic behavior of Dengue and Zika viruses NS1 protein reveals monomer–monomer interaction mechanisms and insights to rational drug design. J. Biomol. Struct. Dyn. 2019, 38, 4353–4363. [Google Scholar] [CrossRef]
- Santos, N.P.; Santos, L.H.; de Magalhães, M.T.Q.; Lei, J.; Hilgenfeld, R.; Ferreira, R.S.; Bleicher, L. Characterization of an Allosteric Pocket in Zika Virus NS2B-NS3 Protease. J. Chem. Inf. Model. 2022, 62, 945–957. [Google Scholar] [CrossRef]
- Lim, S.P.; Wang, Q.Y.; Noble, C.G.; Chen, Y.L.; Dong, H.; Zou, B.; Yokokawa, F.; Nilar, S.; Smith, P.; Beer, D.; et al. Ten years of dengue drug discovery: Progress and prospects. Antivir. Res. 2013, 100, 500–519. [Google Scholar] [CrossRef]
- Luo, D.; Vasudevan, S.G.; Lescar, J. The flavivirus NS2B–NS3 protease–helicase as a target for antiviral drug development. Antivir. Res. 2015, 118, 148–158. [Google Scholar] [CrossRef]
- Nie, S.; Yao, Y.; Wu, F.; Wu, X.; Zhao, J.; Hua, Y.; Wu, J.; Huo, T.; Lin, Y.L.; Kneubehl, A.R.; et al. Synthesis, Structure–Activity Relationships, and Antiviral Activity of Allosteric Inhibitors of Flavivirus NS2B–NS3 Protease. J. Med. Chem. 2021, 64, 2777–2800. [Google Scholar] [CrossRef]
- Takagi, Y.; Matsui, K.; Nobori, H.; Maeda, H.; Sato, A.; Kurosu, T.; Orba, Y.; Sawa, H.; Hattori, K.; Higashino, K.; et al. Discovery of novel cyclic peptide inhibitors of dengue virus NS2B-NS3 protease with antiviral activity. Bioorg. Med. Chem. Lett. 2017, 27, 3586–3590. [Google Scholar] [CrossRef]
- Yao, C.; Xi, C.; Hu, K.; Gao, W.; Cai, X.; Qin, J.; Lv, S.; Du, C.; Wei, Y. Inhibition of enterovirus 71 replication and viral 3C protease by quercetin. Virol. J. 2018, 15, 116. [Google Scholar] [CrossRef]
- Hammamy, M.Z.; Haase, C.; Hammami, M.; Hilgenfeld, R.; Steinmetzer, T. Development and Characterization of New Peptidomimetic Inhibitors of the West Nile Virus NS2B-NS3 Protease. ChemMedChem 2013, 8, 231–241. [Google Scholar] [CrossRef]
- Noble, C.G.; Seh, C.C.; Chao, A.T.; Shi, P.Y. Ligand-Bound Structures of the Dengue Virus Protease Reveal the Active Conformation. J. Virol. 2012, 86, 438–446. [Google Scholar] [CrossRef]
- Noske, G.D.; Gawriljuk, V.O.; Fernandes, R.S.; Furtado, N.D.; Bonaldo, M.C.; Oliva, G.; Godoy, A.S. Structural characterization and polymorphism analysis of the NS2B-NS3 protease from the 2017 Brazilian circulating strain of Yellow Fever virus. Biochim. Biophys. Acta 2020, 1864, 129521. [Google Scholar] [CrossRef]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2017, 27, 293–315. [Google Scholar] [CrossRef]
- Olsson, M.H.M.; Søndergaard, C.R.; Rostkowski, M.; Jensen, J.H. PROPKA3: Consistent treatment of internal and surface residues in empirical pKa predictions. J. Chem. Theory Comput. 2011, 7, 525–537. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1-2, 19–25. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Miyamoto, S.; Kollman, P.A. Settle: An analytical version of the SHAKE and RATTLE algorithm for rigid water models. J. Comput. Chem. 1992, 13, 952–962. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Hutter, J. Car-Parrinello molecular dynamics. WIREs Comput. Mol. Sci. 2011, 2, 604–612. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera?A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- da Silva, I.R.; Parise, M.R.; Pereira, M.; da Silva, R.A. Prospecting for new catechol-O-methyltransferase (COMT) inhibitors as a potential treatment for Parkinson’s disease: A study by molecular dynamics and structure-based virtual screening. J. Biomol. Struct. Dyn. 2020, 39, 5872–5891. [Google Scholar] [CrossRef]
- de Oliveira, F.M.; Procopio, V.O.; de Lima Menezes, G.; da Silva, R.A.; Kipnis, A.; Junqueira-Kipnis, A.P. Mycobacterium bovis PknG R242P Mutation Results in Structural Changes with Enhanced Virulence in the Mouse Model of Infection. Microorganisms 2022, 10, 673. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- de Sousa, L.R.F.; Wu, H.; Nebo, L.; Fernandes, J.B.; das Graças Fernandes da Silva, M.F.; Kiefer, W.; Kanitz, M.; Bodem, J.; Diederich, W.E.; Schirmeister, T.; et al. Flavonoids as noncompetitive inhibitors of Dengue virus NS2B-NS3 protease: Inhibition kinetics and docking studies. Bioorg. Med. Chem. 2015, 23, 466–470. [Google Scholar] [CrossRef]
- Free Download: BIOVIA Discovery Studio Visualizer [Online]. Available online: https://discover.3ds.com/discovery-studio-visualizer-download (accessed on 19 January 2023).
- Zhang, Y.; Skolnick, J. Scoring function for automated assessment of protein structure template quality. Proteins 2004, 57, 702–710. [Google Scholar] [CrossRef]
- Madden, T.; Tatusov, R.; Zhang, J. Applications of Network BLAST Server; Elsevier: Amsterdam, The Netherlands, 1996. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2021. [Google Scholar]
- Noble, C.G.; Shi, P.Y. Structural biology of dengue virus enzymes: Towards rational design of therapeutics. Antivir. Res. 2012, 96, 115–126. [Google Scholar] [CrossRef]
- Erbel, P.; Schiering, N.; D’Arcy, A.; Renatus, M.; Kroemer, M.; Lim, S.P.; Yin, Z.; Keller, T.H.; Vasudevan, S.G.; Hommel, U. Structural basis for the activation of flaviviral NS3 proteases from dengue and West Nile virus. Nat. Struct. Mol. Biol. 2006, 13, 372–373. [Google Scholar] [CrossRef]
- Gormaz, J.; Quintremil, S.; Rodrigo, R. Cardiovascular Disease: A Target for the Pharmacological Effects of Quercetin. CTMC 2015, 15, 1735–1742. [Google Scholar] [CrossRef]
- Mariee, A.D.; Abd-Allah, G.M.; El-Beshbishy, H.A. Protective effect of dietary flavonoid quercetin against lipemic-oxidative hepatic injury in hypercholesterolemic rats. Pharm. Biol. 2012, 50, 1019–1025. [Google Scholar] [CrossRef]
- Javadi, F.; Ahmadzadeh, A.; Eghtesadi, S.; Aryaeian, N.; Zabihiyeganeh, M.; Foroushani, A.R.; Jazayeri, S. The Effect of Quercetin on Inflammatory Factors and Clinical Symptoms in Women with Rheumatoid Arthritis: A Double-Blind, Randomized Controlled Trial. J. Am. Coll. Nutr. 2016, 36, 9–15. [Google Scholar] [CrossRef]
- Jaisinghani, R.N. Antibacterial properties of quercetin. Microbiol. Res. 2017, 8, 6877. [Google Scholar] [CrossRef]
- Mead, J.R.; McNair, N. Antiparasitic activity of flavonoids and isoflavones against Cryptosporidium parvum and Encephalitozoon intestinalis. FEMS Microbiol. Lett. 2006, 259, 153–157. [Google Scholar] [CrossRef]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; Mahdy, N.M.E.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef]
- Ezzati, M.; Yousefi, B.; Velaei, K.; Safa, A. A review on anti-cancer properties of Quercetin in breast cancer. Life Sci. 2020, 248, 117463. [Google Scholar] [CrossRef]
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; Biasi, S.D.; Roat, E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A. Quercetin and Cancer Chemoprevention. Evid. Based Complement. Altern. Med. 2011, 2011, 591356. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ashida, H.; Terao, J. Multitargeted cancer prevention by quercetin. Cancer Lett. 2008, 269, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Friel, H.; Lederman, H. A nutritional supplement formula for influenza A (H5N1) infection in humans. Med. Hypotheses 2006, 67, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Song, J.H.; Park, K.S.; Kwon, D.H. Inhibitory effects of quercetin 3-rhamnoside on influenza A virus replication. Eur. J. Pharm. Sci. 2009, 37, 329–333. [Google Scholar] [CrossRef]
- Johari, J.; Kianmehr, A.; Mustafa, M.; Abubakar, S.; Zandi, K. Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef]
- Zandi, K.; Teoh, B.T.; Sam, S.S.; Wong, P.F.; Mustafa, M.R.; AbuBakar, S. Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol. J. 2011, 8, 1–11. [Google Scholar] [CrossRef]
- dos Santos, A.E.; Kuster, R.M.; Yamamoto, K.A.; Salles, T.S.; Campos, R.; de Meneses, M.D.; Soares, M.R.; Ferreira, D. Quercetin and quercetin 3-O-glycosides from Bauhinia longifolia (Bong.) Steud. show anti-Mayaro virus activity. Parasites Vectors 2014, 7, 1–7. [Google Scholar] [CrossRef]
- Choi, H.J.; Song, J.H.; Kwon, D.H. Quercetin 3-rhamnoside Exerts Antiinfluenza A Virus Activity in Mice. Phytother. Res. 2011, 26, 462–464. [Google Scholar] [CrossRef]
- Wu, W.; Li, R.; Li, X.; He, J.; Jiang, S.; Liu, S.; Yang, J. Quercetin as an Antiviral Agent Inhibits Influenza A Virus (IAV) Entry. Viruses 2015, 8, 6. [Google Scholar] [CrossRef]
- Fanunza, E.; Iampietro, M.; Distinto, S.; Corona, A.; Quartu, M.; Maccioni, E.; Horvat, B.; Tramontano, E. Quercetin Blocks Ebola Virus Infection by Counteracting the VP24 Interferon-Inhibitory Function. Antimicrob. Agents Chemother. 2020, 64, e00530-20. [Google Scholar] [CrossRef]
- Bahun, M.; Jukić, M.; Oblak, D.; Kranjc, L.; Bajc, G.; Butala, M.; Bozovičar, K.; Bratkovič, T.; Podlipnik, Č.; Ulrih, N.P. Inhibition of the SARS-CoV-2 3CLpro main protease by plant polyphenols. Food Chem. 2022, 373, 131594. [Google Scholar] [CrossRef]
- Li, Z.; Cao, H.; Cheng, Y.; Zhang, X.; Zeng, W.; Sun, Y.; Chen, S.; He, Q.; Han, H. Inhibition of Porcine Epidemic Diarrhea Virus Replication and Viral 3C-Like Protease by Quercetin. Int. J. Mol. Sci. 2020, 21, 8095. [Google Scholar] [CrossRef]
- Lima, C.S.; Mottin, M.; de Assis, L.R.; de Moraes Roso Mesquita, N.C.; de Paula Sousa, B.K.; Coimbra, L.D.; dos Santos, K.B.; Zorn, K.M.; Guido, R.V.; Ekins, S.; et al. Flavonoids from Pterogyne nitens as Zika virus NS2B-NS3 protease inhibitors. Bioorg. Chem. 2021, 109, 104719. [Google Scholar] [CrossRef]
- Bhattacharya, K.; Bordoloi, R.; Chanu, N.R.; Kalita, R.; Sahariah, B.J.; Bhattacharjee, A. In silico discovery of 3 novel quercetin derivatives against papain-like protease, spike protein, and 3C-like protease of SARS-CoV-2. J. Genet. Eng. Biotechnol. 2022, 20, 43. [Google Scholar] [CrossRef]
- Saakre, M.; Mathew, D.; Ravisankar, V. Perspectives on plant flavonoid quercetin-based drugs for novel SARS-CoV-2. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 21. [Google Scholar] [CrossRef]
- Tang, Y.; Li, X.; Yuan, Y.; Zhang, H.; Zou, Y.; Xu, Z.; Xu, Q.; Song, J.; Deng, C.; Wang, Q. Network pharmacology-based predictions of active components and pharmacological mechanisms of Artemisia annua L. for the treatment of the novel Corona virus disease 2019 (COVID-19). BMC Complement. Med. Ther. 2022, 22, 56. [Google Scholar] [CrossRef]
- Verma, S.; Pandey, A.K. Factual insights of the allosteric inhibition mechanism of SARS-CoV-2 main protease by quercetin: An in silico analysis. 3 Biotech 2021, 11, 67. [Google Scholar] [CrossRef]
- Arwa, P.S.; Zeraik, M.L.; Ximenes, V.F.; da Fonseca, L.M.; da Silva Bolzani, V.; Silva, D.H.S. Redox-active biflavonoids from Garcinia brasiliensis as inhibitors of neutrophil oxidative burst and human erythrocyte membrane damage. J. Ethnopharmacol. 2015, 174, 410–418. [Google Scholar] [CrossRef]
- Chiang, C.H.; Yeh, C.Y.; Chung, J.G.; Chiang, I.T.; Hsu, F.T. Amentoflavone Induces Apoptosis and Reduces Expression of Anti-apoptotic and Metastasis-associated Proteins in Bladder Cancer. Anticancer Res. 2019, 39, 3641–3649. [Google Scholar] [CrossRef]
- Funakoshi-Tago, M.; Okamoto, K.; Izumi, R.; Tago, K.; Yanagisawa, K.; Narukawa, Y.; Kiuchi, F.; Kasahara, T.; Tamura, H. Anti-inflammatory activity of flavonoids in Nepalese propolis is attributed to inhibition of the IL-33 signaling pathway. Int. Immunopharmacol. 2015, 25, 189–198. [Google Scholar] [CrossRef]
- Salleh, W.M.N.H.W.; Abed, S.A.; Taher, M.; Kassim, H.; Tawang, A. The phytochemistry and biological diversity of Ferulago genus (Apiaceae): A systematic review. J. Pharm. Pharmacol. 2021, 73, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Liu, C.; Zhai, Y.; Li, L.; Wang, X.; Feng, W. Protection effect of amentoflavone in Selaginella tamariscina against TNF-alpha-induced vascular injury of endothelial cells. Yao Xue Xue Bao = Acta Pharm. Sin. 2013, 48, 1503–1509. [Google Scholar]
- Jung, H.J.; Sung, W.S.; Yeo, S.H.; Kim, H.S.; Lee, I.S.; Woo, E.R.; Lee, D.G. Antifungal effect of amentoflavone derived fromSelaginella tamariscina. Arch. Pharm. Res. 2006, 29, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Rizk, Y.S.; Santos-Pereira, S.; Gervazoni, L.; de Jesus Hardoim, D.; de Oliveira Cardoso, F.; da Silva Freitas de Souza, C.; Pelajo-Machado, M.; Carollo, C.A.; de Arruda, C.C.P.; Almeida-Amaral, E.E.; et al. Amentoflavone as an Ally in the Treatment of Cutaneous Leishmaniasis: Analysis of Its Antioxidant/Prooxidant Mechanisms. Front. Cell. Infect. Microbiol. 2021, 11, 615814. [Google Scholar] [CrossRef]
- Rizk, Y.S.; Fischer, A.; de Castro Cunha, M.; Rodrigues, P.O.; Marques, M.C.S.; de Fátima Cepa Matos, M.; Kadri, M.C.T.; Carollo, C.A.; de Arruda, C.C.P. In vitro activity of the hydroethanolic extract and biflavonoids isolated from Selaginella sellowii on Leishmania (Leishmania) amazonensis. Mem. Inst. Oswaldo Cruz 2014, 109, 1050–1056. [Google Scholar] [CrossRef]
- Lin, Y.M.; Flavin, M.T.; Schure, R.; Chen, F.C.; Sidwell, R.; Barnard, D.I.; Huffmann, J.H.; Kern, E.R. Antiviral Activities of Biflavonoids. Planta Medica 1999, 65, 120–125. [Google Scholar] [CrossRef]
- Sun, C.M.; Yu, S.L.; Ou, J.C.; Syu, W.J. Test of the biflavones from Selaginella moellendorffii on the in vitro inhibition of HIV-1 protease. J. Tradit. Chin. Med. 1995, 6, 223–230. [Google Scholar]
- Jayadevappa, M.K.; Karkera, P.R.; Siddappa, R.Y.; Telkar, S.; Karunakara, P. Investigation of plant flavonoids as potential dengue protease inhibitors. J. Herbmed Pharmacol. 2020, 9, 366–373. [Google Scholar] [CrossRef]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Computer aided identification of potential SARS CoV-2 main protease inhibitors from diterpenoids and biflavonoids of Torreya nucifera leaves. J. Biomol. Struct. Dyn. 2020, 40, 2647–2662. [Google Scholar] [CrossRef]
- Park, J.; Park, R.; Jang, M.; Park, Y.I.; Park, Y. Coronavirus enzyme inhibitors-experimentally proven natural compounds from plants. J. Microbiol. 2022, 60, 347–354. [Google Scholar] [CrossRef]
- Saravanan, K.M.; Zhang, H.; Senthil, R.; Vijayakumar, K.K.; Sounderrajan, V.; Wei, Y.; Shakila, H. Structural basis for the inhibition of SARS-CoV2 main protease by Indian medicinal plant-derived antiviral compounds. J. Biomol. Struct. Dyn. 2020, 40, 1970–1978. [Google Scholar] [CrossRef]
- Swargiary, A.; Mahmud, S.; Saleh, M.A. Screening of phytochemicals as potent inhibitor of 3-chymotrypsin and papain-like proteases of SARS-CoV2: An in silico approach to combat COVID-19. J. Biomol. Struct. Dyn. 2020, 40, 2067–2081. [Google Scholar] [CrossRef]
- Lin, Y.M.; Zembower, D.E.; Flavin, M.T.; Schure, R.M.; Anderson, H.M.; Korba, B.E.; Chen, F.C. Robustaflavone, a naturally occurring biflavanoid, is a potent non-nucleoside inhibitor of hepatitis B virus replication in vitro. Bioorg. Med. Chem. Lett. 1997, 7, 2325–2328. [Google Scholar] [CrossRef]
- Shivanika, C.; Deepak, C.S.; Ragunathan, V.; Tiwari, P.; Sumitha, A.; Brindha, D.V. Molecular docking, validation, dynamics simulations, and pharmacokinetic prediction of natural compounds against the SARS-CoV-2 main-protease. J. Biomol. Struct. Dyn. 2020, 40, 585–611. [Google Scholar] [CrossRef]
- Menezes, J.C.; Campos, V.R. Natural biflavonoids as potential therapeutic agents against microbial diseases. Sci. Total. Environ. 2021, 769, 145168. [Google Scholar] [CrossRef]
- Mondal, S.; Karmakar, A.; Mallick, T.; Begum, N.A. Exploring the efficacy of naturally occurring biflavone based antioxidants towards the inhibition of the SARS-CoV-2 spike glycoprotein mediated membrane fusion. Virology 2021, 556, 133–139. [Google Scholar] [CrossRef]
- Ristovski, J.T.; Matin, M.M.; Kong, R.; Kusturica, M.P.; Zhang, H. In vitro testing and computational analysis of specific phytochemicals with antiviral activities considering their possible applications against COVID-19. S. Afr. J. Bot. 2022, 151, 248–258. [Google Scholar] [CrossRef]
- Coluccia, A.; Puxeddu, M.; Nalli, M.; Wei, C.K.; Wu, Y.H.; Mastrangelo, E.; Elamin, T.; Tarantino, D.; Bugert, J.J.; Schreiner, B.; et al. Discovery of Zika Virus NS2B/NS3 Inhibitors That Prevent Mice from Life-Threatening Infection and Brain Damage. ACS Med. Chem. Lett. 2020, 11, 1869–1874. [Google Scholar] [CrossRef]
- Scaturro, P.; Cortese, M.; Chatel-Chaix, L.; Fischl, W.; Bartenschlager, R. Dengue Virus Non-structural Protein 1 Modulates Infectious Particle Production via Interaction with the Structural Proteins. PLoS Pathog. 2015, 11, e1005277. [Google Scholar] [CrossRef]
- Fassio, A.V.; Santos, L.H.; Silveira, S.A.; Ferreira, R.S.; de Melo-Minardi, R.C. nAPOLI: A graph-based strategy to detect and visualize conserved protein-ligand interactions in large-scale. IEEE/ACM Trans. Comput. Biol. Bioinform. 2019, 17, 1317–1328. [Google Scholar] [CrossRef]
- Durrant, J.D.; McCammon, J.A. BINANA: A novel algorithm for ligand-binding characterization. J. Mol. Graph. Model. 2011, 29, 888–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | GenBank Accession Code |
|---|---|
| ROCV | gb:AY632542 |
| ILHV | gb:MH932545 |
| SLEV | gb:KM267635 |
| Virus | Water Molecules | Na+ Ions |
|---|---|---|
| ROCV | 54,186 | 6 |
| ILHV | 49,953 | 1 |
| SLEV | 45,303 | 4 |
| Name | Structure | Formula | Molecular Weight |
|---|---|---|---|
| Amentoflavone |  | C30H18O10 | 538.0899968 |
| Tetrahydrorobustaflavone |  | C30H22O10 | 542.12129692 |
| Quercetin |  | C15H10O7 | 302.04265268 |
| Species (Cluster) | Residues in Favored Region | Residues in Allowed Region | Clashscore † (Percentile) ‡ |
|---|---|---|---|
| ROCV | 90.9% | 98.5% | 2.69 (98th) |
| SLEV | 93% | 99% | 3.32 (97th) |
| ILHV (1) | 92.35% | 99.5% | 1.33 (99th) |
| ILHV (2) | 95% | 100% | 4.66 (95th) |
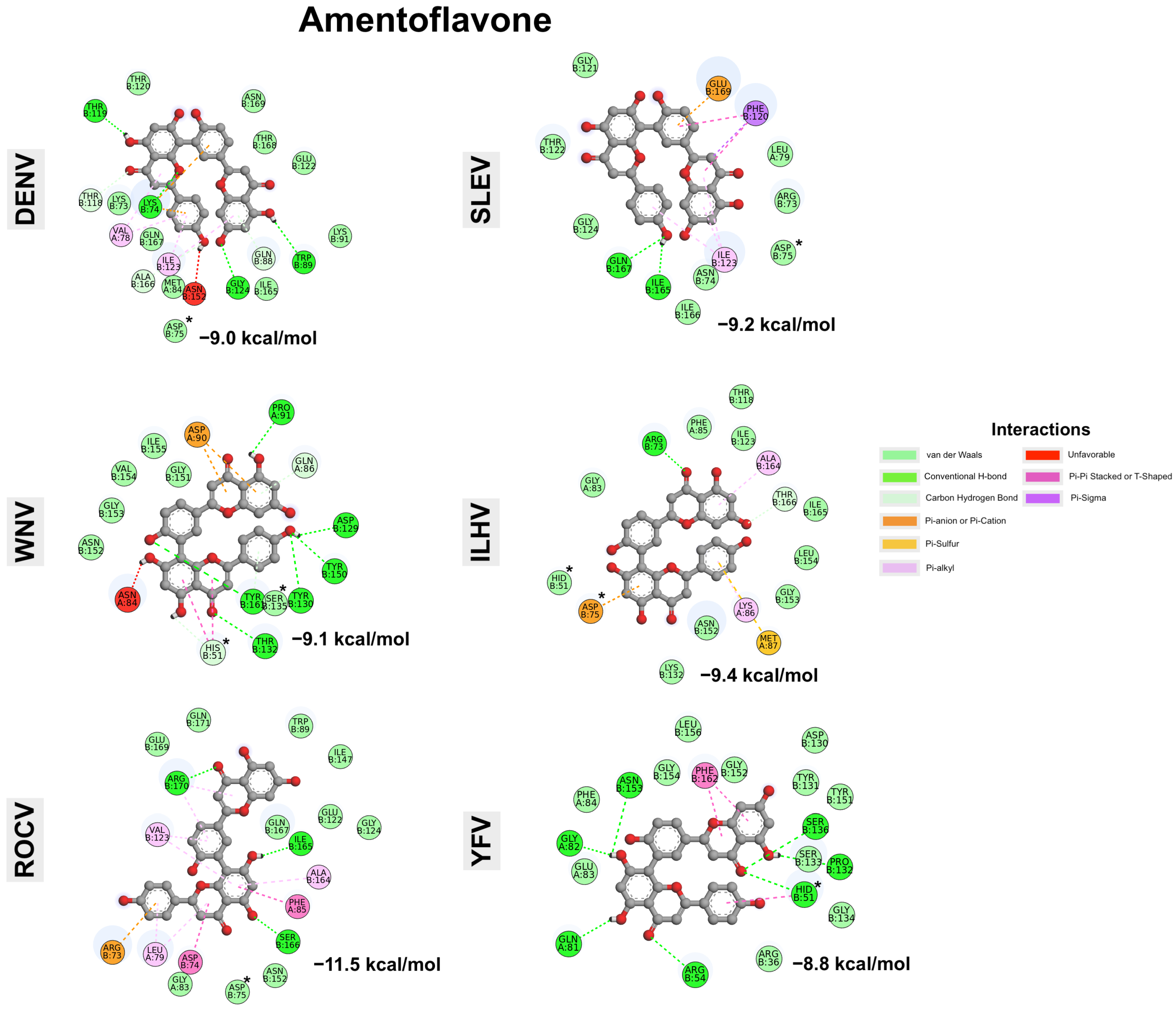
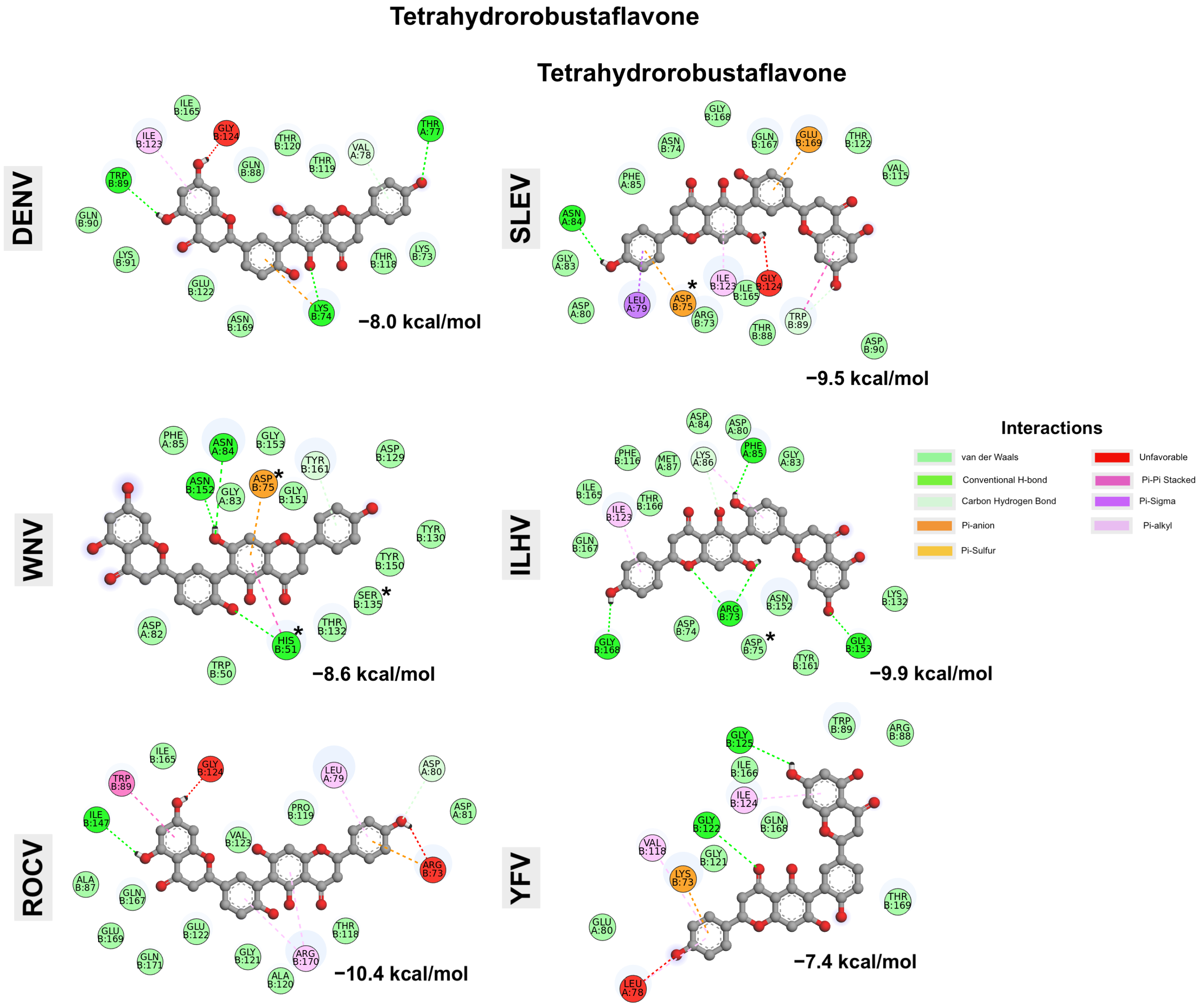
| Compounds | Energy | DENV-3 | WNV | ROCV | SLEV | ILHV | YFV | |
|---|---|---|---|---|---|---|---|---|
| Cluster 1 | Cluster 1 | Cluster 1 | Cluster 2 | |||||
| Amentoflavone | Highest | −7.4 | −6.9 | −8.5 | −7.9 | −7.9 | −7.4 | −6.8 |
| Lowest | −9 | −9.1 | −11.5 | −9.2 | −9.4 | −9.0 | −8.8 | |
| Tetrahydrorobustaflavone | Highest | −6.7 | −3.8 | −7.5 | −8.0 | −9.3 | −7.4 | −2.6 |
| Lowest | −8 | −8.6 | −10.4 | −9.5 | −9.9 | −8.3 | −7.4 | |
| Quercetin | Highest | −5.8 | −5.9 | −7.0 | −6.4 | −6.8 | −5.9 | −6.1 |
| Lowest | −7.1 | −8.4 | −8.5 | −8.9 | −8.3 | −7.0 | −7.9 | |
| Virus | Identity | TM-Score | |
|---|---|---|---|
| DENV-3 | SLEV | 60.24% | 0.635 |
| WNV | 56.63% | 0.872 | |
| YFV | 55.75% | 0.862 | |
| ILHV | 60.45% | 0.587 */0.507 ** | |
| ROCV | 56.90% | 0.790 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saivish, M.V.; Menezes, G.d.L.; Costa, V.G.d.; Nebo, L.; Silva, G.C.D.d.; Pacca, C.C.; Marques, R.E.; Nogueira, M.L.; Da Silva, R.A. Structural Insights into Plasticity and Discovery of Flavonoid Allosteric Inhibitors of Flavivirus NS2B–NS3 Protease. Biophysica 2023, 3, 71-92. https://doi.org/10.3390/biophysica3010006
Saivish MV, Menezes GdL, Costa VGd, Nebo L, Silva GCDd, Pacca CC, Marques RE, Nogueira ML, Da Silva RA. Structural Insights into Plasticity and Discovery of Flavonoid Allosteric Inhibitors of Flavivirus NS2B–NS3 Protease. Biophysica. 2023; 3(1):71-92. https://doi.org/10.3390/biophysica3010006
Chicago/Turabian StyleSaivish, Marielena Vogel, Gabriela de Lima Menezes, Vivaldo Gomes da Costa, Liliane Nebo, Gislaine Celestino Dutra da Silva, Carolina Colombelli Pacca, Rafael Elias Marques, Maurício Lacerda Nogueira, and Roosevelt Alves Da Silva. 2023. "Structural Insights into Plasticity and Discovery of Flavonoid Allosteric Inhibitors of Flavivirus NS2B–NS3 Protease" Biophysica 3, no. 1: 71-92. https://doi.org/10.3390/biophysica3010006
APA StyleSaivish, M. V., Menezes, G. d. L., Costa, V. G. d., Nebo, L., Silva, G. C. D. d., Pacca, C. C., Marques, R. E., Nogueira, M. L., & Da Silva, R. A. (2023). Structural Insights into Plasticity and Discovery of Flavonoid Allosteric Inhibitors of Flavivirus NS2B–NS3 Protease. Biophysica, 3(1), 71-92. https://doi.org/10.3390/biophysica3010006