
Fucoxanthinol Promotes Apoptosis in MCF-7 and MDA-MB-231 Cells by Attenuating Laminins–Integrins Axis


 and
and
Abstract
:Simple Summary
Abstract
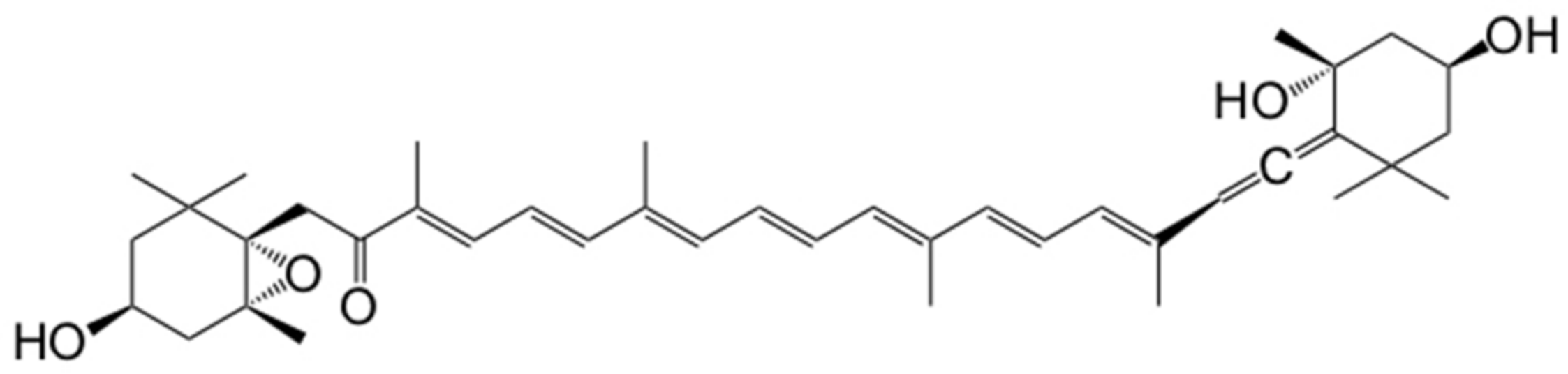
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Viability Assay
2.3. Analysis of Apoptosis-Associated Nuclear Alteration
2.4. Analyses of Apoptotic-Like Cell Body and Cell Cycle Phases
2.5. Extraction and Purification of Total RNA
2.6. Microarray Analysis
2.7. Analysis of Protein Expression and Activation
2.8. Gene Knockdown Experiments
2.9. Statistical Analysis
3. Results
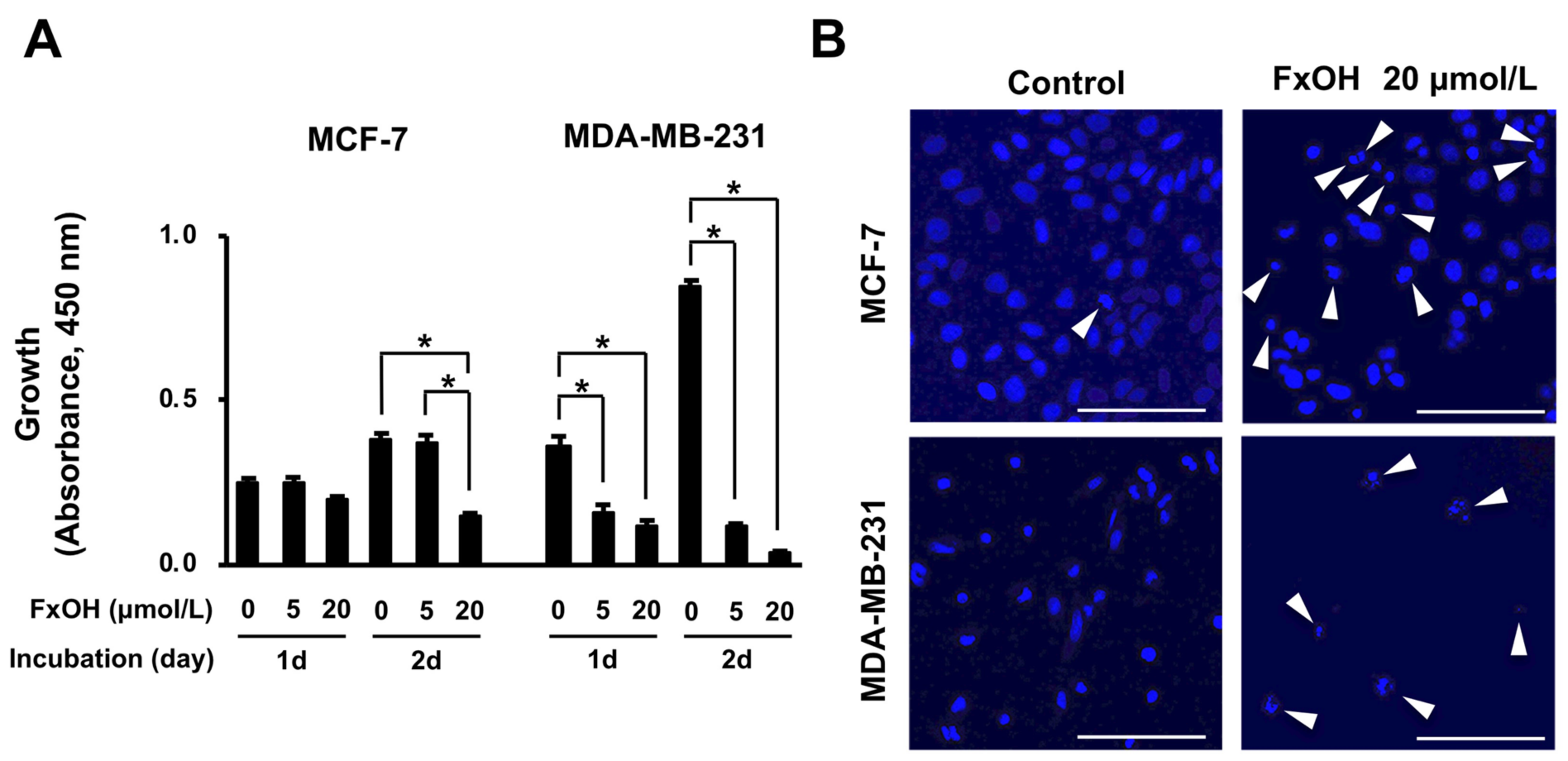
3.1. Effects of FxOH on Cell Growth, Apoptosis, and Cell Cycle in MCF-7 and MDA-MB-231 Cells
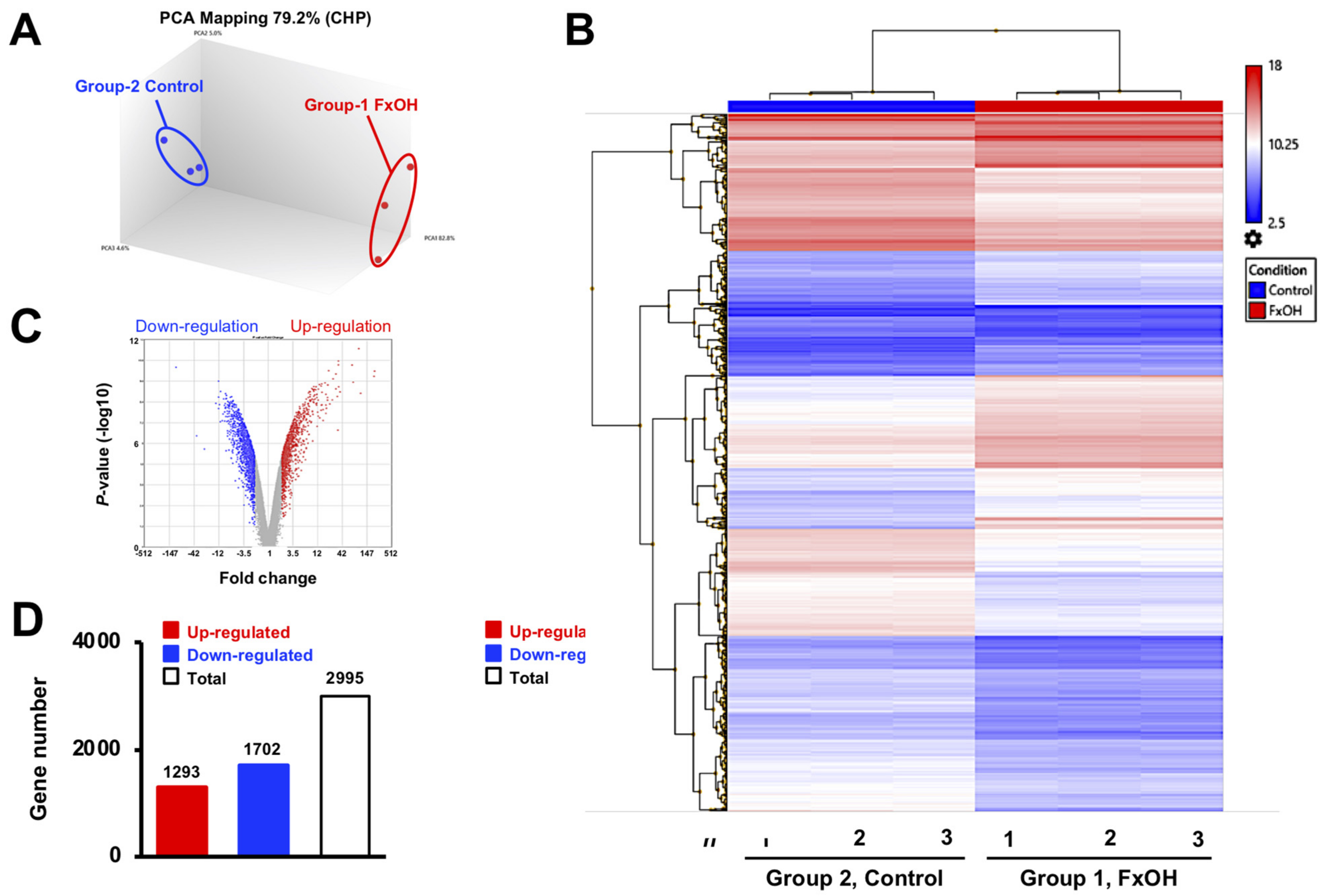
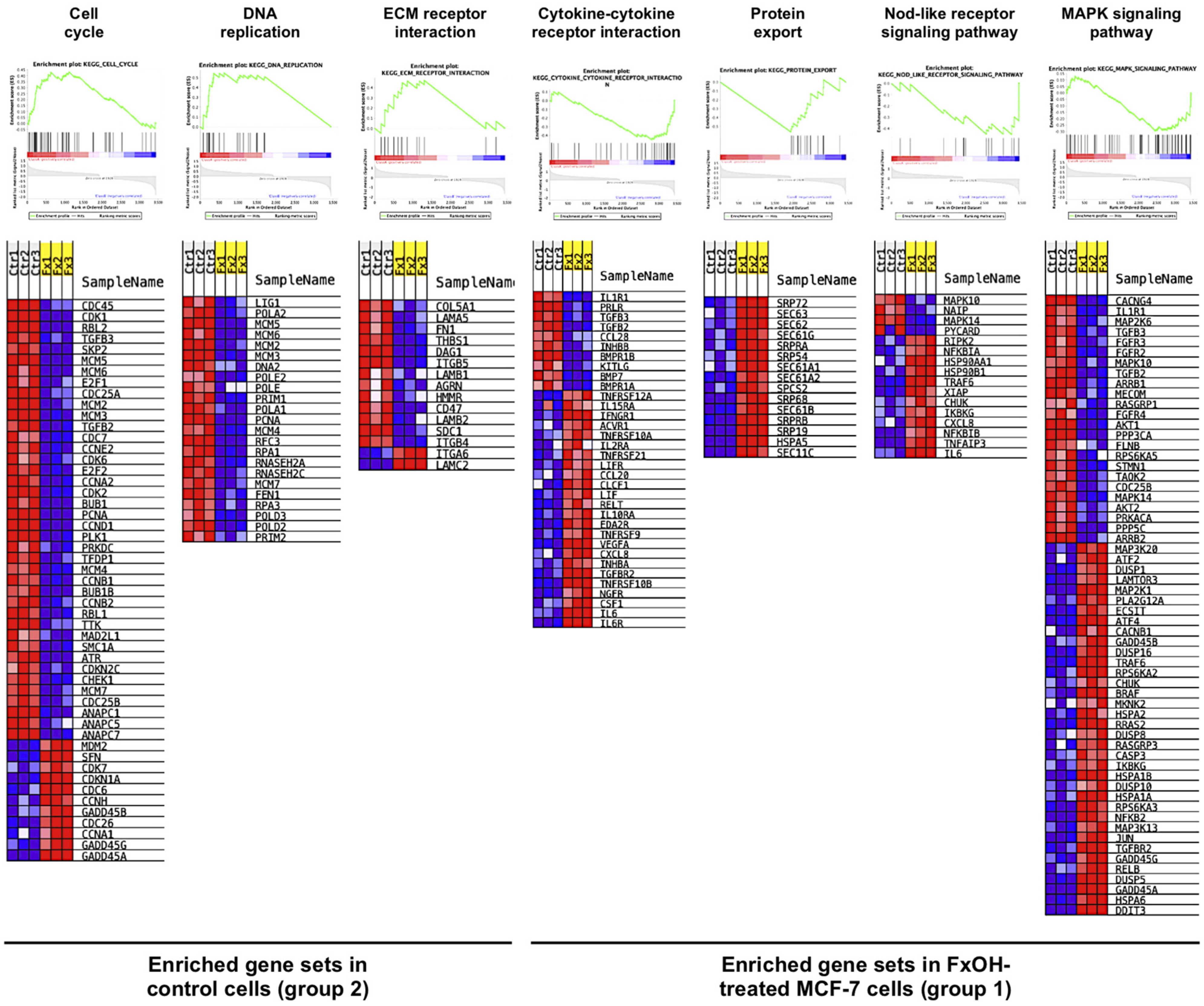
3.2. Effects of FxOH on the Transcriptome Profiles of MCF-7 Cells
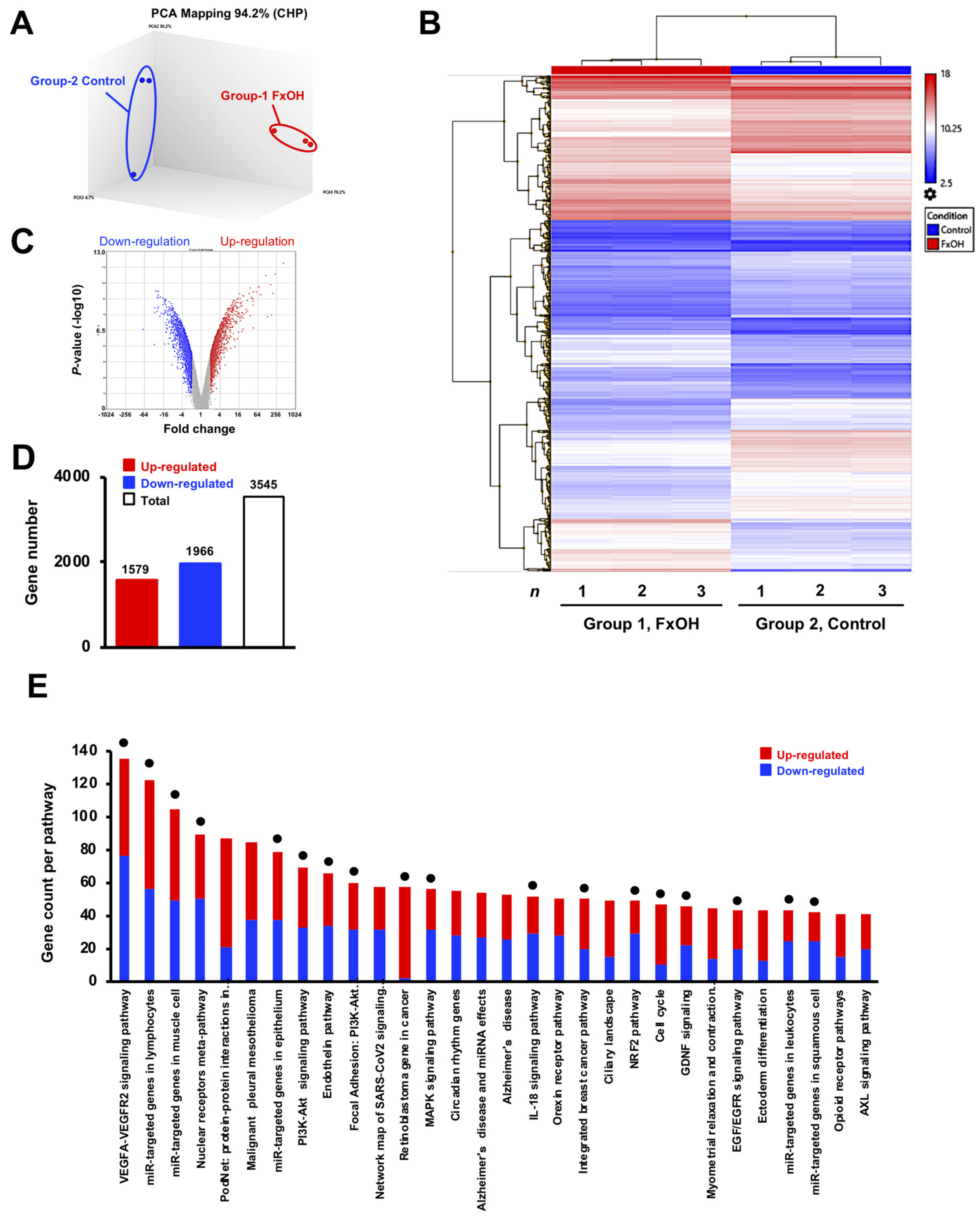
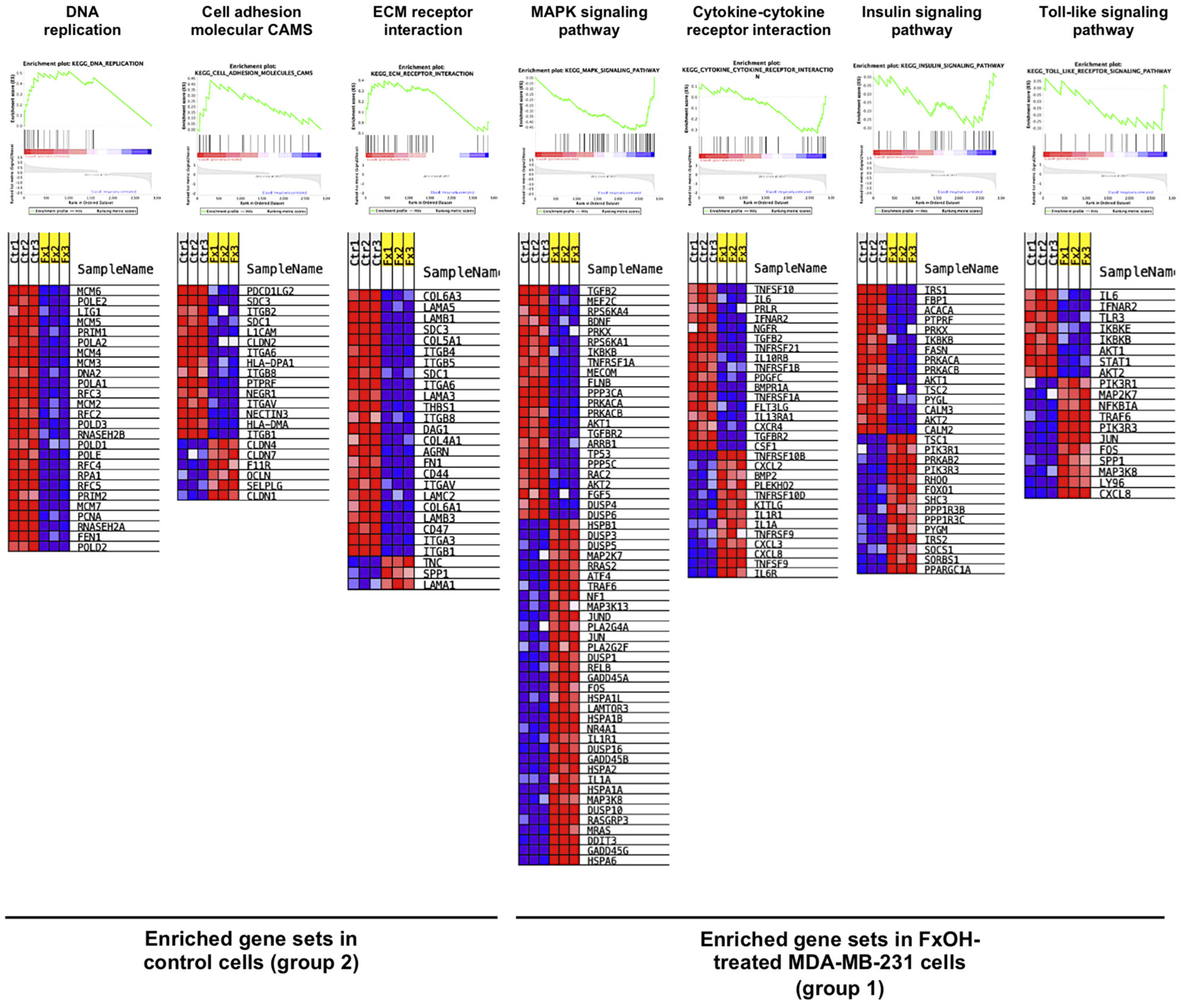
3.3. Effects of FxOH on the Transcriptome Profiles of MDA-MB-231 Cells
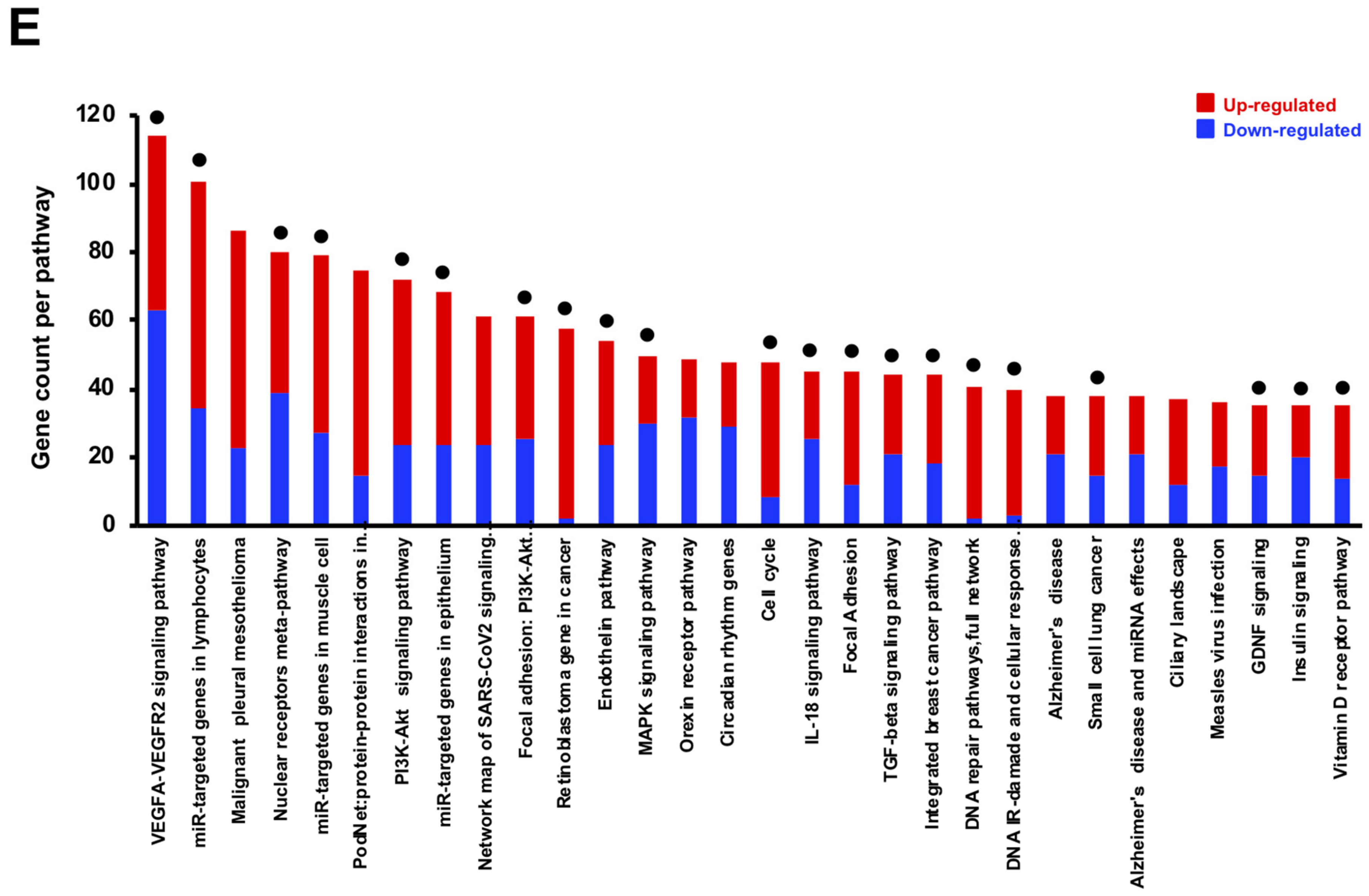
3.4. Bioinformatics Analyses of Transcriptome Profiles of FxOH-Treated MCF-7 and MDA-MB-231 Cells
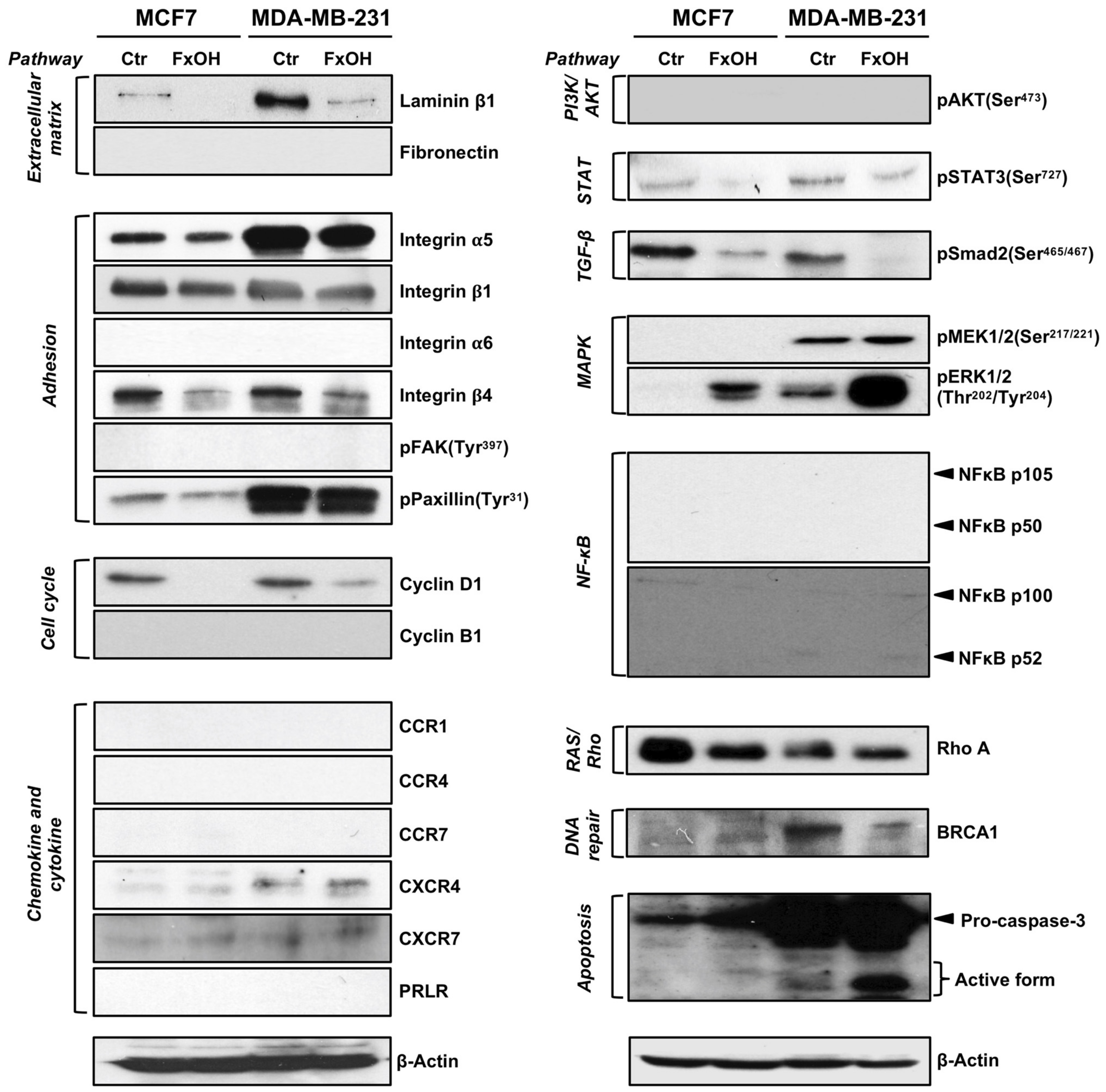
3.5. Effects of FxOH on Protein Expression and Activation in MCF-7 and MDA-MB-231 Cells
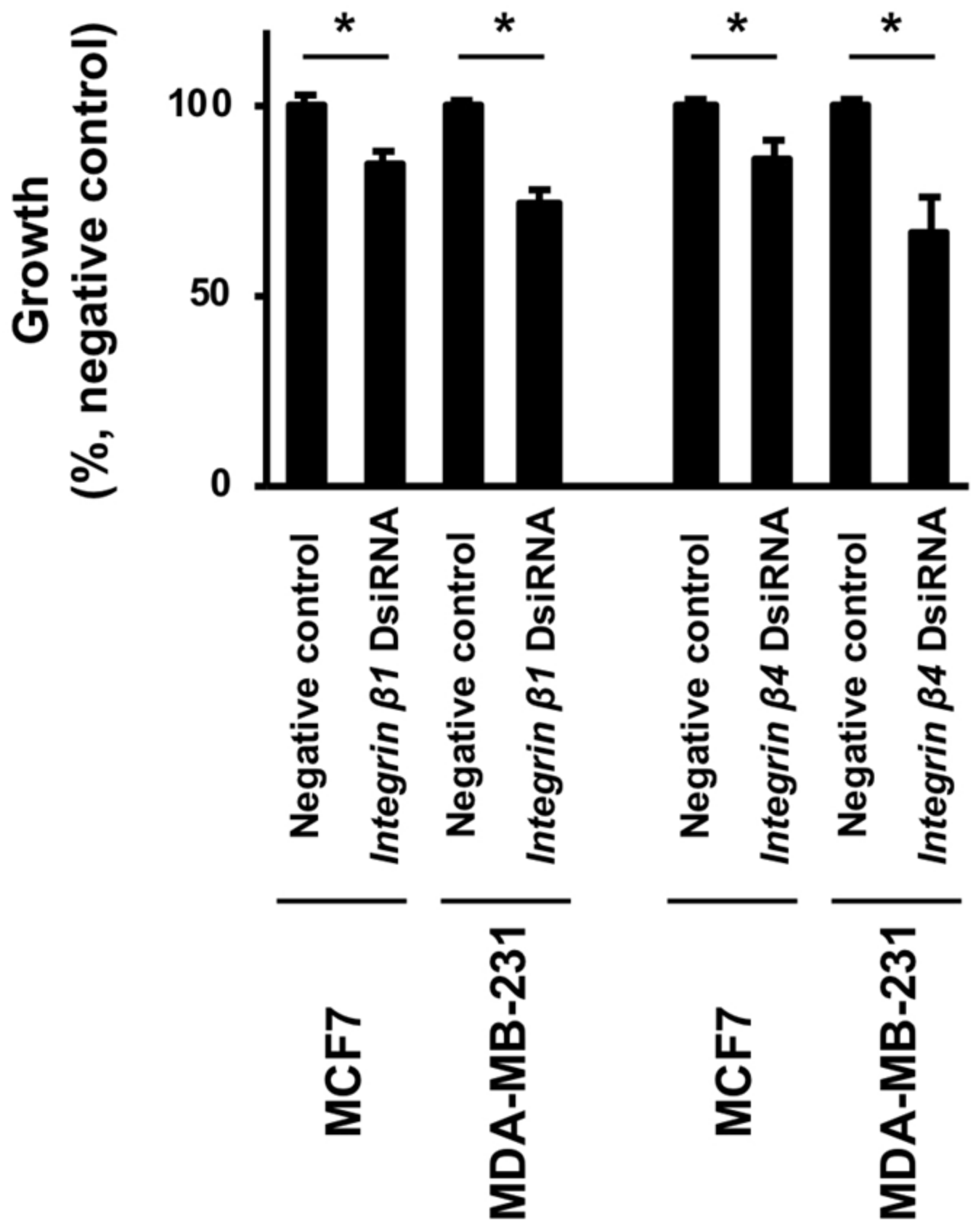
3.6. Effects of FxOH on the Cell Growth of Integrin β1 or β4 Knockdown MCF-7 and MDA-MB-231 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Wehner, M.R.; Matrisian, L.M.; Nead, K.T. Estimated projection of US cancer incidence and death to 2040. JAMA Netw. Open 2021, 4, e214708. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Breast Cancer; American Cancer Society: Atlanta, GA, USA, 2021; Available online: https://www.cancer.org/cancer/breast-cancer.html (accessed on 3 June 2022).
- Miller, N.J.; Sampson, J.; Candeias, L.P.; Bramley, P.M.; Rice-Evans, C.A. Antioxidant activities of carotenes and xanthophylls. FEBS Lett. 1996, 384, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- World Cancer Research Fund/American Institute for Cancer Research. Diet, Nutrition, Physical Activity and Cancer: A Global Perspective, a Summary of the Third Expert Report; World Cancer Research Fund (WCRF) International; WCRF International: London, UK, 2018; p. 38. [Google Scholar]
- Mikami, K.; Hosokawa, M. Biosynthesis pathway and health benefits of fucoxanthin, an algae-specific xanthophyll in brown seaweeds. Int. J. Mol. Sci. 2013, 14, 13763–13781. [Google Scholar] [CrossRef] [Green Version]
- Terasaki, M.; Kubota, A.; Kojima, H.; Maeda, H.; Miyashita, K.; Kawagoe, C.; Mutoh, M.; Tanaka, T. Fucoxanthin and colorectal cancer prevention. Cancers 2021, 13, 2379. [Google Scholar] [CrossRef]
- Tavares, R.S.N.; Maria-Engler, S.S.; Colepicolo, P.; Debonsi, H.M.; Schäfer-Korting, M.; Marx, U.; Gaspar, L.R.; Zoschke, C. Skin irritation testing beyond tissue viability: Fucoxanthin effects on inflammation, homeostasis, and metabolism. Pharmaceutics 2020, 12, 136. [Google Scholar] [CrossRef] [Green Version]
- Beppu, F.; Niwano, Y.; Tsukui, T.; Hosokawa, M.; Miyashita, K. Single and repeated oral dose toxicity study of fucoxanthin (FX), a marine carotenoid, in mice. J. Toxicol. Sci. 2009, 34, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Asai, A.; Yonekura, L.; Nagao, A. Low bioavailability of dietary epoxyxanthophylls in humans. Br. J. Nutr. 2008, 100, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T.; Ozaki, Y.; Mizuno, M.; Yoshida, M.; Nishitani, Y.; Azuma, T.; Komoto, A.; Maoka, T.; Tanino, Y.; Kanazawa, K. Pharmacokinetics of fucoxanthinol in human plasma after the oral administration of kombu extract. Br. J. Nutr. 2012, 107, 1566–1569. [Google Scholar] [CrossRef]
- Yonekura, L.; Kobayashi, M.; Terasaki, M.; Nagao, A. Keto-carotenoids are the major metabolites of dietary lutein and fucoxanthin in mouse tissues. J. Nutr. 2010, 140, 1824–1831. [Google Scholar] [CrossRef]
- Shiratori, K.; Ohgami, K.; Ilieva, I.; Jin, X.H.; Koyama, Y.; Miyashita, K.; Yoshida, K.; Kase, S.; Ohno, S. Effects of fucoxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Exp. Eye Res. 2005, 81, 422–428. [Google Scholar] [CrossRef]
- Hitoe, S.; Shimoda, H. Seaweed fucoxanthin supplementation improves obesity parameters in mild obese Japanese subjects. Funct. Foods Health Dis. 2017, 7, 246–262. [Google Scholar] [CrossRef]
- Mikami, N.; Hosokawa, M.; Miyashita, K.; Sohma, H.; Ito, Y.M.; Kokai, Y. Reduction of HbA1c levels by fucoxanthin-enriched akamoku oil possibly involves the thrifty allele of uncoupling protein 1 (UCP1): A randomised controlled trial in normal-weight and obese Japanese adults. J. Nutr. Sci. 2017, 6, e5. [Google Scholar] [CrossRef] [Green Version]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical scavenging and singlet oxygen quenching activity of marine carotenoid fucoxanthin and its metabolites. J. Agric. Food Chem. 2007, 55, 8516–8522. [Google Scholar] [CrossRef]
- Sugawara, T.; Matsubara, K.; Akagi, R.; Mori, M.; Hirata, T. Antiangiogenic activity of brown algae fucoxanthin and its deacetylated product, fucoxanthinol. J. Agric. Food Chem. 2006, 54, 9805–9810. [Google Scholar] [CrossRef]
- Nishino, H.; Murakoshi, M.; Tokuda, H.; Satomi, Y. Cancer prevention by carotenoids. Arch. Biochem. Biophys. 2009, 483, 165–168. [Google Scholar] [CrossRef]
- Terasaki, M.; Maeda, H.; Miyashita, K.; Tanaka, T.; Miyamoto, S.; Mutoh, M. A marine bio-functional lipid, fucoxanthinol, attenuates human colorectal cancer stem-like cell tumorigenicity and sphere formation. J. Clin. Biochem. Nutr. 2017, 61, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Rwigemera, A.; Mamelona, J.; Martin, L.J. Inhibitory effects of fucoxanthinol on the viability of human breast cancer cell lines MCF-7 and MDA-MB-231 are correlated with modulation of the NF-kappaB pathway. Cell Biol. Toxicol. 2014, 30, 157–167. [Google Scholar] [CrossRef]
- Rwigemera, A.; Mamelona, J.; Martin, L.J. Comparative effects between fucoxanthinol and its precursor fucoxanthin on viability and apoptosis of breast cancer cell lines MCF-7 and MDA-MB-231. Anticancer Res. 2015, 35, 207–219. [Google Scholar]
- Wang, J.; Ma, Y.; Yang, J.; Jin, L.; Gao, Z.; Xue, L.; Hou, L.; Sui, L.; Liu, J.; Zou, X. Fucoxanthin inhibits tumour-related lymphangiogenesis and growth of breast cancer. J. Cell. Mol. Med. 2019, 23, 2219–2229. [Google Scholar] [CrossRef]
- Malhão, F.; Macedo, A.C.; Costa, C.; Rocha, E.; Ramos, A.A. Fucoxanthin holds potential to become a drug adjuvant in breast cancer treatment: Evidence from 2D and 3D cell cultures. Molecules 2021, 26, 4288. [Google Scholar] [CrossRef]
- Eid, S.Y.; Althubiti, M.A.; Abdallah, M.E.; Wink, M.; El-Readi, M.Z. The carotenoid fucoxanthin can sensitize multidrug resistant cancer cells to doxorubicin via induction of apoptosis, inhibition of multidrug resistance proteins and metabolic enzymes. Phytomedicine 2020, 77, 153280. [Google Scholar] [CrossRef]
- De la Mare, J.A.; Sterrenberg, J.N.; Sukhthankar, M.G.; Chiwakata, M.T.; Beukes, D.R.; Blatch, G.L.; Edkins, A.L. Assessment of potential anti-cancer stem cell activity of marine algal compounds using an in vitro mammosphere assay. Cancer Cell Int. 2013, 13, 39. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, A.S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Terasaki, M.; Takahashi, S.; Nishimura, R.; Kubota, A.; Kojima, H.; Ohta, T.; Hamada, J.; Kuramitsu, Y.; Maeda, H.; Miyashita, K.; et al. A marine carotenoid of fucoxanthinol accelerates the growth of human pancreatic cancer PANC-1 cells. Nutr. Cancer 2022, 74, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Inoue, T.; Murase, W.; Kubota, A.; Kojima, H.; Kojoma, M.; Ohta, T.; Maeda, H.; Miyashita, K.; Mutoh, M.; et al. A fucoxanthinol induces apoptosis in a pancreatic intraepithelial neoplasia cell. Cancer Genom. Proteom. 2021, 18, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Nishizaka, Y.; Murase, W.; Kubota, A.; Kojima, H.; Kojoma, M.; Tanaka, T.; Maeda, H.; Miyashita, K.; Mutoh, M.; et al. Effect of fucoxanthinol on pancreatic ductal adenocarcinoma cells from an N-nitrosobis(2-oxopropyl)amine-initiated syrian golden hamster pancreatic carcinogenesis model. Cancer Genom. Proteom. 2021, 18 (Suppl. 3), 407–423. [Google Scholar] [CrossRef]
- Govaere, O.; Petz, M.; Wouters, J.; Vandewynchel, Y.P.; Scott, E.J.; Topal, B.; Nevens, F.; Verslype, C.; Anstee, Q.M.; Van Vilerberghe, H.; et al. The PDGFR-α-laminin B1-keratin 19 cascade drives tumor progression at the invasive front of human hepatocellular carcinoma. Oncogene 2017, 36, 6605–6616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petz, M.; Kozina, D.; Huber, H.; Siwiec, T.; Seipelt, J.; Sommergruber, W.; Mikulits, W. The leader region of Laminin B1 mRNA confers cap-independent translation. Nucleic Acids Res. 2007, 35, 2473–2482. [Google Scholar] [CrossRef] [Green Version]
- Fujita, M.; Khazenzon, N.M.; Bose, S.; Sekiguchi, K.; Sasaki, T.; Carter, W.G.; Ljubimov, A.V.; Black, K.L.; Ljubimova, J.Y. Overexpression of beta1-chain-containing laminins in capillary basement memebranes of human breast cancer and its metastasis. Breast Cancer Res. 2005, 7, R411–R421. [Google Scholar] [CrossRef] [Green Version]
- Arimori, T.; Miyazaki, N.; Mihara, E.; Takizawa, M.; Taniguchi, Y.; Cabañas, C.; Sekiguchi, K.; Takagi, J. Structural mechanism of laminin recognition by integrin. Nat. Commun. 2021, 12, 4012. [Google Scholar] [CrossRef]
- Buchheit, C.L.; Weigel, K.J.; Schafer, Z.T. Cancer cell survival during detachment from the ECM: Multiple barriers to tumour progression. Nat. Rev. Cancer 2014, 14, 632–641. [Google Scholar] [CrossRef]
- Paoli, P.; Giannoni, E.; Chiarugi, P. Anoikis molecular pathways and its role in cancer progression. Biochim. Biophys. Acta 2013, 1833, 3481–3498. [Google Scholar] [CrossRef] [Green Version]
- Horbinski, C.; Mojesky, C.; Kyprianou, N. Live free or die: Tales of homeless (cells) in cancer. Am. J. Pathol. 2010, 177, 1044–1052. [Google Scholar] [CrossRef]
- Alday-Parejo, B.; Stupp, R.; Rüegg, C. Are integrins still practicable targets for anti-cancer therapy? Cancers 2019, 11, E978. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, R.; Satoh, R.; Takasaki, T. ERK: A double-edged sword in cancer. ERK-dependent apoptosis as a potential therapeutic strategy for cancer. Cells 2021, 10, 2509. [Google Scholar] [CrossRef]
- Wu, P.K.; Becker, A.; Park, J.I. Growth inhibitory signaling of the Raf/MEK/ERK pathway. Int. J. Mol. Sci. 2020, 21, 5436. [Google Scholar] [CrossRef]
- Struewing, J.P.; Hartge, P.; Wacholder, S.; Baker, S.M.; Berlin, M.; McAdams, M.; Timmerman, M.M.; Brody, L.C.; Tucker, M.A. The risk of cancer associated with specific mutations of BRCA1 and BRCA2 among Ashkenazi Jews. N. Engl. J. Med. 1997, 336, 1401–1408. [Google Scholar] [CrossRef]
- King, M.C.; Marks, J.H.; Mandell, J.B.; New York Breast Cancer Study Group. Breast and ovarian cancer risks due to inherited mutations in BRCA1 and BRCA2. Science 2003, 302, 643–646. [Google Scholar] [CrossRef]
- Warner, E.; Plewes, D.B.; Hill, K.A.; Causer, P.A.; Zubovits, J.T.; Jong, R.A.; Cutrara, M.R.; DeBoer, G.; Yaffe, M.J.; Messner, S.J.; et al. Surveillance of BRCA1 and BRCA2 mutation carriers with magnetic resonance imaging, ultrasound, mammography, and clinical breast examination. JAMA 2004, 292, 1317–1325. [Google Scholar] [CrossRef] [Green Version]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [Green Version]
- Sørlie, T.; Tibshirani, R.; Parker, J.; Hastie, T.; Marron, J.S.; Nobel, A.; Deng, S.; Johnsen, H.; Pesich, R.; Geisler, S.; et al. Repeated observation of breast tumor subtypes in independent gene expression data sets. Proc. Natl. Acad. Sci. USA 2003, 100, 8418–8423. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [Green Version]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [Green Version]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Morris, G.J.; Naidu, S.; Topham, A.; Guiles, F.; Xu, Y.; McCue, P.; Schwartz, G.F.; Park, P.K.; Rosenberg, A.L.; Brill, K.; et al. Differences in breast carcinoma characteristics in newly diagnosed African-American and Caucasian patients: A single-institution compilation compared with the National Cancer Institute’s Surveillance, Epidemiology, and Endo Results Database. Cancer 2007, 110, 876–884. [Google Scholar] [CrossRef]
- Gerratana, L.; Basile, D.; Buono, G.; De Placido, S.; Giuliano, M.; Minichillo, S.; Coinu, A.; Martorana, F.; De Santo, I.; Del Mastro, L.; et al. Androgen receptor in triple negative breast cancer: A potential target for the targetless subtype. Cancer Treat. Rev. 2018, 68, 102–110. [Google Scholar] [CrossRef]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast cancer cell line classification and its relevance with breast tumor subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [Green Version]
- Leroy, B.; Girard, L.; Hollestelle, A.; Minna, J.D.; Gazdar, A.F.; Soussi, T. Analysis of TP53 mutation status in human cancer cell lines: A reassessment. Hum. Mutat. 2014, 35, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Amirabadi, H.E.; Tuerlings, M.; Hollestelle, A.; SahebAli, S.; Luttge, R.; van Donkelaar, C.C.; Martens, J.W.M.; den Toonder, J.M.J. Characterizing the invasion of different breast cancer cell lines with distinct E-cadherin status in 3D using a microfluidic system. Biomed. Microdevices 2019, 21, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaki, M.; Maeda, H.; Miyashita, K.; Mutoh, M. Induction of anoikis in human colorectal cancer cells by fucoxanthinol. Nutr. Cancer 2017, 69, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, R.; Kojima, H.; Takai, R.; Ohta, T.; Maeda, H.; Miyashita, K.; Mutoh, M.; Terasaki, M. Effects of CLIC4 on fucoxanthinol-induced apoptosis in human colorectal cancer cells. Nutr. Cancer 2021, 73, 889–898. [Google Scholar] [CrossRef]
- Jung, H.; Lee, D.S.; Park, S.K.; Choi, J.S.; Jung, W.K.; Park, W.S.; Choi, I.W. Fucoxanthin inhibits myofibroblast differentiation and extracellular matrix production in nasal polyp-derived fibroblasts via modulation of Smad-dependent and Smad-independent signaling pathways. Mar. Drugs 2018, 16, 323. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG Pathway b | Size c | NES d | NOM e p-Value | FDR f q-Value |
|---|---|---|---|---|
| Enriched gene sets in control cells (group 2) | ||||
| Cell cycle g | 51 | 2.32 | 0.000 | 0.000 |
| DNA replication | 23 | 2.31 | 0.000 | 0.001 |
| Systemic lupus erythematosus | 41 | 2.24 | 0.000 | 0.002 |
| Valine leucine and isoleucine degradation | 23 | 2.11 | 0.000 | 0.009 |
| Calcium signaling pathway | 24 | 2.03 | 0.000 | 0.015 |
| Purine metabolism | 41 | 1.93 | 0.000 | 0.027 |
| Melanogenesis | 22 | 1.79 | 0.000 | 0.075 |
| Dilated cardiomyopathy | 17 | 1.78 | 0.008 | 0.071 |
| Propanoate metabolism | 15 | 1.75 | 0.008 | 0.074 |
| ECM receptor interaction | 15 | 1.72 | 0.023 | 0.083 |
| Enriched gene sets FxOH-treated cells (group 1) | ||||
| Cytokine–cytokine receptor interaction | 34 | 2.11 | 0.000 | 0.033 |
| Protein export | 15 | 2.02 | 0.008 | 0.033 |
| Nod-like receptor signaling pathway | 16 | 1.70 | 0.026 | 0.203 |
| MAPK signaling pathway | 60 | 1.64 | 0.012 | 0.212 |
| Amino sugar and nucleotide sugar metabolism | 15 | 1.58 | 0.061 | 0.253 |
| Spliceosome | 21 | 1.54 | 0.050 | 0.249 |
| Natural killer cell mediated cytotoxicity | 24 | 1.54 | 0.056 | 0.214 |
| Metabolism of xenobiotics by cytochrome P450 | 15 | 1.43 | 0.097 | 0.333 |
| Epithelial cell signaling in Helicobacter pylori | 20 | 1.41 | 0.111 | 0.322 |
| Rig I-like receptor signaling pathway | 15 | 1.28 | 0.199 | 0.506 |
| KEGG Pathway b | Size c | NES d | NOM e p-Value | FDR f q-Value |
|---|---|---|---|---|
| Enriched gene sets in control cells (group 2) | ||||
| Systemic lupus erythematosus | 37 | 2.72 | 0.000 | 0.000 |
| DNA replication g | 26 | 2.38 | 0.000 | 0.000 |
| Purine metabolism | 37 | 2.18 | 0.000 | 0.006 |
| Pyrimidine metabolism | 32 | 1.90 | 0.006 | 0.051 |
| Cell adhesion molecular (CAMS) | 21 | 1.86 | 0.004 | 0.055 |
| ECM–receptor interaction | 27 | 1.82 | 0.008 | 0.057 |
| Nucleotide excision repair | 15 | 1.82 | 0.008 | 0.049 |
| Base excision repair | 15 | 1.74 | 0.019 | 0.072 |
| Arrhythmogenic right ventricular cardiomyopathy (ARVC) | 15 | 1.58 | 0.054 | 0.160 |
| Homologous recombination | 15 | 1.53 | 0.055 | 0.188 |
| Enriched gene sets FxOH-treated cells (group 1) | ||||
| MAPK signaling pathway | 57 | 2.68 | 0.000 | 0.000 |
| Spliceosome | 17 | 2.12 | 0.002 | 0.012 |
| Epithelial cell signaling in Helicobacter pyrori infection | 16 | 2.00 | 0.004 | 0.020 |
| Cytokine–cytokine receptor interaction | 30 | 1.59 | 0.042 | 0.236 |
| Antigen processing and presentation | 16 | 1.55 | 0.051 | 0.230 |
| Insulin signaling pathway | 29 | 1.41 | 0.098 | 0.366 |
| Endocytosis | 28 | 1.37 | 0.134 | 0.379 |
| Toll-like receptor signaling pathway | 19 | 1.32 | 0.149 | 0.417 |
| T-cell receptor signaling pathway | 17 | 1.29 | 0.179 | 0.406 |
| B-cell signaling pathway | 16 | 1.25 | 0.191 | 0.444 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasuda, A.; Wagatsuma, M.; Murase, W.; Kubota, A.; Kojima, H.; Ohta, T.; Hamada, J.; Maeda, H.; Terasaki, M. Fucoxanthinol Promotes Apoptosis in MCF-7 and MDA-MB-231 Cells by Attenuating Laminins–Integrins Axis. Onco 2022, 2, 145-163. https://doi.org/10.3390/onco2030010
Yasuda A, Wagatsuma M, Murase W, Kubota A, Kojima H, Ohta T, Hamada J, Maeda H, Terasaki M. Fucoxanthinol Promotes Apoptosis in MCF-7 and MDA-MB-231 Cells by Attenuating Laminins–Integrins Axis. Onco. 2022; 2(3):145-163. https://doi.org/10.3390/onco2030010
Chicago/Turabian StyleYasuda, Ayaka, Momoka Wagatsuma, Wataru Murase, Atsuhito Kubota, Hiroyuki Kojima, Tohru Ohta, Junichi Hamada, Hayato Maeda, and Masaru Terasaki. 2022. "Fucoxanthinol Promotes Apoptosis in MCF-7 and MDA-MB-231 Cells by Attenuating Laminins–Integrins Axis" Onco 2, no. 3: 145-163. https://doi.org/10.3390/onco2030010
APA StyleYasuda, A., Wagatsuma, M., Murase, W., Kubota, A., Kojima, H., Ohta, T., Hamada, J., Maeda, H., & Terasaki, M. (2022). Fucoxanthinol Promotes Apoptosis in MCF-7 and MDA-MB-231 Cells by Attenuating Laminins–Integrins Axis. Onco, 2(3), 145-163. https://doi.org/10.3390/onco2030010