Plant Diversity in Sardinian Mountain Rangelands: Analysis of Its Relationships with Grazing, Land Management, and Pastoral Value

 , ,
, ,
Abstract
:1. Introduction
2. Experimental Section
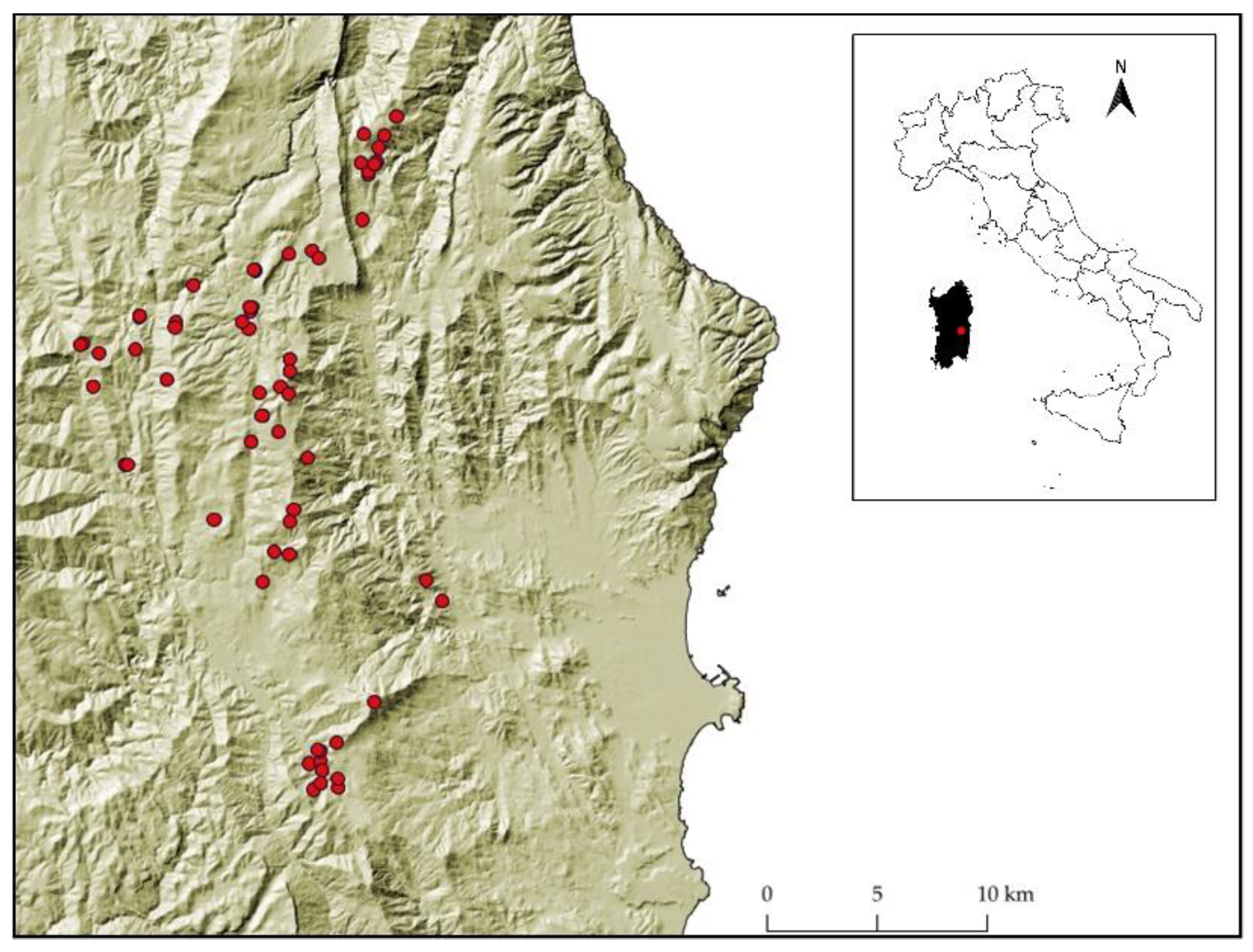
2.1. Study Area
2.2. Vegetation Surveys
2.3. Estimation of Response Variables
Plant Diversity and Pastoral Value
2.4. Estimation of Explanatory Variables
Abiotic and Biotic Predictors
2.5. Statistical Analysis
3. Results
3.1. Floristic Richness, Native and Endemic Species
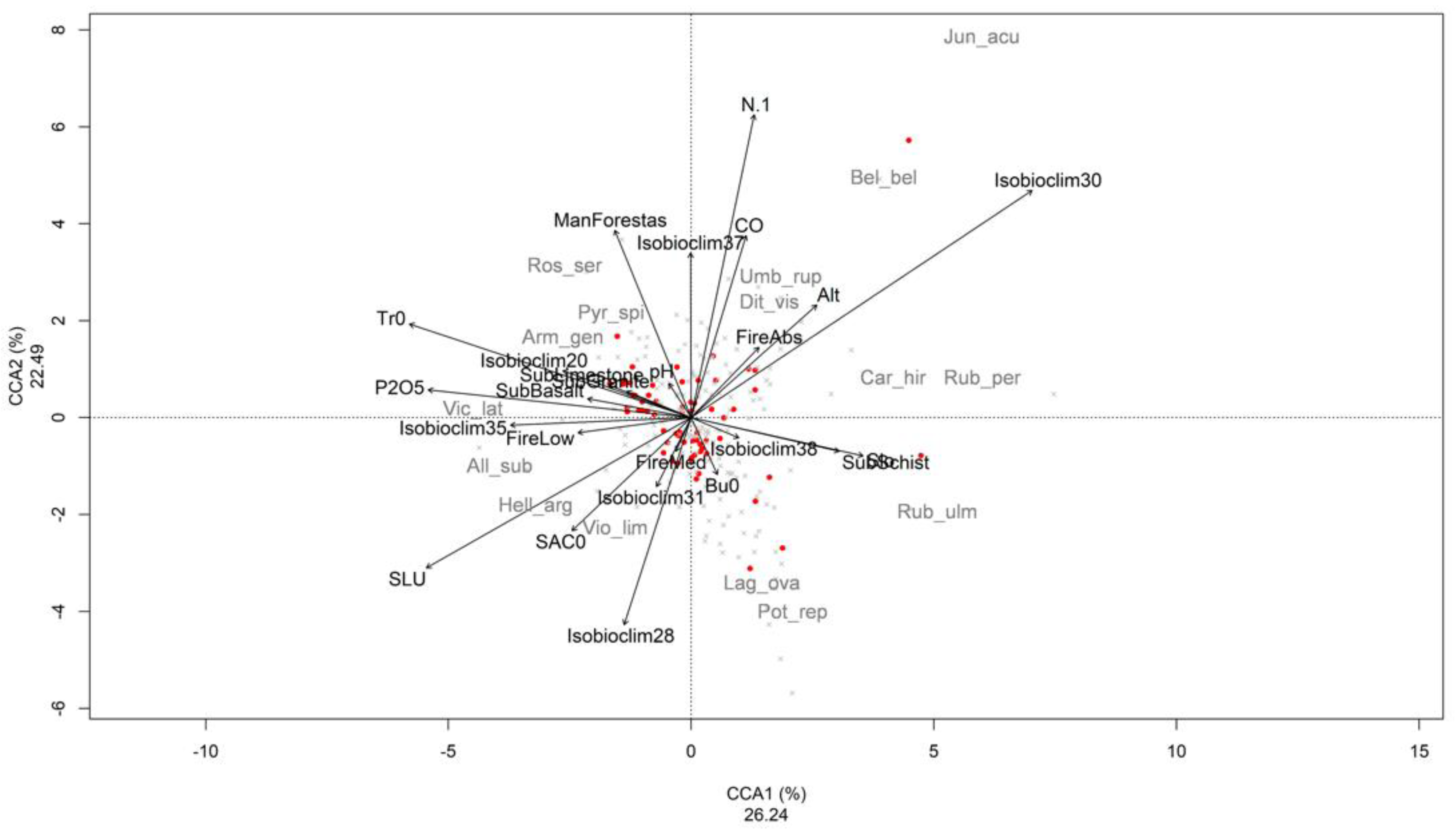
3.2. Ordination Analysis
3.3. Pastoral Value
3.4. Management Type
3.5. Effect of Grazing on Plant Traits
4. Discussion
4.1. Plant Diversity, Endemic Species, and Pastoral Value
4.2. Effect of Grazing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ragab, R.; Prudhomme, C. SW—Soil and water. Biosyst. Eng. 2002, 81, 3–34. [Google Scholar] [CrossRef]
- Follett, R.F.; Reed, D.A. Soil carbon sequestration in grazing lands: Societal benefits and policy implications. Rangel. Ecol. Manag. 2010, 63, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Jouven, M.; Lapeyronie, P.; Moulin, C.H.; Bocquier, F. Rangeland utilization in Mediterranean farming systems. Animal 2010, 4, 1746–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-H.; Grant, W.E.; Teague, R. Modeling rangelands as spatially-explicit complex adaptive systems. J. Environ. Manag. 2020, 269, 110762. [Google Scholar] [CrossRef]
- Jacobo, E.J.; Rodriguez, A.M.; Bartoloni, N.; Deregibus, V.A. Rotational grazing effects on rangeland vegetation at a farm scale. Rangelands 2006, 59, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Briske, D.; Derner, J.; Brown, J.; Fuhlendorf, S.; Teague, W.; Havstad, K.; Gillen, R.; Ash, A.; Willms, W. Rotational grazing on rangelands: Reconciliation of perception and experimental evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Teague, R.; Provenza, F.; Kreuter, U.; Steffens, T.; Barnes, M. Multi-paddock grazing on rangelands: Why the perceptual dichotomy between research results and rancher experience? J. Environ. Manag. 2013, 128, 699–717. [Google Scholar] [CrossRef]
- Sabatini, S.; Argenti, G.; Staglianò, N.; Bianchetto, E. Il Monitoraggio delle Risorse Prative e Pascolive per la Definizione di Idonee Linee di Gestione Pastorale Sostenibile; Comunicazioni di Ricerca 2001/2; Istituto Sperimentale per l’Assestamento Forestale e l’Alpicoltura (ISAFA): Trento, Italy, 2001; pp. 93–99. [Google Scholar]
- Papanastasis, V.P.; Bautista, S.; Chouvardas, D.; Mantzanas, K.; Papadimitriou, M.; Mayor, A.G.; Koukioumi, P.; Papaioannou, A.; Vallejo, R.V. Comparative assessment of goods and services provided by grazing regulation and reforestation in degraded Mediterranean rangelands. Land Degrad. Dev. 2017, 28, 1178–1187. [Google Scholar] [CrossRef] [Green Version]
- Bagella, S.; Caria, M.C.; Seddaiu, G.; Leites, L.; Roggero, P.P. Patchy landscapes support more plant diversity and ecosystem services than wood grasslands in Mediterranean silvopastoral agroforestry systems. Agric. Syst. 2020, 185, 102945. [Google Scholar] [CrossRef]
- Farris, E.; Secchi, Z.; Rosati, L.; Filigheddu, R. Are all pastures eligible for conservation? A phytosociological survey of the Sardinian–Corsican Province as a basic tool for the Habitats Directive. Plant Biosyst. 2013, 147, 931–946. [Google Scholar] [CrossRef]
- Camarda, I. Indicators of Degradation in Agro-ecosystems. In Indicators for Assessing Desertification in the Mediterranean; Os-servatorio Nazionale sulla Desertificazione: Porto Torres, Italy, 1998; pp. 159–163. [Google Scholar]
- Merdas, S.; Menad, A.; Mostephaoui, T.; Sakaa, B. Plant community structure and diversity under grazing gradient in arid Mediterranean steppe of Algeria. J. Mater. Environ. Sci. 2017, 8, 4329–4338. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Arrigoni, P.V. Flora Dell’Isola di Sardegna; Carlo Delfino Editore: Sassari, Italy, 2006; Volumes 1–6. [Google Scholar]
- Camarda, I.; Carta, L.; Brunu, A.; Angelini, P.; Laureti, L. Carta degli Habitat della Regione Sardegna per il Sistema Informativo di Carta della Natura alla Scala 1:50.000; ISPRA-Regione Sardegna Università degli Studi di Sassari: Sassari, Italy, 2010. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R package version 2.5-7. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 December 2020).
- Vacca, G.; Camarda, I.; Brundu, G.; Sanna, F.; Caredda, S.; Re, G. Comparison of quadrate and transect survey methods to evaluate Pastoral Value (PV) in SE Sardinian rangelands. Options Méditerranéennes Série A Séminaires Méditerranéens 2016, 114, 135–138. [Google Scholar]
- Tomaselli, R.I. Introduzione allo Studio della Fitosociologia; Industria Poligrafica Lombarda: Milano, Italy, 1956; 319p. [Google Scholar]
- Roggero, P.P.; Bagella, S.; Farina, R. Un archivio dati di Indici Specifici per la valutazione integrata del Valore Pastorale. Rivista di Agronomia 2002, 36, 149–156. [Google Scholar]
- Cavallero, A.; Aceto, P.; Gorlier, A.; Lombardi, G.; Lonati, M.; Martinasso, B.; Tagliatori, C. I Tipi Pastorali delle Alpi Piemontesi; Alberto Perdisa Editore: Bologna, Italy, 2007. [Google Scholar]
- Bagella, S. Valore Pastorale delle associazioni vegetali: Un esempio di applicazione nell’Appennino Umbro-Marchigiano, Italia. Fitosociologia 2001, 38, 153–165. [Google Scholar]
- Aru, A.; Baldaccini, P.; Delogu, G.; Dessena, M.A.; Madrau, S.; Melis, S.T.; Vacca, S.; Vacca, A. Carta dei Suoli della Sardegna; Regione Autonoma della Sardegna: Cagliari, Italy, 1989; Available online: http://www.sardegnaportalesuolo.it/ (accessed on 15 December 2020).
- Canu, S.; Rosati, L.; Fiori, M.; Motroni, A.; Filigheddu, R.; Farris, E. Bioclimate map of Sardinia, Italy. J. Maps 2015, 11, 711–718. [Google Scholar] [CrossRef] [Green Version]
- Ministero delle Politiche Agricole e Forestali. Decreto Ministeriale 13 Settembre 1999. Approvazione dei Metodi ufficiali di analisi chimica del suolo. (GU Serie Generale n. 248 del 21-10-1999-Suppl. Ordinario n. 185). Rome (Italy). Available online: https://www.gazzettaufficiale.it/eli/id/1999/10/21/099A8497/sg (accessed on 15 December 2020).
- Pebesma, E.J. Multivariable geostatistics in S: The gstat package. Comput. Geosci. 2004, 30, 683–691. [Google Scholar] [CrossRef]
- Olmedo, O.E. Kriging: Ordinary Kriging. R package Version 1.1. 2014. Available online: https://CRAN.R-project.org/package=kriging (accessed on 15 December 2020).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Usinglme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- International-Harvard Statistical Consulting Company. My Stepwise: Stepwise Variable Selection Procedures for Regression Analysis; R package Version 0.1.0; International-Harvard Statistical Consulting Company: Taipei, Taiwan, 2017. [Google Scholar]
- Abbasi-Kesbi, M.; Tataian, M.R.; Tamartash, R.; Fattahi, B. Relationships between soil properties and plant diversity indices (Case Study: Lashgardar Protected Rangeland, Malyer, Iran). J. Rangel. Sci. 2016, 7, 79–89. [Google Scholar]
- Hobohm, C.; Janišová, M.; Jansen, J.; Bruchmann, I.; Deppe, U. Biogeography of endemic vascular plants–overview. In Endemism in Vascular Plants; Springer: Dordrecht, The Netherlands, 2014; pp. 85–163. [Google Scholar]
- Thompson, J.D.; Lavergne, S.; Affre, L.; Gaudeul, M.; Debussche, M. Ecological differentiation of Mediterranean endemic plants. Taxon 2005, 54, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Brundu, G.; Peruzzi, L.; Domina, G.; Bartolucci, F.; Galasso, G.; Peccenini, S.; Raimondo, F.M.; Albano, A.; Alessandrini, A.; Banfi, E.; et al. At the intersection of cultural and natural heritage: Distribution and conservation of the type localities of Italian endemic vascular plants. Biol. Conserv. 2017, 214, 109–118. [Google Scholar] [CrossRef]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Fois, M.; Fenu, G.; Cañadas, E.M.; Bacchetta, G. Disentangling the influence of environmental and anthropogenic factors on the distribution of endemic vascular plants in Sardinia. PLoS ONE 2017, 12, e0182539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trigas, P.; Panitsa, M.; Tsiftsis, S. Elevational gradient of vascular plant species richness and endemism in Crete–The effect of post-isolation mountain uplift on a continental island system. PLoS ONE 2013, 8, e59425. [Google Scholar] [CrossRef]
- Aronson, J.; Shmida, A. Plant species diversity along a Mediterranean-desert gradient and its correlation with interannual rainfall fluctuations. J. Arid. Environ. 1992, 23, 235–247. [Google Scholar] [CrossRef]
- Drissen, T.; Faust, C.; Stadtmann, R.; Treitler, J.T.; Zerbe, S.; Mantilla-Contreras, J. Plant composition and diversity in a semi-natural Mediterranean island landscape: The importance of environmental factors. Plant Biosyst. 2019, 153, 756–766. [Google Scholar] [CrossRef]
- Chytrý, M.; Danihelka, J.; Axmanová, I.; Božková, J.; Hettenbergerova, E.; Li, C.; Rozbrojova, Z.; Sekulová, L.; Tichý, L.; Vymazalova, M. Floristic diversity of an Eastern Mediterranean dwarf shrubland: The importance of soil pH. J. Veg. Sci. 2010, 21, 1125–1137. [Google Scholar] [CrossRef]
- Médail, F. The specific vulnerability of plant biodiversity and vegetation on Mediterranean islands in the face of global change. Reg. Environ. Chang. 2017, 17, 1775–1790. [Google Scholar] [CrossRef] [Green Version]
- Noy-Meir, I.; Oron, T. Effects of grazing on geophytes in Mediterranean vegetation. J. Veg. Sci. 2001, 12, 749–760. [Google Scholar] [CrossRef]
- Camarda, I.; Brundu, G.; Carta, L.; Vacca, G.; Brunu, A. Plants and grazing: An evaluation of the effects on Sardinian endemic plants conservation. Flora Mediterranea 2015, 25, 263–275. [Google Scholar]
- Naveh, Z.; Whittaker, R.H. Structural and floristic diversity of shrublands and woodlands in Northern Israel and other Mediterranean areas. Vegetatio 1980, 41, 171–190. [Google Scholar] [CrossRef]
- Ahlborn, J.; Von Wehrden, H.; Lang, B.; Römermann, C.; Oyunbileg, M.; Oyuntsetseg, B.; Wesche, K. Climate–grazing interactions in Mongolian rangelands: Effects of grazing change along a large-scale environmental gradient. J. Arid. Environ. 2020, 173, 104043. [Google Scholar] [CrossRef]
- Pisanu, S.; Farris, E.; Filigheddu, R.; García, M.B. Demographic effects of large, introduced herbivores on a long-lived endemic Plant. Plant Ecol. 2012, 213, 1543–1553. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Type of Variable | Variable | Unit | Mean | Sd | Min | Max |
|---|---|---|---|---|---|---|
| Response variables | ||||||
| Pastoral value (PV) | No. | 29.43 | 6.23 | 18.05 | 46.27 | |
| Species richness | No. | 42.35 | 7.91 | 27.00 | 67.00 | |
| Evenness (J) | No. | 0.81 | 0.04 | 0.71 | 0.92 | |
| Endemic species richness | No. | 2.39 | 1.05 | 1.00 | 4.00 | |
| Endemic species—plot cover | % | 5.88 | 3.79 | 1.00 | 39.5 | |
| Life forms—plot cover | % | Seven types found in the study area | ||||
| Life span—plot cover | % | Three types considered | ||||
| Pollination type—plot cover | % | Two types considered | ||||
| Presence of thorns—plot cover | % | 7.16 | 8.05 | 1.00 | 41.00 | |
| Explanatory variables | ||||||
| Abiotic | pH | No. | 5.91 | 0.54 | 4.80 | 7.75 |
| OC | % | 4.90 | 1.46 | 1.32 | 7.80 | |
| P2O5 | ppm | 8.38 | 6.15 | 1.86 | 35.25 | |
| N | g·kg−1 | 4.52 | 2.06 | 0.41 | 10.54 | |
| Altitude | m a.s.l. | 1011 | 195 | 280 | 1362 | |
| Slope | ° | 11.63 | 8.25 | 0 | 41.62 | |
| Geological substrate | Five types found in the study area | |||||
| Fire | Classified into four categories | |||||
| Iso-bioclimatic type | Eight types found in the study area | |||||
| Biotic | Spatialized livestock unit pressure (SLU) | LU·km−2 | 35.32 | 23.63 | 0.11 | 115.90 |
| Trees | – | – | – | 0 | 1 | |
| Shrubs | – | – | – | 0 | 1 | |
| Management | Land management | Common land vs. land managed by FoReSTAS | ||||
| Special Areas of Conservation (SAC) | – | – | – | 0 | 1 | |
| Predictors | r2 | Pr (>r) |
|---|---|---|
| Trees | 0.07 | 0.01 * |
| P2O5 (ppm) | 0.14 | 0.01 * |
| N (g·kg−1) | 0.02 | 0.02 * |
| SLU (kg·km−2) | 0.12 | 0.02 * |
| Iso-bioclimatic type | 0.18 | 0.09 |
| pH | 0.06 | 0.14 |
| Altitude (m a.s.l.) | 0.06 | 0.15 |
| Slope (°) | 0.04 | 0.26 |
| Land management | 0.02 | 0.26 |
| Geological substrate | 0.10 | 0.28 |
| SAC | 0.02 | 0.31 |
| OC (%) | 0.02 | 0.60 |
| Fire | 0.02 | 0.88 |
| Shrubs | 0.00 | 0.96 |
| Response Variables | Predictors | Estimate | Std. Error | z Value | p-Value |
|---|---|---|---|---|---|
| Pastoral value | N (g·kg−1) | 1.03 | 0.38 | 2.69 | 0.01 ** |
| SLU (kg·km−2) | 0.08 | 0.03 | 2.27 | 0.03 * | |
| Iso-bioclimatic type 20 | 0.55 | 0.25 | 2.23 | 0.03 * | |
| Species richness | SLU (kg·km−2) | 0.01 | 0.00 | 13.81 | <2 × 10−16 *** |
| Management: FoReSTAS | −0.19 | 0.02 | −7.86 | 4.00 × 10−15 *** | |
| N (g·kg−1) | 0.06 | 0.01 | 7.11 | 1.17 × 10−12 *** | |
| Altitude (m a.s.l.) | 0.00 | 0.00 | 5.94 | 2.93 × 10−9 *** | |
| OC (%) | 13.14 | 3.39 | 3.88 | 1.00 × 10−4 *** | |
| P2O5 (ppm) | −0.01 | 0.02 | −3.30 | 1.00 × 10−3 *** | |
| Iso-bioclimatic type 31 | −0.46 | 0.15 | −3.00 | 3.00 × 10−3 ** | |
| Iso-bioclimatic type 20 | 0.30 | 0.13 | 2.28 | 0.02 * | |
| Endemic species | Altitude (m a.s.l.) | 0.00 | 0.00 | 2.91 | 3.00 × 10−3 ** |
| SLU (kg·km−2) | −0.01 | 0.01 | −2.35 | 0.02 * |
| Response Variables | Estimate | Std. Error | z Value | p-Value |
|---|---|---|---|---|
| Chamaephytes | 0.02 | 0.10 | 0.17 | 0.87 |
| Geophytes | 0.01 | 0.04 | 0.18 | 0.86 |
| Hemicryptophytes | 0.26 | 0.13 | 2.01 | 0.05 * |
| Helophytes | −0.02 | 0.01 | −1.59 | 0.12 |
| Nanophanerophytes | −0.00 | 0.05 | −0.02 | 0.98 |
| Phanerophytes | −0.02 | 0.07 | −0.24 | 0.81 |
| Therophytes | 0.17 | 0.20 | 0.86 | 0.39 |
| Annual | 0.17 | 0.20 | 0.86 | 0.39 |
| Biennial | 0.08 | 0.03 | 2.68 | 0.01 ** |
| Perennial | 0.18 | 0.19 | 0.95 | 0.35 |
| Thorns | 0.06 | 0.09 | 0.64 | 0.53 |
| Anemophily | 0.23 | 0.16 | 1.49 | 0.14 |
| Entomophily | 0.19 | 0.21 | 0.91 | 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano, V.; Vacca, G.; Camarda, I.; Brundu, G.; Sanna, F.; Caredda, S.; Re, G.A. Plant Diversity in Sardinian Mountain Rangelands: Analysis of Its Relationships with Grazing, Land Management, and Pastoral Value. Ecologies 2021, 2, 164-174. https://doi.org/10.3390/ecologies2010009
Lozano V, Vacca G, Camarda I, Brundu G, Sanna F, Caredda S, Re GA. Plant Diversity in Sardinian Mountain Rangelands: Analysis of Its Relationships with Grazing, Land Management, and Pastoral Value. Ecologies. 2021; 2(1):164-174. https://doi.org/10.3390/ecologies2010009
Chicago/Turabian StyleLozano, Vanessa, Gabriella Vacca, Ignazio Camarda, Giuseppe Brundu, Federico Sanna, Salvatore Caredda, and Giovanni Antonio Re. 2021. "Plant Diversity in Sardinian Mountain Rangelands: Analysis of Its Relationships with Grazing, Land Management, and Pastoral Value" Ecologies 2, no. 1: 164-174. https://doi.org/10.3390/ecologies2010009
APA StyleLozano, V., Vacca, G., Camarda, I., Brundu, G., Sanna, F., Caredda, S., & Re, G. A. (2021). Plant Diversity in Sardinian Mountain Rangelands: Analysis of Its Relationships with Grazing, Land Management, and Pastoral Value. Ecologies, 2(1), 164-174. https://doi.org/10.3390/ecologies2010009