Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Community Ecological Interactions Involving Exogenous Trait Acquisition
2.1. Competition
2.1.1. Indirect Preemption
2.1.2. Indirect Territoriality
2.1.3. Indirect Overgrowth
2.1.4. Indirect Allelopathy
2.2. Mutualism
2.2.1. Neighborhood Habitat Amelioration (“Adversity Makes Strange Bedfellows”)
2.2.2. Association-Derived Heat and Cold Tolerance
2.2.3. Cooperative Shelter Building
2.3. Consumer–Resource Interactions
2.3.1. Associational Defenses (“The Enemy of My Enemy Is My Ally”)
2.3.2. Associational Offenses (“Enemies Can Have Allies, Too”)
2.4. Mixed Ecological Interactions
2.5. Implications of Exogenous Trait Acquisition for Management and Theory
3. Conclusions
Funding
Acknowledgments
Disclaimer
Conflicts of Interest
References
- McKitrick, M.C. Phylogenetic constraint in evolutionary theory: Has it any explanatory power? Ann. Rev. Ecol. Syst. 1993, 24, 307–330. [Google Scholar] [CrossRef]
- Diamond, J.M. Dirty eating for healthy living. Nat. Cell Biol. 1999, 400, 120–121. [Google Scholar] [CrossRef] [PubMed]
- Pyerin, W.; Taniguchi, H. Phosphorylation of hepatic phenobarbital-inducible cytochrome P-450. EMBO J. 1989, 8, 3003–3010. [Google Scholar] [CrossRef]
- Tanimoto, Y.; Kaneko, H.; Ohkuma, T.; Oguri, K.; Yoshimura, H. Site-selective oxidation of strychnine by phenobarbital inducible cytochrome P-450. J. Pharm. Dyn. 1991, 14, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Rampazzo, N.; Blum, W.E.H. Changes in chemistry and mineralogy of forest soils by acid rain. Water Air Soil Pollut. 1992, 61, 209–220. [Google Scholar] [CrossRef]
- Freebairn, D.; Loch, R.; Silburn, D. Chapter 9 Soil erosion and soil conservation for vertisols. Dev. Pedotransfer Funct. Soil Hydrol. 1996, 24, 303–362. [Google Scholar] [CrossRef]
- Molina, E.; Espelta, J.M.; Pino, J.; Bagaria, G.; Armenteras, D. Influence of clay licks on the diversity and structure of an Amazonian forest. Biotropica 2018, 50, 740–749. [Google Scholar] [CrossRef]
- Walsh, M.R.; Munch, S.B.; Chiba, S.; Conover, D.O. Maladaptive changes in multiple traits caused by fishing: Impediments to population recovery. Ecol. Lett. 2005, 9, 142–148. [Google Scholar] [CrossRef]
- Baird, D.J.; Rubach, M.N.; Van den Brinkt, P.J. Trait-based ecological risk assessment (TERA): The new frontier? Integr. Environ. Assess. Manag. 2009, 4, 2–3. [Google Scholar] [CrossRef]
- Baker, R.H.A.; Cannon, R.; Bartlett, P.; Barker, I. Novel strategies for assessing and managing the risks posed by invasive alien species to global crop production and biodiversity. Ann. Appl. Biol. 2005, 146, 177–191. [Google Scholar] [CrossRef]
- Tremlová, K.; Münzbergová, Z. Importance of species traits for species distribution in fragmented landscapes. Ecology 2007, 88, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Halliday, F.W.; Rohr, J.R. Measuring the shape of the biodiversity-disease relationship across systems reveals new findings and key gaps. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lips, K.R.; Reeve, J.D.; Witters, L.R. Ecological traits predicting amphibian population declines in Central America. Conserv. Biol. 2003, 17, 1078–1088. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, E.; Wells, J.A.; Declerck, F.; Metcalfe, D.J.; Catterall, C.P.; Queiroz, C.; Aubin, I.; Bonser, S.P.; Ding, Y.; Fraterrigo, J.M.; et al. Land-use intensification reduces functional redundancy and response diversity in plant communities. Ecol. Lett. 2010, 13, 76–86. [Google Scholar] [CrossRef]
- Butterfield, B.J.; Callaway, R.M. A functional comparative approach to facilitation and its context dependence. Funct. Ecol. 2013, 27, 907–917. [Google Scholar] [CrossRef]
- Morris, L.; O’Brien, A.; Natera, S.H.; Lutz, A.; Roessner, U.; Long, S.M. Structural and functional measures of marine microbial communities: An experiment to assess implications for oil spill management. Mar. Pollut. Bull. 2018, 131, 525–529. [Google Scholar] [CrossRef]
- Sun, D. Pull in and push out: Mechanisms of horizontal gene transfer in bacteria. Front. Microbiol. 2018, 9, 2154. [Google Scholar] [CrossRef]
- Dufty, A.M., Jr.; Clobert, J.; Møller, A.P. Hormones, developmental plasticity and adaptation. Trends Ecol. Evol. 2002, 17, 190–196. [Google Scholar] [CrossRef]
- Fusco, G.; Minelli, A. Phenotypic plasticity in development and evolution: Facts and concepts. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Belcourt, M. Outsourcing-the benefits and the risks. Hum. Res. Manag. Rev. 2006, 16, 269–279. [Google Scholar] [CrossRef]
- Sapp, J. Evolution by Association: A History of Symbiosis; Oxford University Press: New York, NY, USA, 1994. [Google Scholar]
- Flower, T.P.; Child, M.F.; Ridley, A.R. The ecological economics of kleptoparasitism: Pay-offs from self-foraging versus kleptoparasitism. J. Anim. Ecol. 2012, 82, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Paini, D.R.; Funderburk, J.E.; Reitz, S.R. Competitive exclusion of a worldwide invasive pest by a native. Quantifying competition between two phytophagous insects on two host plant species. J. Anim. Ecol. 2008, 77, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Van De Waal, D.B.; Verspagen, J.M.H.; Finke, J.F.; Vournazou, V.; Immers, A.K.; Kardinaal, W.E.A.; Tonk, L.; Becker, S.; Van Donk, E.; Visser, P.M.; et al. Reversal in competitive dominance of a toxic versus non-toxic cyanobacterium in response to rising CO2. ISME J. 2011, 5, 1438–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Bourlot, V.; Tully, T.; Claessen, D. Interference versus exploitative competition in the regulation of size-structured populations. Am. Nat. 2014, 184, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Wignall, V.R.; Harry, I.C.; Davies, N.L.; Kenny, S.D.; McMINN, J.K.; Ratnieks, F.L.W. Seasonal variation in exploitative competition between honeybees and bumblebees. Oecologia 2019, 192, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. On the prevalence and relative importance of interspecific competition: Evidence from field experiments. Am. Nat. 1983, 122, 661–696. [Google Scholar] [CrossRef]
- Halpern, C.B.; Lutz, J.A. Canopy closure exerts weak controls on understory dynamics: A 30-year study of overstory–understory interactions. Ecol. Monogr. 2013, 83, 221–237. [Google Scholar] [CrossRef] [Green Version]
- Oda, R. Scent marking and contact call production in ring-tailed Lemurs (Lemur catta). Folia Primatol. 1999, 70, 121–124. [Google Scholar] [CrossRef]
- Wegener, C.; Martin, B.; Didden, C.; Edmunds, P.J. Overgrowth of Caribbean octocorals by milleporid hydrocorals. Invertebr. Biol. 2017, 137, 29–37. [Google Scholar] [CrossRef]
- Prati, D.; Bossdorf, O. Allelopathic inhibition of germination by Alliaria petiolata (Brassicaceae). Am. J. Bot. 2004, 91, 285–288. [Google Scholar] [CrossRef] [Green Version]
- Samhouri, J.F.; Vance, R.R.; Forrester, G.E.; Steele, M.A. Musical chairs mortality functions: Density-dependent deaths caused by competition for unguarded refuges. Oecologia 2009, 160, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Hussain, M.; O’Neill, S.L.; Asgari, S. Wolbachia uses a host microRNA to regulate transcripts of a methyltransferase, contributing to dengue virus inhibition in Aedes aegypti. Proc. Natl. Acad. Sci. USA 2013, 110, 10276–10281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousa, W.K.; Shearer, C.; Limay-Rios, V.; Ettinger, C.L.; Eisen, J.A.; Raizada, M.N. Root-hair endophyte stacking in finger millet creates a physicochemical barrier to trap the fungal pathogen Fusarium graminearum. Nat. Microbiol. 2016, 1, 16167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, A.S.; Moffat, C.S.; Lopez-Ruiz, F.J.; Gibberd, M.R.; Hamblin, J.; Zerihun, A. Host-multi-pathogen warfare: Pathogen interactions in co-infected plants. Front. Plant Sci. 2017, 8, 1806. [Google Scholar] [CrossRef] [Green Version]
- Cézilly, F.; Perrot-Minnot, M.-J.; Rigaud, T. Cooperation and conflict in host manipulation: Interactions among macro-parasites and micro-organisms. Front. Microbiol. 2014, 5, 248. [Google Scholar] [CrossRef]
- Hilker, F.M.; Allen, L.J.S.; Bokil, V.; Briggs, C.J.; Feng, Z.; Garrett, K.A.; Gross, L.J.; Hamelin, F.; Jeger, M.J.; Manore, C.A.; et al. Modeling virus coinfection to inform management of maize lethal necrosis in Kenya. Phytopathology 2017, 107, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.R. Mosquito-borne human viral diseases: Why Aedes aegypti? Am. J. Trop. Med. Hyg. 2018, 98, 1563–1565. [Google Scholar] [CrossRef]
- Nazareth, T.; Craveiro, I.; Moutinho, A.; Seixas, G.; Gonçalves, C.; Gonçalves, L.; Teodósio, R.; Sousa, C.A. What happens when we modify mosquitoes for disease prevention? A systematic review. Emerg. Microbes Infect. 2020, 9, 348–365. [Google Scholar] [CrossRef]
- Chu, Z.; Jin, X.; Yang, B.; Zeng, Q. Buoyancy regulation of Microcystis flos-aquae during phosphorus-limited and nitrogen-limited growth. J. Plankton Res. 2007, 29, 739–745. [Google Scholar] [CrossRef] [Green Version]
- Wilde, A.; Mullineaux, C.W. Motility in cyanobacteria: Polysaccharide tracks and Type IV pilus motors. Mol. Microbiol. 2015, 98, 998–1001. [Google Scholar] [CrossRef]
- Pratt, J.R.; Rosen, B.H. Associations of species of Vorticella (Peritrichida) and Planktonic Algae. Trans. Am. Microsc. Soc. 1983, 102, 48. [Google Scholar] [CrossRef]
- Canter, H.M.; Walsby, A.; Kinsman, R.; Ibelings, B. The effect of attached vorticellids on the buoyancy of the colonial cyanobacteriumAnabaena lemmermannii. Br. Phycol. J. 1992, 27, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Chia, M.A.; Jankowiak, J.G.; Kramer, B.J.; Goleski, J.A.; Huang, I.-S.; Zimba, P.V.; Bittencourt-Oliveira, M.D.C.; Gobler, C. Succession and toxicity of microcystis and Anabaena (Dolichospermum) blooms are controlled by nutrient-dependent allelopathic interactions. Harmful Algae 2018, 74, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.G., Jr. The ecological consequences of interspecific territorial behavior in birds. Ecology 1971, 52, 414–423. [Google Scholar] [CrossRef]
- Waser, P.M. Interactions among primate species. In Primate Societies; Smuts, B.B., Cheny, D.L., Seyfarth, R.M., Wrangham, R.W., Struhsaker, T.T., Eds.; University of Chicago Press: Chicago, IL, USA, 1986; pp. 210–226. [Google Scholar]
- Stensland, E.; Angerbjörn, A.; Berggren, P. Mixed species groups in mammals. Mammal. Rev. 2003, 33, 205–223. [Google Scholar] [CrossRef]
- Jullien, M.; Thiollay, J.-M. Multi-species territoriality and dynamic of neotropical forest understory bird flocks. J. Anim. Ecol. 1998, 67, 227–252. [Google Scholar] [CrossRef]
- Bshary, R.; Grutter, A.S. Punishment and partner switching cause cooperative behaviour in a cleaning mutualism. Biol. Lett. 2005, 1, 396–399. [Google Scholar] [CrossRef]
- Weyl, E.G.; Frederickson, M.E.; Yu, D.W.; Pierce, N.E. Economic contract theory tests models of mutualism. Proc. Natl. Acad. Sci. USA 2010, 107, 15712–15716. [Google Scholar] [CrossRef] [Green Version]
- Groom, M.J. Sand-colored nighthawks parasitize the antipredator behavior of three nesting bird species. Ecology 1992, 73, 785–793. [Google Scholar] [CrossRef]
- Nolen, M.T.; Lucas, J.R. Asymmetries in mobbing behaviour and correlated intensity during predator mobbing by nuthatches, chickadees and titmice. Anim. Behav. 2009, 77, 1137–1146. [Google Scholar] [CrossRef]
- Brown, J.L. The evolution of diversity in avian territorial systems. Wilson Bull. 1964, 76, 160–169. [Google Scholar]
- Hinsch, M.; Komdeur, J. Defence, intrusion and the evolutionary stability of territoriality. J. Theor. Biol. 2010, 266, 606–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezende, E.L.; Lavabre, J.E.; Guimarães, P.R.; Jordano, P.; Bascompte, J. Non-random coextinctions in phylogenetically structured mutualistic networks. Nat. Cell Biol. 2007, 448, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.C.; Almeida-Neto, M. A simple stochastic model for complex coextinctions in mutualistic networks: Robustness decreases with connectance. Ecol. Lett. 2015, 18, 144–152. [Google Scholar] [CrossRef] [PubMed]
- LeClaire, S.; Jacob, S.; Greene, L.K.; Dubay, G.R.; Drea, C.M. Social odours covary with bacterial community in the anal secretions of wild meerkats. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Cohan, F.M. Bacterial species and speciation. Syst. Biol. 2001, 50, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Fondi, M.; Karkman, A.; Tamminen, M.V.; Bosi, E.; Virta, M.; Fani, R.; Alm, E.; McInerney, J.O. “Every gene is everywhere but the environment selects”: Global geolocalization of gene sharing in environmental samples through network analysis. Genome Biol. Evol. 2016, 8, 1388–1400. [Google Scholar] [CrossRef]
- Giller, K.E.; Witter, E.; McGrath, S.P. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem. 1998, 30, 1389–1414. [Google Scholar] [CrossRef]
- Daghrir, R.; Drogui, P. Tetracycline antibiotics in the environment: A review. Environ. Chem. Lett. 2013, 11, 209–227. [Google Scholar] [CrossRef]
- Adamo, S.A. Parasitic aphrodisiacs: Manipulation of the hosts’ behavioral defenses by sexually transmitted parasites. Integr. Comp. Biol. 2014, 54, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Sadeh, A.; Northfield, T.D.; Rosenheim, J.A. The epidemiology and evolution of parasite transmission through cannibalism. Ecology 2016, 97, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Veiga, J.P.; Polo, V.; Viñuela, J. Nest green plants as a male status signal and courtship display in the spotless starling. Ethology 2006, 112, 196–204. [Google Scholar] [CrossRef]
- Albo, M.J.; Toft, S.; Bilde, T. Female spiders ignore condition-dependent information from nuptial gift wrapping when choosing mates. Anim. Behav. 2012, 84, 907–912. [Google Scholar] [CrossRef]
- Airoldi, L. Effects of disturbance, life histories, and overgrowth on coexistence of algal crusts and turfs. Ecology 2000, 81, 798–814. [Google Scholar] [CrossRef]
- Sikkel, P.C. Effects of nest quality on male courtship and female spawning-site choice in an algal-nesting damselfish. Bull. Mar. Sci. 1995, 57, 682–689. [Google Scholar]
- Denley, D.; Metaxas, A. Effects of intrinsic and extrinsic factors on reproduction of an ecologically significant invasive bryozoan: Implications for invasion success. Mar. Biol. 2017, 164, 145. [Google Scholar] [CrossRef]
- Yoshioka, P.M. Role of planktonic and benthic factors in the population dynamics of the Bryozoan Membranipora membranacea. Ecology 1982, 63, 457–468. [Google Scholar] [CrossRef]
- Pratt, M.C.; Grason, E.W. Invasive species as a new food source: Does a nudibranch predator prefer eating an invasive bryozoan? Biol. Invasions 2006, 9, 645–655. [Google Scholar] [CrossRef]
- Green, P.T.; O’Dowd, D.J.; Abbott, K.L.; Jeffery, M.; Retallick, K.; Mac Nally, R. Invasional meltdown: Invader–invader mutualism facilitates a secondary invasion. Ecology 2011, 92, 1758–1768. [Google Scholar] [CrossRef]
- Dillon, R.J.; Vennard, C.T.; Buckling, A.; Charnley, A.K. Diversity of locust gut bacteria protects against pathogen invasion. Ecol. Lett. 2005, 8, 1291–1298. [Google Scholar] [CrossRef]
- Brown, S.P.; Le Chat, L.; Taddei, F. Evolution of virulence: Triggering host inflammation allows invading pathogens to exclude competitors. Ecol. Lett. 2007, 11, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stecher, B.; Robbiani, R.; Walker, A.W.; Westendorf, A.M.; Barthel, M.; Kremer, M.; Chaffron, S.; MacPherson, A.J.; Buer, J.; Parkhill, J.; et al. Salmonella enterica Serovar Typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol. 2007, 5, e244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoro, C.K.; Kingsley, R.A.; Connor, T.R.; Harris, S.R.; Parry, C.M.; Al-Mashhadani, M.N.; Kariuki, S.; Msefula, C.L.; Gordon, M.A.; De Pinna, E.; et al. Intracontinental spread of human invasive Salmonella Typhimurium pathovariants in sub-Saharan Africa. Nat. Genet. 2012, 44, 1215–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, C.R.; Garrone, R.; Vacelet, J. Marine sponges discriminate between food bacteria and bacterial symbionts: Electron microscope radioautography and in situ evidence. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1984, 220, 519. [Google Scholar]
- Dolenec, T.; Lojen, S.; Kniewald, G.; Dolenec, M.; Rogan, N. Nitrogen stable isotope composition as a tracer of fish farming in invertebrates Aplysina aerophoba, Balanus perforatus and Anemonia sulcata in central Adriatic. Aquaculture 2007, 262, 237–249. [Google Scholar] [CrossRef]
- Bayer, K.; Schmitt, S.; Hentschel, U. Physiology, phylogeny and in situ evidence for bacterial and archaeal nitrifiers in the marine sponge Aplysina aerophoba. Environ. Microbiol. 2008, 10, 2942–2955. [Google Scholar] [CrossRef]
- Harris, R.J.; Jenner, R.A. Evolutionary ecology of fish venom: Adaptations and consequences of evolving a venom system. Toxins 2019, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, O.J.; Nudds, T.D. Parasite-mediated competition in deer and moose: How strong is the effect of Meningeal worm on moose? Ecol. Appl. 1994, 4, 91–103. [Google Scholar] [CrossRef]
- Price, P.W.; Westoby, M.; Rice, B. Parasite-mediated competition: Some predictions and tests. Am. Nat. 1988, 131, 544–555. [Google Scholar] [CrossRef] [Green Version]
- Mlynarek, J.J.; Knee, W.; Forbes, M.R. Host phenology, geographic range size and regional occurrence explain interspecific variation in damselfly-water mite associations. Ecography 2014, 38, 670–680. [Google Scholar] [CrossRef]
- Willis, R.J. The History of Allelopathy; Springer: New York, NY, USA, 2007. [Google Scholar]
- Mangla, S.; Callaway, R.M. Exotic invasive plant accumulates native soil pathogens which inhibit native plants. J. Ecol. 2007, 96, 58–67. [Google Scholar] [CrossRef]
- Osman, R.W.; Haugsness, J.A. Mutualism among sessile invertebrates: A mediator of competition and predation. Science 1981, 211, 846–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodall, J.; Erasmus, D. Review of the status and integrated control of the invasive alien weed, Chromolaena odorata, in South Africa. Agric. Ecosyst. Environ. 1996, 56, 151–164. [Google Scholar] [CrossRef]
- Canning-Clode, J.; Souto, J.; McCann, L. First record of Celleporaria brunnea (Bryozoa: Lepraliellidae) in Portugal and in the East Atlantic. Mar. Biodivers. Rec. 2013, 6, 108. [Google Scholar] [CrossRef]
- Haeder, S.; Wirth, R.; Herz, H.; Spiteller, D. Candicidin-producing Streptomyces support leaf-cutting ants to protect their fungus garden against the pathogenic fungus Escovopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 4742–4746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otani, S.; Challinor, V.L.; Kreuzenbeck, N.B.; Kildgaard, S.; Christensen, S.K.; Larsen, L.L.M.; Aanen, D.K.; Rasmussen, S.A.; Beemelmanns, C.; Poulsen, M. Disease-free monoculture farming by fungus-growing termites. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kasson, M.T.; Wickert, K.L.; Stauder, C.M.; Macias, A.M.; Berger, M.C.; Simmons, D.R.; Short, D.P.; DeVallance, D.B.; Hulcr, J. Mutualism with aggressive wood-degrading Flavodon ambrosius (Polyporales) facilitates niche expansion and communal social structure in Ambrosiophilus ambrosia beetles. Fungal Ecol. 2016, 23, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Ranger, C.M.; Biedermann†, P.H.W.; Phuntumart, V.; Beligala, G.U.; Ghosh, S.; Palmquist, D.E.; Mueller, R.; Barnett, J.; Schultz, P.B.; Reding, M.E.; et al. Symbiont selection via alcohol benefits fungus farming by ambrosia beetles. Proc. Natl. Acad. Sci. USA 2018, 115, 4447–4452. [Google Scholar] [CrossRef] [Green Version]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Epsky, N.D. An uncertain future for American Lauraceae: A lethal threat from redbay ambrosia beetle and laurel wilt disease (a review). Am. J. Plant Sci. 2013, 4, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Ploetz, R.C.; Hulcr, J.; Wingfield, M.J.; De Beer, Z.W. Destructive tree diseases associated with ambrosia and bark beetles: Black swan events in tree pathology? Plant Dis. 2013, 97, 856–872. [Google Scholar] [CrossRef] [Green Version]
- Kusch, J.; Stremmel, M.; Breiner, H.-W.; Adams, V.; Schweikert, M.; Schmidt, H.J. The toxic symbiont Caedibacter caryophila in the cytoplasm of Paramecium novaurelia. Microb. Ecol. 2000, 40, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Schrallhammer, M. The killer trait of Paramecium and its causative agents. Palaeodiversity 2010, 3, 79–88. [Google Scholar]
- Chen, T.-T. Paramecin 34, A Killer substance produced by Paramecium bursaria. Exp. Biol. Med. 1955, 88, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.P.; Hauzy, C.; Hulot, F.D. Ingredients for protist coexistence: Competition, endosymbiosis and a pinch of biochemical interactions. J. Anim. Ecol. 2011, 81, 222–232. [Google Scholar] [CrossRef]
- Bronstein, J.L. Our current understanding of mutualism. Q. Rev. Biol. 1994, 69, 31–51. [Google Scholar] [CrossRef]
- Connor, R.C. The benefits of mutualism: A conceptual framework. Biol. Rev. 1995, 70, 427–457. [Google Scholar] [CrossRef] [Green Version]
- Stachowicz, J.J. Mutualism, facilitation, and the structure of ecological communities: Positive interactions play a critical, but underappreciated, role in ecological communities by reducing physical or biotic stresses in existing habitats and by creating new habitats on which many species depend. BioScience 2001, 51, 235–246. [Google Scholar]
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary origin of insect-Wolbachia nutritional mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef] [Green Version]
- Gammans, N.; Bullock, J.M.; Schönrogge, K. Ant benefits in a seed dispersal mutualism. Oecologia 2005, 146, 43–49. [Google Scholar] [CrossRef]
- Peoples, B.K.; Frimpong, E.A. Context-dependent outcomes in a reproductive mutualism between two freshwater fish species. Ecol. Evol. 2016, 6, 1214–1223. [Google Scholar] [CrossRef]
- Bull, L.; Fogarty, T.C. Artificial symbiogenesis. Artif. Life 1995, 2, 269–292. [Google Scholar] [CrossRef] [PubMed]
- Chapman, M.J.; Margulis, L. Morphogenesis by symbiogenesis. Int. Microbiol. 1998, 1, 319–326. [Google Scholar] [PubMed]
- Martin, W.F.; Garg, S.; Zimorski, V. Endosymbiotic theories for eukaryote origin. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corning, P.A. “The synergism hypothesis”: On the concept of synergy and its role in the evolution of complex systems. J. Soc. Evol. Syst. 1998, 21, 133–172. [Google Scholar] [CrossRef]
- Feijen, F.A.A.; Vos, R.A.; Nuytinck, J.; Merckx, V.S.F.T. Evolutionary dynamics of mycorrhizal symbiosis in land plant diversification. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kerney, R.R. Symbioses between salamander embryos and green algae. Symbiosis 2011, 54, 107–117. [Google Scholar] [CrossRef]
- Cohen, J.E.; Goldstone, A.B.; Paulsen, M.J.; Shudo, Y.; Steele, A.N.; Edwards, B.B.; Patel, J.B.; MacArthur, J.W., Jr.; Hopkins, M.S.; Burnett, C.E.; et al. An innovative biologic system for photon-powered myocardium in the ischemic heart. Sci. Adv. 2017, 3, e1603078. [Google Scholar] [CrossRef] [Green Version]
- Ewers, R. Maxilla sinus grafting with marine algae derived bone forming material: A clinical report of long-term results. J. Oral Maxillofac. Surg. 2005, 63, 1712–1723. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Holzapfel, C. Bidirectional facilitation and interference between shrubs and annuals in the Mojave Desert. Ecology 1999, 80, 1747–1761. [Google Scholar] [CrossRef]
- Holmgren, M.; Scheffer, M.; Huston, M.A. The interplay of facilitation and competition in plant communities. Ecology 1997, 78, 1966–1975. [Google Scholar] [CrossRef]
- Webb, J.S.; Givskov, M.C.; Kjelleberg, S. Bacterial biofilms: Prokaryotic adventures in multicellularity. Curr. Opin. Microbiol. 2003, 6, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Claessen, D.; Rozen, D.E.; Kuipers, O.P.; Søgaard-Andersen, L.; Van Wezel, G.P. Bacterial solutions to multicellularity: A tale of biofilms, filaments and fruiting bodies. Nat. Rev. Genet. 2014, 12, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teitzel, G.M.; Parsek, M.R. Heavy metal resistance of biofilm and planktonic Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313–2320. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Røder, H.L.; Madsen, J.S.; Bjarnsholt, T.; Sørensen, S.J.; Burmølle, M. Interspecific bacterial interactions are reflected in multispecies biofilm spatial organization. Front. Microbiol. 2016, 7, 1366. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Banerji, A.; Jahne, M.; Herrmann, M.; Brinkman, N.; Keely, S. Bringing community ecology to bear on the issue of antimicrobial resistance. Front. Microbiol. 2019, 10, 2626. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Hamayun, M.; Radhakrishnan, R.; Waqas, M.; Kang, S.-M.; Kim, Y.; Shin, J.-H.; Choo, Y.-S.; Kim, J.-G.; Lee, I.-J. Mutualistic association of Paecilomyces formosus LHL10 offers thermotolerance to Cucumis sativus. Antonie Leeuwenhoek 2011, 101, 267–279. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Imran, M.; Jan, R.; Kim, K.M.; Lee, I.-J. Synergistic association of endophytic fungi enhances Glycine max L. resilience to combined abiotic stresses: Heavy metals, high temperature and drought stress. Ind. Crop. Prod. 2020, 143, 111931. [Google Scholar] [CrossRef]
- Torabi, A.; Bonjar, G.H.S.; Abdolshahi, R.; Pournamdari, M.; Saadoun, I.; Barka, E.A. Biological control of Paecilomyces formosus, the causal agent of dieback and canker diseases of pistachio by two strains of Streptomyces misionensis. Biol. Control. 2019, 137, 104029. [Google Scholar] [CrossRef]
- Hanna, H. Black polyethylene mulch does not reduce yield of cucumbers double-cropped with tomatoes under heat stress. HortScience 2000, 35, 190–191. [Google Scholar] [CrossRef] [Green Version]
- Neelakanta, G.; Sultana, H.; Fish, D.; Anderson, J.F.; Fikrig, E. Anaplasma phagocytophilum induces Ixodes scapularis ticks to express an antifreeze glycoprotein gene that enhances their survival in the cold. J. Clin. Investig. 2010, 120, 3179–3190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loebermann, M.; Fingerle, V.; Lademann, M.; Fritzsche, C.; Reisinger, E.C. Borrelia burgdorferi and Anaplasma phagocytophilum coinfection. Emerg. Infect. Dis. 2006, 12, 353–355. [Google Scholar] [CrossRef]
- Ruf, T.; Bieber, C. Use of social thermoregulation fluctuates with mast seeding and reproduction in a pulsed resource consumer. Oecologia 2020, 192, 919–928. [Google Scholar] [CrossRef] [Green Version]
- Cook, A. Huddling and the control of water loss by the slug Limax pseudoflavus Evans. Anim. Behav. 1981, 29, 289–298. [Google Scholar] [CrossRef]
- Eiserer, L.A. Communal roosting in birds. Bird Behav. 1984, 5, 61–80. [Google Scholar]
- Waterman, J.M.; Roth, J.D. Interspecific associations of Cape ground squirrels with two mongoose species: Benefit or cost? Behav. Ecol. Sociobiol. 2007, 61, 1675–1683. [Google Scholar] [CrossRef]
- Ancillotto, L.; Allegrini, C.; Serangeli, M.T.; Jones, G.; Russo, D. Sociality across species: Spatial proximity of newborn bats promotes heterospecific social bonding. Behav. Ecol. 2014, 26, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.N.; Bowman, J.; Burness, G. Social thermoregulation does not explain heterospecific nesting in North American flying squirrels. Biol. J. Linn. Soc. 2018, 123, 805–813. [Google Scholar] [CrossRef]
- Egley, R.L.; Breed, M.D. The fanner honey bee: Behavioral variability and environmental cues in workers performing a specialized task. J. Insect Behav. 2012, 26, 238–245. [Google Scholar] [CrossRef]
- Briscoe, N.J.; Handasyde, K.A.; Griffiths, S.R.; Porter, W.P.; Krockenberger, A.; Kearney, M.R. Tree-hugging koalas demonstrate a novel thermoregulatory mechanism for arboreal mammals. Biol. Lett. 2014, 10, 20140235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.-X.; Tan, K.; Radloff, S.E.; Phiancharoen, M.; Hepburn, H.R. Comb construction in mixed-species colonies of honeybees, Apis cerana and Apis mellifera. J. Exp. Biol. 2010, 213, 1659–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonhardt, S.D.; Baumann, A.-M.; Wallace, H.M.; Brooks, P.; Schmitt, T. The chemistry of an unusual seed dispersal mutualism: Bees use a complex set of olfactory cues to find their partner. Anim. Behav. 2014, 98, 41–51. [Google Scholar] [CrossRef]
- Chen, Y.P.; Pettis, J.S.; Corona, M.; Chen, W.P.; Li, C.J.; Spivak, M.; Visscher, P.K.; DeGrandi-Hoffman, G.; Boncristiani, H.; Zhao, Y.; et al. Israeli acute paralysis virus: Epidemiology, pathogenesis and implications for honey bee health. PLoS Pathog. 2014, 10, e1004261. [Google Scholar] [CrossRef]
- Pegg, G.S.; Webb, R.I.; Carnegie, A.J.; Wingfield, M.J.; Drenth, A. Infection and disease development of Quambalaria spp. on Corymbia and Eucalyptus species. Plant Pathol. 2009, 58, 642–654. [Google Scholar] [CrossRef] [Green Version]
- Meyers, M.A.; Lin, Y.S.; Olevsky, E.; Chen, P.-Y. Battle in the Amazon: Arapaima versus Piranha. Adv. Eng. Mater. 2012, 14, B279–B288. [Google Scholar] [CrossRef]
- Schuster, S. Hunting in archerfish-An ecological perspective on a remarkable combination of skills. J. Exp. Biol. 2018, 221, jeb159723. [Google Scholar] [CrossRef] [Green Version]
- Nelson, X.J.; Jackson, R.R. Aggressive use of Batesian mimicry by an ant-like jumping spider. Biol. Lett. 2009, 5, 755–757. [Google Scholar] [CrossRef] [Green Version]
- Cheney, K.L. Multiple selective pressures apply to a coral reef fish mimic: A case of Batesian–aggressive mimicry. Proc. R. Soc. B Biol. Sci. 2010, 277, 1849–1855. [Google Scholar] [CrossRef] [Green Version]
- Mougi, A.; Kishida, O.; Iwasa, Y. Coevolution of phenotypic plasticity in predator and prey: Why are inducible offenses rarer than inducible defenses? Evolution 2010, 65, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Banerji, A.; Morin, P.J. Trait-mediated apparent competition in an intraguild predator-prey system. Oikos 2013, 123, 567–574. [Google Scholar] [CrossRef]
- Campbell, S.; Harada, R.; DeFelice, S.; Bienfang, P.K.; Li, Q. Bacterial production of tetrodotoxin in the pufferfish Arothron hispidus. Nat. Prod. Res. 2009, 23, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.F.; Arakawa, O.; Saito, T.; Noguchi, T.; Simidu, U.; Tsukamoto, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin-producing bacteria from the blue-ringed octopus Octopus maculosus. Mar. Biol. 1989, 100, 327–332. [Google Scholar] [CrossRef]
- Gochfeld, D.J. Territorial damselfishes facilitate survival of corals by providing an associational defense against predators. Mar. Ecol. Prog. Ser. 2010, 398, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.B.; Fong, P.; Kennison, R.; Smith, J. Spatial refuges and associational defenses promote harmful blooms of the alga Caulerpa sertularioides onto coral reefs. Oecologia 2010, 164, 1039–1048. [Google Scholar] [CrossRef]
- Steinke, M.; Malin, G.; Liss, P.S. Trophic interactions in the sea: An ecological role for climate relevant volatiles?1. J. Phycol. 2002, 38, 630–638. [Google Scholar] [CrossRef]
- Yuan, J.S.; Himanen, S.J.; Holopainen, J.K.; Chen, F.; Stewart, C.N., Jr. Smelling global climate change: Mitigation of function for plant volatile organic compounds. Trends Ecol. Evol. 2009, 24, 323–331. [Google Scholar] [CrossRef]
- Hay, M.E. Associational plant defenses and the maintenance of species diversity: Turning competitors into accomplices. Am. Nat. 1986, 128, 617–641. [Google Scholar] [CrossRef]
- Clay, K.; Holah, J. Fungal endophyte symbiosis and plant diversity in successional fields. Science 1999, 285, 1742–1744. [Google Scholar] [CrossRef]
- Rudgers, J.A.; Mattingly, W.B.; Koslow, J.M. Mutualistic fungus promotes plant invasion into diverse communities. Oecologia 2005, 144, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Rai, B.; Freedman, H.; Addicott, J.F. Analysis of three species models of mutualism in predator-prey and competitive systems. Math. Biosci. 1983, 65, 13–50. [Google Scholar] [CrossRef]
- Addicott, J.F.; Freedman, H. On the structure and stability of mutualistic systems: Analysis of predator-prey and competition models as modified by the action of a slow-growing mutualist. Theor. Popul. Biol. 1984, 26, 320–339. [Google Scholar] [CrossRef]
- van Rijn, P.C.J.; van Houten, Y.M.; Sabelis, M.W. How plants benefit from providing food to predators even when it is also edible to herbivores. Ecology 2002, 83, 2664–2679. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Janssen, A.; Kant, M.R. ECOLOGY: Enhanced: The enemy of my enemy is my ally. Science 2001, 291, 2104–2105. [Google Scholar] [CrossRef] [Green Version]
- Hay, M.E.; Parker, J.D.; Burkepile, D.E.; Caudill, C.C.; Wilson, A.E.; Hallinan, Z.P.; Chequer, A.D. Mutualisms and aquatic community structure: The enemy of my enemy is my friend. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 175–197. [Google Scholar] [CrossRef] [Green Version]
- Nishida, R. Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol. 2002, 47, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.L.; Brodie, E.D., Jr.; Brodie, E.D., III. A resistant predator and its toxic prey: Persistence of newt toxin leads to poisonous (not venomous) snakes. J. Chem. Ecol. 2004, 30, 1901–1919. [Google Scholar] [CrossRef]
- Bush, S.E.; Clayton, D.H. Anti-parasite behaviour of birds. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170196. [Google Scholar] [CrossRef]
- Noë, R.; Bshary, R. The formation of red colobus-Diana monkey associations under predation pressure from chimpanzees. Proc. Biol. Sci. 1997, 264, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Semeniuk, C.A.D.; Dill, L.M. Anti-predator benefits of mixed-species groups of Cowtail Stingrays (Pastinachus sephen) and whiprays (Himantura uarnak) at rest. Ethology 2006, 112, 33–43. [Google Scholar] [CrossRef]
- Hetrick, S.A.; Sieving, K.E. Antipredator calls of tufted titmice and interspecific transfer of encoded threat information. Behav. Ecol. 2011, 23, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Marcus, M.J. Feeding associations between capybaras and jacanas: A case of interspecific grooming and possibly mutualism. Ibis 2008, 127, 240–243. [Google Scholar] [CrossRef]
- Cote, I.M. Evolution and ecology of cleaning symbioses in the sea. Oceanogr. Mar. Biol. 2000, 38, 311–355. [Google Scholar]
- Frederickson, M.E. Ant species confer different partner benefits on two neotropical myrmecophytes. Oecologia 2005, 143, 387–395. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Janssen, A.; Takabayashi, J. Can plants evolve stable alliances with the enemies’ enemies? J. Plant Interact. 2011, 6, 71–75. [Google Scholar] [CrossRef]
- Kessler, A.; Baldwin, I.-T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef]
- Cusick, K.D.; Widder, E.A. Intensity differences in bioluminescent dinoflagellates impact foraging efficiency in a nocturnal predator. Bull. Mar. Sci. 2014, 90, 797–811. [Google Scholar] [CrossRef]
- Verpoorte, R. Secondary metabolism. In Metabolic Engineering of Plant Secondary Metabolism; Verpoorte, R., Alfermann, A.W., Eds.; Springer: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Valiadi, M.; Iglesias-Rodriguez, D. Understanding bioluminescence in dinoflagellates-How far have we come? Microorganisms 2013, 1, 3–25. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.; Rodriguez-Saona, C.; Del Conte, S.C.C. Methyl jasmonate induction of cotton: A field test of the ‘attract and reward’ strategy of conservation biological control. AoB Plants 2017, 9, 032. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, N.; Barber, P.H.; Weisz, J.B. Episymbiotic microbes as food and defence for marine isopods: Unique symbioses in a hostile environment. Proc. Biol. Sci. 2005, 272, 1209–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proud, D.N.; Wade, R.R.; Rock, P.; Townsend, V.R., Jr.; Jiménez Chavez, D.J. Epizoic cyanobacteria associated with a Neotropical harvestman (Opiliones: Sclerosomatidae) from Costa Rica. J. Arachnol. 2012, 40, 259–261. [Google Scholar] [CrossRef]
- Vance, R.R. A mutualistic interaction between a sessile marine clam and its epibionts. Ecology 1978, 59, 679–685. [Google Scholar] [CrossRef]
- Maldonado, M.; Uriz, M.J. Relationships between sponges and crabs: Patterns of epibiosis on Inachus aguiarii (Decapoda: Majidae). Mar. Biol. 1992, 113, 281–286. [Google Scholar]
- Suutari, M.; Majaneva, M.; Fewer, D.P.; Voirin, B.; Aiello, A.; Friedl, T.; Chiarello, A.G.; Blomster, J. Molecular evidence for a diverse green algal community growing in the hair of sloths and a specific association with Trichophilus welckeri (Chlorophyta, Ulvophyceae). BMC Evol. Biol. 2010, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, A.G. The role of the extrafloral nectaries of catalpa speciosa in limiting herbivory and increasing fruit production. Ecology 1982, 63, 663–669. [Google Scholar] [CrossRef]
- Chanam, J.; Borges, R.M. Cauline domatia of the ant-plant Humboldtia brunonis (Fabaceae). Flora Morphol. Distrib. Funct. Ecol. Plants 2017, 236, 58–66. [Google Scholar] [CrossRef]
- Fonseca-Romero, M.A.; Fornoni, J.; Del-Val, E.; Boege, K. Ontogenetic trajectories of direct and indirect defenses of myrmecophytic plants colonized either by mutualistic or opportunistic ant species. Oecologia 2019, 190, 857–865. [Google Scholar] [CrossRef]
- Stiefel, K.M.; Reyes, R.B., Jr. The marine goby-Alpheid shrimp symbiosis does not correlate with larger fish eye size. BioRxiv 2018. [Google Scholar] [CrossRef]
- Mouritsen, K.N.; Poulin, R. The mud flat anemone-cockle association: Mutualism in the intertidal zone? Oecologia 2003, 135, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Lafferty, K.D. The evolution of trophic transmission. Parasitol. Today 1999, 15, 111–115. [Google Scholar] [CrossRef]
- Fenton, A.; Rands, S.A. The impact of parasite manipulation and predator foraging behavior on predator-prey communities. Ecology 2006, 87, 2832–2841. [Google Scholar] [CrossRef]
- Schneider, D.S.; James, A.A. Bridging the gaps in vector biology. EMBO Rep. 2006, 7, 259–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, Y.; Jang, S.; Wang, H. Cooperative hunting in a discrete predator–prey system II. J. Biol. Dyn. 2018, 13, 247–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinsted, L.; Schou, M.F.; Settepani, V.; Holm, C.; Bird, T.L.; Bilde, T. Prey to predator body size ratio in the evolution of cooperative hunting-A social spider test case. Dev. Genes Evol. 2019, 230, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Mlot, C. A Soil Story. Sci. News 1997, 152, 58–59. [Google Scholar] [CrossRef]
- Chung, S.H.; Rosa, C.; Scully, E.D.; Peiffer, M.; Tooker, J.F.; Hoover, K.; Luthe, D.S.; Felton, G.W. Herbivore exploits orally secreted bacteria to suppress plant defenses. Proc. Natl. Acad. Sci. USA 2013, 110, 15728–15733. [Google Scholar] [CrossRef] [Green Version]
- Shiga, D. Poisonous partnership: Parasitoid wasps use viruses as a weapon. Sci. News 2005, 167, 136–137. [Google Scholar] [CrossRef]
- Dowd, P.F.; Shen, S.K. The contribution of symbiotic yeast to toxin resistance of the cigarette beetle (Lasioderma serricorne). Entomol. Exp. Appl. 1990, 56, 241–248. [Google Scholar] [CrossRef]
- Kohl, K.D.; Connelly, J.W.; Dearing, M.D.; Forbey, J.S. Microbial detoxification in the gut of a specialist avian herbivore, the Greater Sage-Grouse. FEMS Microbiol. Lett. 2016, 363, 144. [Google Scholar] [CrossRef] [Green Version]
- Hulet, W.H.; Musil, G. Intracellular bacteria in the light organ of the deep sea angler fish, Melanocetus murrayi. Copeia 2006, 1968, 506. [Google Scholar] [CrossRef]
- Widder, E.A. A predatory use of counterillumination by the squaloid shark, Isistius brasiliensis. Environ. Boil. Fishes 1998, 53, 267–273. [Google Scholar] [CrossRef]
- Jones, B.W.; Nishiguchi, M.K. Counterillumination in the Hawaiian bobtail squid, Euprymna scolopes Berry (Mollusca: Cephalopoda). Mar. Biol. 2004, 144, 1151–1155. [Google Scholar] [CrossRef]
- Herring, P.J.; Cope, C. Red bioluminescence in fishes: On the suborbital photophores of Malacosteus, Pachystomias and Aristostomias. Mar. Biol. 2005, 148, 383–394. [Google Scholar] [CrossRef]
- Sutton, T. Trophic ecology of the deep-sea fish Malacosteus niger (Pisces: Stomiidae): An enigmatic feeding ecology to facilitate a unique visual system? Deep. Sea Res. Part I Oceanogr. Res. Pap. 2005, 52, 2065–2076. [Google Scholar] [CrossRef] [Green Version]
- Dean, W.R.J.; Siegfried, W.R.; Macdonald, I.A.W. The fallacy, fact, and fate of guiding behavior in the greater honeyguide. Conserv. Biol. 1990, 4, 99–101. [Google Scholar] [CrossRef]
- Minta, S.C.; Minta, K.A.; Lott, D.F. Hunting associations between badgers (Taxidea taxus) and coyotes (Canis latrans). J. Mammal. 1992, 73, 814–820. [Google Scholar] [CrossRef]
- Bshary, R.; Hohner, A.; Ait-El-Djoudi, K.; Fricke, H. Interspecific communicative and coordinated hunting between groupers and giant moray eels in the Red Sea. PLoS Biol. 2006, 4, e431. [Google Scholar] [CrossRef]
- D’Angelo, G.B.; Sazima, I. Commensal association of piscivorous birds with foraging otters in southeastern Brazil, and a comparison with such a relationship of piscivorous birds with cormorants. J. Nat. Hist. 2013, 48, 241–249. [Google Scholar] [CrossRef]
- Lönnstedt, O.M.; Ferrari, M.C.O.; Chivers, D.P. Lionfish predators use flared fin displays to initiate cooperative hunting. Biol. Lett. 2014, 10, 20140281. [Google Scholar] [CrossRef]
- Sridhar, H.; Guttal, V. Friendship across species borders: Factors that facilitate and constrain heterospecific sociality. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170014. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.S.; Knollenberg, W.G. Adaptive indirect effects: The fitness of burying beetles with and without their phoretic mites. Evol. Ecol. 1987, 1, 139–159. [Google Scholar] [CrossRef]
- Krishnan, A.; Muralidharan, S.; Sharma, L.; Borges, R.M. A hitchhiker’s guide to a crowded syconium: How do fig nematodes find the right ride? Funct. Ecol. 2010, 24, 741–749. [Google Scholar] [CrossRef]
- Badets, M.; Du Preez, L. Phoretic interaction between the kangaroo leech Marsupiobdella africana (Hirudinea: Glossiphoniidae) and the cape river crab Potamonautes perlatus (Decapoda: Potamonautidae). Int. J. Parasitol. Parasites Wildl. 2014, 3, 6–11. [Google Scholar] [CrossRef] [Green Version]
- McLeod, G.; Gries, R.; Von Reuss, S.H.; Rahe, J.E.; McIntosh, R.; König, W.A.; Gries, G. The pathogen causing Dutch elm disease makes host trees attract insect vectors. Proc. Biol. Sci. 2005, 272, 2499–2503. [Google Scholar] [CrossRef]
- Robinson, A.; Busula, A.O.; Voets, M.A.; Beshir, K.B.; Caulfield, J.C.; Powers, S.J.; Verhulst, N.O.; Winskill, P.; Muwanguzi, J.; Birkett, M.A.; et al. Plasmodium-associated changes in human odor attract mosquitoes. Proc. Natl. Acad. Sci. USA 2018, 115, E4209–E4218. [Google Scholar] [CrossRef] [Green Version]
- de Vega, C.; Arista, M.; Ortiz, P.L.; Talavera, S. Anatomical relations among endophytic holoparasitic angiosperms, autotrophic host plants and mycorrhizal fungi: A novel tripartite interaction. Am. J. Bot. 2010, 97, 730–737. [Google Scholar] [CrossRef]
- Anderson, B. Adaptations to foliar absorption of faeces: A pathway in plant carnivory. Ann. Bot. 2005, 95, 757–761. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.C. Tremoctopus violaceus uses physalia tentacles as weapons. Science 1963, 139, 764–766. [Google Scholar] [CrossRef]
- Tauber, C.A.; De León, T.; Penny, N.D.; Tauber, M.J. The Genus Ceraeochrysa (Neuroptera: Chrysopidae) of America north of Mexico: Larvae, adults, and comparative biology. Ann. Entomol. Soc. Am. 2000, 93, 1195–1221. [Google Scholar] [CrossRef]
- Robinson, S.K. Use of bait and lures by green-backed herons in Amazonian Peru. Wilson Bull. 1994, 106, 567–569. [Google Scholar]
- Krützen, M.; Kreicker, S.; MacLeod, C.D.; Learmonth, J.; Kopps, A.M.; Walsham, P.; Allen, S.J. Cultural transmission of tool use by Indo-Pacific bottlenose dolphins (Tursiops sp.) provides access to a novel foraging niche. Proc. Biol. Sci. 2014, 281, 20140374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefèvre, T.; Lebarbenchon, C.; Gauthier-Clerc, M.; Missé, D.; Poulin, R.; Thomas, F. The ecological significance of manipulative parasites. Trends Ecol. Evol. 2009, 24, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R. Chapter 5-Parasite manipulation of host behavior: An update and frequently asked questions. Adv. Stud. Behav. 2010, 41, 151–186. [Google Scholar]
- Nedergaard, O.A. Curare: The flying death. Pharmacol. Toxicol. 2003, 92, 154–155. [Google Scholar] [CrossRef]
- Myers, C.W.; Daly, J.W.; Malkin, B. A dangerously toxic new frog (Phyllobates) used by Emberá Indians of western Colombia, with discussion of blowgun fabrication and dart poisoning. Bull. Am. Mus. Nat. Hist. 1978, 161. Available online: http://hdl.handle.net/2246/1286 (accessed on 6 January 2021).
- Spottiswoode, C.N.; Begg, K.S.; Begg, C.M. Reciprocal signaling in honeyguide-human mutualism. Science 2016, 353, 387–389. [Google Scholar] [CrossRef] [Green Version]
- Koster, J. The impact of hunting with dogs on wildlife harvests in the Bosawas Reserve, Nicaragua. Environ. Conserv. 2008, 35, 211–220. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M.J.; Surovell, T.A. Spatial expression of kinship among the Dukha reindeer herders of Northern Mongolia. Arct. Anthr. 2017, 54, 110–119. [Google Scholar] [CrossRef]
- Park, K.J.; Lee, M.J.; Kang, H.; Kim, K.S.; Lee, S.-H.; Cho, I.; Lee, H.H. Saeng-Maek-San, a medicinal herb complex, protects liver cell damage induced by alcohol. Biol. Pharm. Bull. 2002, 25, 1451–1455. [Google Scholar] [CrossRef] [Green Version]
- Jaeschke, H.; Williams, C.D.; McGill, M.R.; Farhood, A. Herbal extracts as hepatoprotectants against acetaminophen hepatotoxicity. World J. Gastroenterol. 2010, 16, 2448–2450. [Google Scholar] [CrossRef] [PubMed]
- Sherman, P.W.; Flaxman, S.M. Protecting ourselves from food: Spices and morning sickness may shield us from toxins and microorganisms in the diet. Am. Sci. 2001, 89, 142–151. [Google Scholar] [CrossRef]
- Marini, E.; Magi, G.; Mingoia, M.; Pugnaloni, A.; Facinelli, B. Antimicrobial and anti-virulence activity of capsaicin against erythromycin-resistant, cell-invasive group a Streptococci. Front. Microbiol. 2015, 6, 1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lettieri, J.T.; Dain, B. Effects of beano on the tolerability and pharmacodynamics of acarbose. Clin. Ther. 1998, 20, 497–504. [Google Scholar] [CrossRef]
- Czerucka, D.; Piche, T.; Rampal, P. Review article: Yeast as probiotics-Saccharomyces boulardii. Aliment. Pharmacol. Ther. 2007, 26, 767–778. [Google Scholar] [CrossRef]
- Burton, K.J.; Rosikiewicz, M.; Pimentel, G.; Bütikofer, U.; von Ah, U.; Voirol, M.-J.; Croxatto, A.; Aeby, S.; Drai, J.; McTernan, P.G.; et al. Probiotic yogurt and acidified milk similarly reduce postprandial inflammation and both alter the gut microbiota of healthy, young men. Br. J. Nutr. 2017, 117, 1312–1322. [Google Scholar] [CrossRef] [Green Version]
- Beukemaj, J.J. Acquired hook-avoidance in the pike Esox lucius L. fished with artificial and natural baits. J. Fish Biol. 1970, 2, 155–160. [Google Scholar] [CrossRef]
- Drake, D.A.R.; Mandrak, N.E. Ecological risk of live bait fisheries: A new angle on selective fishing. Fisheries 2014, 39, 201–211. [Google Scholar] [CrossRef]
- Daura-Jorge, F.G.; Cantor, M.; Ingram, S.N.; Lusseau, S.M.; Simões-Lopes, P.C. The structure of a bottlenose dolphin society is coupled to a unique foraging cooperation with artisanal fishermen. Biol. Lett. 2012, 8, 702–705. [Google Scholar] [CrossRef] [Green Version]
- Talou, T.; Gaset, A.; Delmas, M.; Kulifaj, M.; Montant, C. Dimethyl sulphide: The secret for black truffle hunting by animals? Mycol. Res. 1990, 94, 277–278. [Google Scholar] [CrossRef]
- Ruslin, F.; Azmi, M.A.; Matsuda, I.; Amir, R.; Md-Zain, B.M. Monkey school: Training phases for coconut-picking macaques (Macaca nemestrina). Malay. Nat. J. 2017, 69, 301–306. [Google Scholar]
- Marcone, M. Composition and properties of Indonesian palm civet coffee (Kopi Luwak) and Ethiopian civet coffee. Food Res. Int. 2004, 37, 901–912. [Google Scholar] [CrossRef]
- Pasha, C.; Reddy, G. Nutritional and medicinal improvement of black tea by yeast fermentation. Food Chem. 2005, 89, 449–453. [Google Scholar] [CrossRef]
- Schwan, R.F.; Wheals, A.E. The microbiology of cocoa fermentation and its role in chocolate quality. Crit. Rev. Food Sci. Nutr. 2004, 44, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M. Yeasts in botrytized wine making. In Yeasts in the Production of Wine; Romano, P., Ciani, M., Fleet, G., Eds.; Springer: New York, NY, USA, 2019. [Google Scholar]
- Zeckhauser, R.J. Human hunters and nonhuman predators: Fundamental differences. Proc. Natl. Acad. Sci. USA 2016, 114, 13–14. [Google Scholar] [CrossRef] [Green Version]
- Dicke, M.; Sabelis, M.W. Infochemical terminology: Based on cost-benefit analysis rather than origin of compounds? Funct. Ecol. 1988, 2, 131. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.C.; Di Bitetti, M.S.; Janson, C.; Slobodkin, L.B.; Valenzuela, N. An “Audience effect” for ecological terminology: Use and misuse of jargon. Oikos 1997, 80, 632. [Google Scholar] [CrossRef] [Green Version]
- Queller, D.C.; Strassmann, J.E. Problems of multi-species organisms: Endosymbionts to holobionts. Biol. Philos. 2016, 31, 855–873. [Google Scholar] [CrossRef] [Green Version]
- Michálek, O.; Kuhn-Nentwig, L.; Natarajan, R. High specific efficiency of venom of two prey-specialized spiders. Toxins 2019, 11, 687. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Shokal, U.; Forst, S.; Eleftherianos, I. An improved method for generating axenic entomopathogenic nematodes. BMC Res. Notes 2015, 8, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Persoons, A.; Hayden, K.J.; Fabre, B.; Frey, P.; De Mita, S.; Tellier, A.; Halkett, F. The escalatory Red Queen: Population extinction and replacement following arms race dynamics in poplar rust. Mol. Ecol. 2017, 26, 1902–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strotz, L.C.; Simões, M.; Girard, M.G.; Breitkreuz, L.; Kimmig, J.; Lieberman, B.S. Getting somewhere with the Red Queen: Chasing a biologically modern definition of the hypothesis. Biol. Lett. 2018, 14, 20170734. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Atamian, H.S.; Shen, Z.; Briggs, S.P.; Kaloshian, I. GroEL from the endosymbiont Buchnera aphidicola betrays the aphid by triggering plant defense. Proc. Natl. Acad. Sci. USA 2014, 111, 8919–8924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronek, M.; Sajnaga, E.; Pleszczyńska, M.; Kazimierczak, W.; Lis, M.; Wiater, A. Bacteria from the midgut of common cockchafer (Melolontha melolontha L.) larvae exhibiting antagonistic activity against bacterial symbionts of entomopathogenic nematodes: Isolation and molecular identification. Int. J. Mol. Sci. 2020, 21, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polis, G.A.; Sears, A.L.; Huxel, G.R.; Strong, D.R.; Maron, J. When is a trophic cascade a trophic cascade? Trends Ecol. Evol. 2000, 15, 473–475. [Google Scholar] [CrossRef]
- Fortin, D.; Beyer, H.L.; Boyce, M.S.; Smith, D.W.; Duchesne, T.; Mao, J.S. Wolves influence elk movements: Behavior shapes a trophic cascade in Yellowstone National Park. Ecology 2005, 86, 1320–1330. [Google Scholar] [CrossRef] [Green Version]
- Berger, J. Anthropogenic extinction of top carnivores and interspecific animal behaviour: Implications of the rapid decoupling of a web involving wolves, bears, moose and ravens. Proc. Biol. Sci. 1999, 266, 2261–2267. [Google Scholar] [CrossRef] [Green Version]
- Stahler, D.; Heinrich, B.; Smith, D. Common ravens, Corvus corax, preferentially associate with grey wolves, Canis lupus, as a foraging strategy in winter. Anim. Behav. 2002, 64, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Despret, V. The enigma of the raven. Angelaki 2015, 20, 57–72. [Google Scholar] [CrossRef]
- Walker, L.E.; Marzluff, J.M.; Metz, M.C.; Wirsing, A.J.; Moskal, L.M.; Stahler, D.R.; Smith, D.W. Population responses of common ravens to reintroduced gray wolves. Ecol. Evol. 2018, 8, 11158–11168. [Google Scholar] [CrossRef] [Green Version]
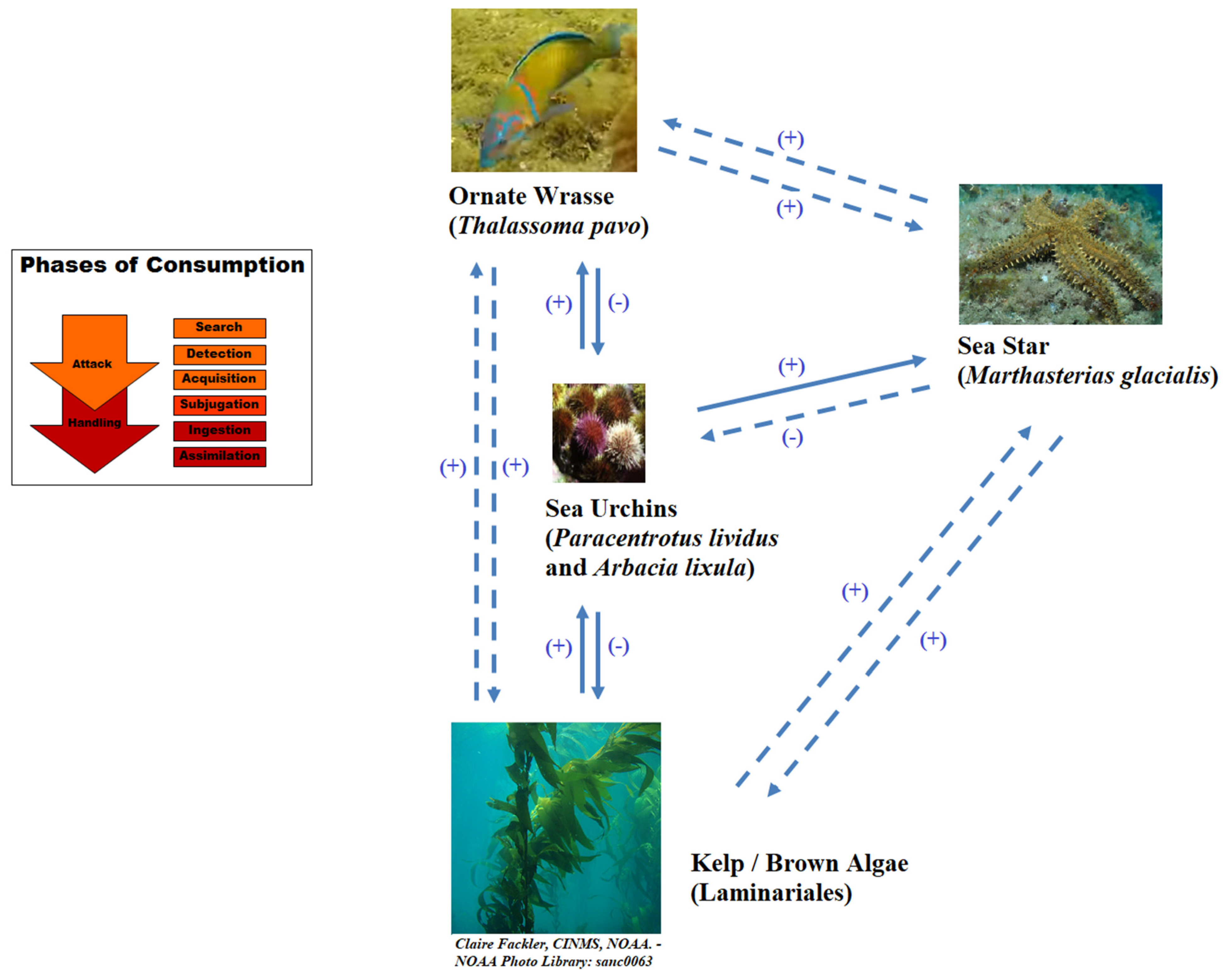
- Galasso, N.M.; Bonaviri, C.; Di Trapani, F.; Picciotto, M.; Gianguzza, P.; Agnetta, D.; Badalamenti, F. Fish-seastar facilitation leads to algal forest restoration on protected rocky reefs. Sci. Rep. 2015, 5, srep12409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myerscough, M.R.; Darwen, M.; Hogarth, W. Stability, persistence and structural stability in a classical predator-prey model. Ecol. Model. 1996, 89, 31–42. [Google Scholar] [CrossRef]
- Samways, M.J. Asymmetrical competition and amensalism through soil dumping by the ant, Myrmicaria natalensis. Ecol. Entomol. 1983, 8, 191–194. [Google Scholar] [CrossRef]
- Hulme-Beaman, A.; Dobney, K.; Cucchi, T.; Searle, J.B. An ecological and evolutionary framework for commensalism in anthropogenic environments. Trends Ecol. Evol. 2016, 31, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Leung, T.L.F.; Poulin, R. Parasitism, commensalism, and mutualism: Exploring the many shades of symbioses. Life Environ. 2008, 58, 107–115. [Google Scholar]
- Dutton, E.M.; Luo, E.Y.; Cembrowski, A.R.; Shore, J.S.; Frederickson, M.E. Three’s a crowd: Trade-offs between attracting pollinators and ant bodyguards with nectar rewards in Turnera. Am. Nat. 2016, 188, 38–51. [Google Scholar] [CrossRef]
- Shankar, U. Predation risk of pollinators on flowers. Bee World 2015, 92, 84–85. [Google Scholar] [CrossRef]
- Webster, M.S. Interspecific brood parasitism of Montezuma oropendolas by giant cowbirds: Parasitism or mutualism? Condor 1994, 96, 794–798. [Google Scholar] [CrossRef]
- Greenwood, P.G. Acquisition and use of nematocysts by cnidarian predators. Toxicon 2009, 54, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Janzen, D.H. Allelopathy by myrmecophytes: The ant azteca as an allelopathic agent of cecropia. Ecology 2006, 50, 147–153. [Google Scholar] [CrossRef]
- Anderson, B.; Midgley, J.J.; Stewart, B.A. Facilitated selfing offers reproductive assurance: A mutualism between a hemipteran and carnivorous plant. Am. J. Bot. 2003, 90, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Hountondji, F.C.; Sabelis, M.W.; Hanna, R.; Janssen, A. Herbivore-induced plant volatiles trigger sporulation in entomopathogenic fungi: The case of Neozygites tanajoae infecting the cassava green mite. J. Chem. Ecol. 2005, 31, 1003–1021. [Google Scholar] [CrossRef] [PubMed]
- Loudon, A.H.; Holland, J.A.; Umile, T.P.; Burzynski, E.A.; Minbiole, K.P.C.; Harris, R.N. Interactions between amphibians’ symbiotic bacteria cause the production of emergent anti-fungal metabolites. Front. Microbiol. 2014, 5, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.C.; Garner, T.W.J. Chytrid fungi and global amphibian declines. Nat. Rev. Genet. 2020, 18, 332–343. [Google Scholar] [CrossRef]
- Saikkonen, K.; Saari, S.; Helander, M. Defensive mutualism between plants and endophytic fungi? Fungal Divers. 2010, 41, 101–113. [Google Scholar] [CrossRef]
- Karnosky, D.F. Dutch elm disease: A review of the history, environmental implications, control, and research needs. Environ. Conserv. 1979, 6, 311–322. [Google Scholar] [CrossRef]
- Wilde, S.B.; Johansen, J.R.; Wilde, H.D.; Jiang, P.; Bartelme, B.; Haynie, R.S. Aetokthonos hydrillicola gen. et sp. nov.: Epiphytic cyanobacteria on invasive aquatic plants implicated in Avian Vacuolar Myelinopathy. Phytotaxa 2014, 181, 243–260. [Google Scholar] [CrossRef] [Green Version]
- Fleming, J.-A.G. The integration of polydnavirus genomes in parasitoid genomes: Implications for biocontrol and genetic analyses of parasitoid wasps. Biol. Control. 1991, 1, 127–135. [Google Scholar] [CrossRef]
- Georgis, R.; Gaugler, R. Predictability in biological control using entomopathogenic nematodes. J. Econ. Entomol. 1991, 84, 713–720. [Google Scholar] [CrossRef]
- Orzech, K.M.; Nichter, M. From resilience to resistance: Political ecological lessons from antibiotic and pesticide resistance. Annu. Rev. Anthr. 2008, 37, 267–282. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hayatsu, M.; Hosokawa, T.; Nagayama, A.; Tago, K.; Fukatsu, T. Symbiont-mediated insecticide resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 8618–8622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Liu, Y.; Graham, R.I.; Wilson, K.; Wu, K. Densovirus is a mutualistic symbiont of a global crop pest (Helicoverpa armigera) and protects against a baculovirus and Bt biopesticide. PLoS Pathog. 2014, 10, e1004490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauldin, J.K.; Carter, F.L.; Rich, N.M. Protozoan populations of Reticulitermes flavipes (Kollar) exposed to heart-wood blocks of 21 American species. Mater. Org. 1981, 16, 15–28. [Google Scholar]
- Waller, D.A. Ampicillin, tetracycline and urea as protozoicides for symbionts of Reticulitermes flavipes and R. virginicus (Isoptera: Rhinotermitidae). Bull. Entomol. Res. 1996, 86, 77–81. [Google Scholar] [CrossRef]
- Esenther, G.R.; Beal, R.H. Termite control: Decayed wood bait. Sociobiology 1979, 4, 215–222. [Google Scholar]
- Tarasco, E.; Alvarez, C.S.; Triggiani, O.; Moraga, E.Q. Laboratory studies on the competition for insect haemocoel between Beauveria bassiana and Steinernema ichnusae recovered in the same ecological niche. Biol. Control. Sci. Technol. 2011, 21, 693–704. [Google Scholar] [CrossRef]
- Machado, R.A.R.; Thönen, L.; Arce, C.C.M.; Theepan, V.; Prada, F.; Wüthrich, D.; Robert, C.A.M.; Vogiatzaki, E.; Shi, Y.-M.; Schaeren, O.P.; et al. Engineering bacterial symbionts of nematodes improves their biocontrol potential to counter the western corn rootworm. Nat. Biotechnol. 2020, 38, 600–608. [Google Scholar] [CrossRef]
- Karban, R.; Hougen-Eitzmann, D.; English-Loeb, G. Predator-mediated apparent competition between two herbivores that feed on grapevines. Oecologia 1994, 97, 508–511. [Google Scholar] [CrossRef]
- Faist, A.; Antoninka, A.; Belnap, J.; Bowker, M.A.; Duniway, M.C.; Garcia-Pichel, F.; Nelson, C.; Reed, S.C.; Giraldo-Silva, A.; Ayuso, S.V.; et al. Inoculation and habitat amelioration efforts in biological soil crust recovery vary by desert and soil texture. Restor. Ecol. 2020, 28. [Google Scholar] [CrossRef]
- Temple, S.A. Plant-animal mutualism: Coevolution with dodo leads to near extinction of plant. Science 1977, 197, 885–886. [Google Scholar] [CrossRef]
- Witmer, M.C.; Cheke, A.S. The Dodo and the Tambalacoque tree: An obligate mutualism reconsidered. Oikos 1991, 61, 133. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.Y.; Collinson, M.E.; Rudall, P.J.; Simpson, D.A.; Marone, F.; Stampanoni, M. Virtual taphonomy using synchrotron tomographic microscopy reveals cryptic features and internal structure of modern and fossil plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12013–12018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinseel, E.; Vesela, J.; Scharfen, V.; Urbánková, P.; Van De Vijver, B.; Vyverman, W. Extensive Cryptic Diversity in the Terrestrial Diatom Pinnularia borealis (Bacillariophyceae). Protist 2018, 170, 121–140. [Google Scholar] [CrossRef]
- Hahn, A.S.; Konwar, K.M.; Louca, S.; Hanson, N.W.; Hallam, S.J. The information science of microbial ecology. Curr. Opin. Microbiol. 2016, 31, 209–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roumpeka, D.D.; Wallace, R.J.; Escalettes, F.; Fotheringham, I.; Watson, M. A review of bioinformatics tools for bio-prospecting from metagenomic sequence data. Front. Genet. 2017, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagley, M.; Pilgrim, E.; Knapp, M.; Yoder, C.; Domingo, J.S.; Banerji, A. High-throughput environmental DNA analysis informs a biological assessment of an urban stream. Ecol. Indic. 2019, 104, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Titcomb, G.; Jerde, C.L.; Young, H.S. High-throughput sequencing for understanding the ecology of emerging infectious diseases at the wildlife-human interface. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- van Tienderen, P.H.; de Haan, A.A.; van der Linden, C.G.; Vosman, B. Biodiversity assessment using markers for ecologically important traits. Trends Ecol. Evol. 2002, 17, 577–582. [Google Scholar] [CrossRef]
- Dinsdale, E.A.; Edwards, R.A.; Hall, D.; Angly, F.; Breitbart, M.; Brulc, J.M.; Furlan, M.; Desnues, C.; Haynes, M.; Li, L.; et al. Functional metagenomic profiling of nine biomes. Nat. Cell Biol. 2008, 452, 629–632. [Google Scholar] [CrossRef]
- Peršoh, D. Plant-associated fungal communities in the light of meta’omics. Fungal Divers. 2015, 75, 1–25. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grube, M.; Cernava, T.; Soh, J.; Fuchs, S.; Aschenbrenner, I.A.; Lassek, C.; Wegner, U.; Becher, D.; Riedel, K.; Sensen, C.W.; et al. Exploring functional contexts of symbiotic sustain within lichen-associated bacteria by comparative omics. ISME J. 2015, 9, 412–424. [Google Scholar] [CrossRef] [Green Version]
- Scherlach, K.; Hertweck, C. Mediators of mutualistic microbe–microbe interactions. Nat. Prod. Rep. 2018, 35, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Arango-Argoty, G.; Dai, D.; Pruden, A.; Vikesland, P.J.; Heath, L.S.; Zhang, W.-F. NanoARG: A web service for detecting and contextualizing antimicrobial resistance genes from nanopore-derived metagenomes. Microbiome 2019, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, H.F.; Baycin-Hizal, D.; Lewis, N.E.; Bettenbaugh, M.J. The emerging CHO systems biology era: Harnessing the ‘omics revolution for biotechnology. Curr. Opin. Biotechnol. 2013, 24, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Pirih, N.; Kunej, T. Toward a taxonomy for multi-omics science? Terminology development for whole genome study approaches by omics technology and hierarchy. OMICS A J. Integr. Biol. 2017, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Zorin, A.; Dass, G.; Vu, M.-T.; Xu, P.; Glont, M.; Vizcaíno, J.A.; Jarnuczak, A.F.; Petryszak, R.; Ping, P.; et al. Quantifying the impact of public omics data. Nat. Commun. 2019, 10, 3512. [Google Scholar] [CrossRef] [Green Version]
- Ratnaparkhe, A.; Tiwari, A. Bio-prospecting approach for biocatalysts from extremophiles. Int. J. Biomed. Res. 2011, 2, 1–17. [Google Scholar] [CrossRef] [Green Version]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Verberk, W.C.E.P.; Van Noordwijk, C.G.E.; Hildrew, A.G. Delivering on a promise: Integrating species traits to transform descriptive community ecology into a predictive science. Freshw. Sci. 2013, 32, 531–547. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.; Twieg, B.D.; Ward, V.; Barker, J.; Durall, D.M.; Simard, S.W. Functional complementarity of Douglas-fir ectomycorrhizas for extracellular enzyme activity after wildfire or clearcut logging. Funct. Ecol. 2010, 24, 1139–1151. [Google Scholar] [CrossRef]
- Blüthgen, N.; Klein, A.-M. Functional complementarity and specialisation: The role of biodiversity in plant-pollinator interactions. Basic Appl. Ecol. 2011, 12, 282–291. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banerji, A. Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications. Ecologies 2021, 2, 43-70. https://doi.org/10.3390/ecologies2010004
Banerji A. Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications. Ecologies. 2021; 2(1):43-70. https://doi.org/10.3390/ecologies2010004
Chicago/Turabian StyleBanerji, Aabir. 2021. "Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications" Ecologies 2, no. 1: 43-70. https://doi.org/10.3390/ecologies2010004
APA StyleBanerji, A. (2021). Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications. Ecologies, 2(1), 43-70. https://doi.org/10.3390/ecologies2010004