

Speed Breeding for Crop Improvement and Food Security


Abstract
:1. Introduction
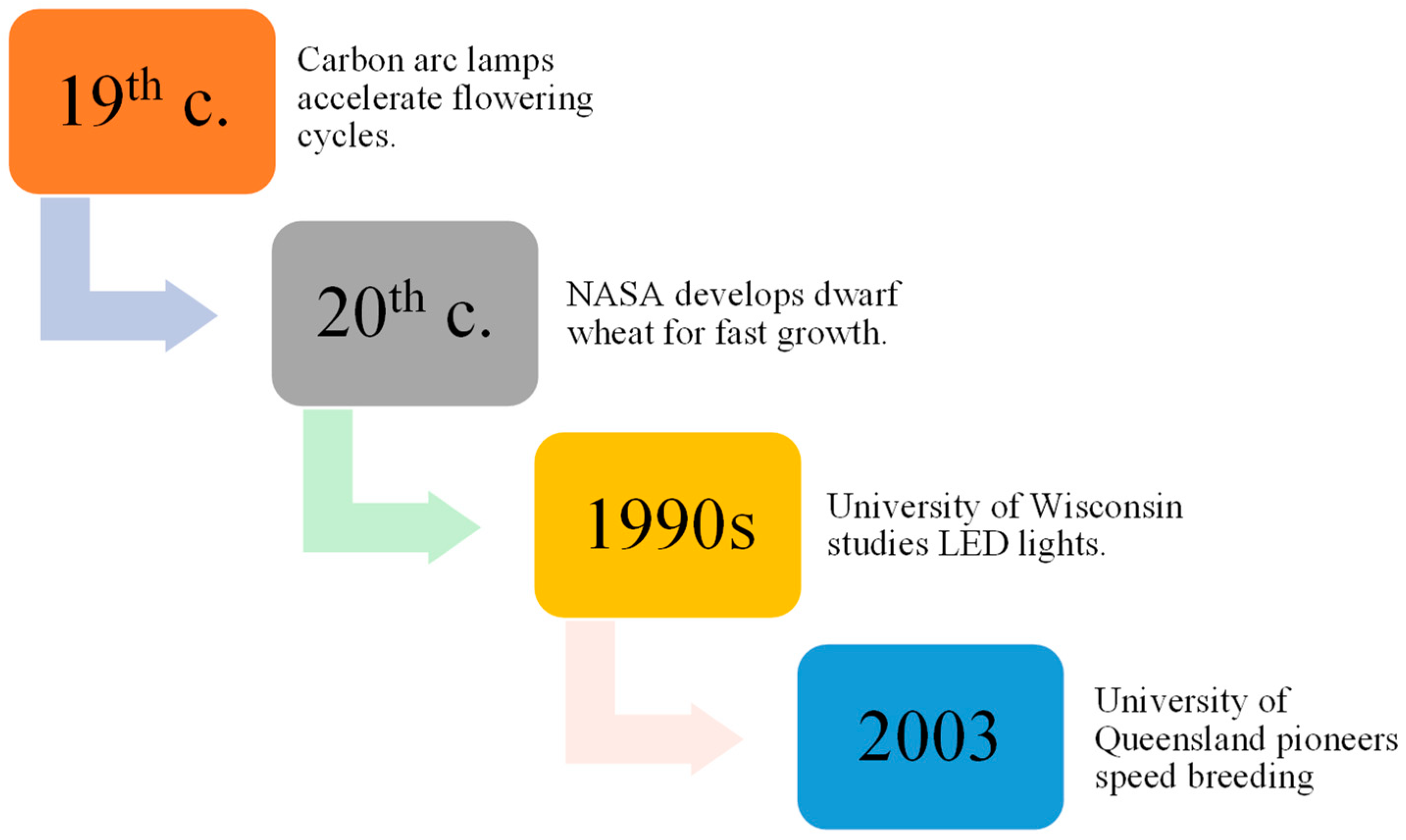
2. The History and Development of SB in Enhancing Crop Genetics
3. SB Applications and Selection Methods in Plant Breeding
4. Potential Advantages of SB Techniques
5. Applications of SB beyond Crop Improvement
5.1. Genetic Mapping Populations
5.2. Genetic Modification (GM) Crop Development
5.3. Trait Stacking for Resilient Crop Varieties
5.4. Enhancing Research across Disciplines
6. Limitations Associated with SB
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ahmar, S.; Gill, R.A.; Jung, K.-H.; Faheem, A.; Qasim, M.U.; Mubeen, M.; Zhou, W. Conventional and molecular techniques from simple breeding to speed breeding in crop plants: Recent advances and future outlook. Int. J. Mol. Sci. 2020, 21, 2590. [Google Scholar] [CrossRef] [PubMed]
- Andrus, C.F. Plant breeding systems. Euphytica 1963, 12, 205–228. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Zohaib, A.; Tanveer, M.; Naeem, M.; Ali, I.; Tabassum, T.; Nazir, U. Growth and developmental responses of crop plants under drought stress: A review. Zemdirb.-Agric. 2017, 104, 267–276. [Google Scholar] [CrossRef]
- Bailey, L.H. Some Preliminary Studies of the Influence of the Electric Arc Lamp upon Greenhouse Plants; Cornell University: Ithaca, NY, USA, 1891; Volume 30. [Google Scholar]
- Begna, T. Speed breeding to accelerate crop improvement. Int. J. Agric. Sc. Food Technol. 2022, 8, 178–186. [Google Scholar] [CrossRef]
- Bermejo, C.; Gatti, I.; Cointry, E. In vitro embryo culture to shorten the breeding cycle in lentil (Lens culinaris Medik). Plant Cell Tiss. Organ. Cult. 2016, 127, 585–590. [Google Scholar] [CrossRef]
- Bhatta, M.; Sandro, P.; Smith, M.R.; Delaney, O.; Voss-Fels, K.P.; Gutierrez, L.; Hickey, L.T. Need for speed: Manipulating plant growth to accelerate breeding cycles. Curr. Opin. Plant Biol. 2021, 60, 101986. [Google Scholar] [CrossRef]
- Bonea, D. Speed breeding and its importance for the improvement of agricultural crops. AAMC 2022, 52, 59–66. [Google Scholar] [CrossRef]
- Bugbee, B.; Koerner, G. Yield comparisons and unique characteristics of the dwarf wheat cultivar ‘USU-Apogee’. Adv. Space Res. 1997, 20, 1891–1894. [Google Scholar] [CrossRef]
- Byerlee, D.; Fischer, K. Accessing modern science: Policy and institutional options for agricultural biotechnology in developing countries. World Dev. 2002, 30, 931–948. [Google Scholar] [CrossRef]
- Cazzola, F.; Bermejo, C.J.; Guindon, M.F.; Cointry, E. Speed breeding in pea (Pisum sativum L.), an efficient and simple system to accelerate breeding programs. Euphytica 2020, 216, 178. [Google Scholar] [CrossRef]
- Chiurugwi, T.; Kemp, S.; Powell, W.; Hickey, L.T. Speed breeding orphan crops. Theor. Appl. Genet. 2019, 132, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Phil. Trans. R. Soc. B 2014, 369, 20130243. [Google Scholar] [CrossRef]
- Ferrie, A.M.R. Doubled haploid production in nutraceutical species: A review. Euphytica 2007, 158, 347–357. [Google Scholar] [CrossRef]
- Gaba, Y.; Pareek, A.; Singla-Pareek, S.L. Raising climate-resilient crops: Journey from the conventional breeding to new breeding approaches. Curr. Genom. 2021, 22, 450–467. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Watson, A.; Gonzalez-Navarro, O.E.; Ramirez-Gonzalez, R.H.; Yanes, L.; Mendoza-Suárez, M.; Simmonds, J.; Wells, R.; Rayner, T.; Green, P.; et al. Speed breeding in growth chambers and glasshouses for crop breeding and model plant research. Nat. Protoc. 2018, 13, 2944–2963. [Google Scholar] [CrossRef]
- Gilland, B. World population and food supply. Food Policy 2002, 27, 47–63. [Google Scholar] [CrossRef]
- Gray, S.B.; Brady, S.M. Plant developmental responses to climate change. Dev. Biol. 2016, 419, 64–77. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather. Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Hickey, L.T.; Germán, S.E.; Pereyra, S.A.; Diaz, J.E.; Ziems, L.A.; Fowler, R.A.; Platz, G.J.; Franckowiak, J.D.; Dieters, M.J. Speed breeding for multiple disease resistance in barley. Euphytica 2017, 213, 64. [Google Scholar] [CrossRef]
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.M.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B.H. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef]
- Houston, R.D.; Bean, T.P.; Macqueen, D.J.; Gundappa, M.K.; Jin, Y.H.; Jenkins, T.L.; Selly, S.L.C.; Martin, S.A.M.; Stevens, J.R.; Santos, E.M.; et al. Harnessing genomics to fast-track genetic improvement in aquaculture. Nat. Rev. Genet. 2020, 21, 389–409. [Google Scholar] [CrossRef] [PubMed]
- Hussain, K.; Mahrukh; Nisa, R.T.; Zaid, A.; Mushtaq, M. The utilization of speed breeding and genome editing to achieve zero hunger. In Sustainable Agriculture in the Era of the OMICs Revolution; Prakash, C.S., Fiaz, S., Nadeem, M.A., Baloch, F.S., Qayyum, A., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 1–15. ISBN 978-3-031-15567-3. [Google Scholar]
- Jacquier, N.M.; Gilles, L.M.; Martinant, J.-P.; Rogowsky, P.M.; Widiez, T. Maize in Planta haploid inducer lines: A cornerstone for doubled haploid technology. In Doubled Haploid Technology; Volume 2: Hot Topics, Apiaceae, Brassicaceae, Solanaceae; Humana: New York, NY, USA, 2021; Volume 2288, pp. 25–48. [Google Scholar] [CrossRef]
- Bula, R.J.; Morrow, R.C.; Tibbitts, T.; Barta, D.; Ignatius, R.; Martin, T. Light-emitting diodes as a radiation source for plants. HortScience 1991, 26, 203–205. [Google Scholar] [CrossRef]
- Hoenecke, M.; Bula, R.; Tibbitts, T. Importance of blue photon levels for lettuce seedlings grown under red-light-emitting diodes. HortScience 1992, 27, 427–430. [Google Scholar] [CrossRef]
- Zhou, W. Advanced ASTROCULTURETM Plant Growth Unit: Capabilities and Performances; SAE Technical Paper; SAE International: Warrendale, PA, USA, 2005. [Google Scholar] [CrossRef]
- Morrow, R.C.; Duffie, N.A.; Tibbitts, T.W.; Bula, R.J.; Barta, D.J.; Ming, D.W.; Wheeler, R.M.; Porterfield, D.M. Plant Response in the ASTROCULTURETM Flight Experiment Unit; SAE Technical Paper; SAE International: Warrendale, PA, USA, 1995. [Google Scholar] [CrossRef]
- Link, B.; Durst, S.; Zhou, W.; Stankovic, B. Seed-to-seed growth of Arabidopsis thaliana on the International Space Station. Adv. Space Res. 2003, 31, 2237–2243. [Google Scholar] [CrossRef]
- Croxdale, J.; Cook, M.; Tibbitts, T.W.; Brown, C.S.; Wheeler, R.M. Structure of Potato tubers formed during spaceflight. J. Exp. Bot. 1997, 48, 2037–2043. [Google Scholar] [CrossRef]
- Wu, B.-S.; Mansoori, M.; Trumpler, K.; Addo, P.W.; MacPherson, S.; Lefsrud, M. Effect of amber (595 Nm) light supplemented with narrow blue (430 Nm) light on tomato biomass. Plants 2023, 12, 2457. [Google Scholar] [CrossRef]
- Guo, X.; Xue, X.; Chen, L.; Li, J.; Wang, Z.; Zhang, Y. Effects of LEDs light spectra on the growth, yield, and quality of winter wheat (Triticum aestivum L.) cultured in plant factory. J. Plant Growth Regul. 2023, 42, 2530–2544. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Bahuguna, R.N.; Djanaguiraman, M.; Gamuyao, R.; Prasad, P.V.V.; Craufurd, P.Q. Implications of high temperature and elevated CO2 on flowering time in plants. Front. Plant Sci. 2016, 7, 913. [Google Scholar] [CrossRef]
- Jähne, F.; Hahn, V.; Würschum, T.; Leiser, W.L. Speed breeding short-day crops by LED-controlled light schemes. Theor. Appl. Genet. 2020, 133, 2335–2342. [Google Scholar] [CrossRef]
- Croser, J.S.; Pazos-Navarro, M.; Bennett, R.G.; Tschirren, S.; Edwards, K.; Erskine, W.; Creasy, R.; Ribalta, F.M. Time to flowering of temperate pulses in vivo and generation turnover in vivo–in vitro of narrow-leaf lupin accelerated by low red to far-red ratio and high intensity in the far-red region. Plant Cell Tiss. Organ. Cult. 2016, 127, 591–599. [Google Scholar] [CrossRef]
- O’Connor, D.J.; Wright, G.C.; Dieters, M.J.; George, D.L.; Hunter, M.N.; Tatnell, J.R.; Fleischfresser, D.B. Development and application of speed breeding technologies in a commercial peanut breeding program. Peanut Sci. 2013, 40, 107–114. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, K.; Shah, M.P.; Singh, A.K.; Kumar, A.; Kumar, Y. Application of omics technologies for microbial community structure and function analysis in contaminated environment. In Wastewater Treatment: Cutting-Edge Molecular Tools, Techniques and Applied Aspects; Shah, M.P., Sarkar, A., Mandal, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–40. [Google Scholar] [CrossRef]
- Mao, H.; Hang, T.; Zhang, X.; Lu, N. Both multi-segment light intensity and extended photoperiod lighting strategies, with the same daily light integral, promoted Lactuca sativa L. growth and photosynthesis. Agronomy 2019, 9, 857. [Google Scholar] [CrossRef]
- Alahmad, S.; Dinglasan, E.; Leung, K.M.; Riaz, A.; Derbal, N.; Voss-Fels, K.P.; Able, J.A.; Bassi, F.M.; Christopher, J.; Hickey, L.T. Speed breeding for multiple quantitative traits in durum wheat. Plant Methods 2018, 14, 36. [Google Scholar] [CrossRef] [PubMed]
- Gudi, S.; Kumar, P.; Singh, S.; Tanin, M.J.; Sharma, A. Strategies for accelerating genetic gains in crop plants: Special focus on speed breeding. Physiol. Mol. Biol. Plants 2022, 28, 1921–1938. [Google Scholar] [CrossRef] [PubMed]
- Shariatpanahi, M.E.; Niazian, M.; Ahmadi, B. Methods for chromosome doubling. Methods Mol. Biol. 2021, 2287, 127–148. [Google Scholar]
- Melchinger, A.E.; Molenaar, W.S.; Mirdita, V.; Schipprack, W. Colchicine alternatives for chromosome doubling in maize haploids for doubled-haploid production. Crop Sci. 2016, 56, 559–569. [Google Scholar] [CrossRef]
- Watts, A.; Sankaranarayanan, S.; Raipuria, R.K.; Watts, A. Production and application of doubled haploid in brassica improvement. In Brassica Improvement; Wani, S., Thakur, A., Jeshima Khan, Y., Eds.; Springer: Cham, Switzerland, 2020; pp. 67–84. [Google Scholar] [CrossRef]
- Croser, J.S.; Lülsdorf, M.M.; Davies, P.A.; Clarke, H.J.; Bayliss, K.L.; Mallikarjuna, N.; Siddique, K.H.M. Toward doubled haploid production in the Fabaceae: Progress, constraints, and opportunities. Crit. Rev. Plant Sci. 2006, 25, 139–157. [Google Scholar] [CrossRef]
- Forster, B.P.; Heberle-Bors, E.; Kasha, K.J.; Touraev, A. The resurgence of haploids in higher plants. Trends Plant Sci. 2007, 12, 368–375. [Google Scholar] [CrossRef]
- Seguí-Simarro, J.M.; Corral-Martínez, P.; Parra-Vega, V.; González-García, B. Androgenesis in recalcitrant Solanaceous crops. Plant Cell Rep. 2011, 30, 765–778. [Google Scholar] [CrossRef]
- Dong, Y.-Q.; Zhao, W.-X.; Li, X.-H.; Liu, X.-C.; Gao, N.-N.; Huang, J.-H.; Wang, W.-Y.; Xu, X.-L.; Tang, Z.-H. Androgenesis, gynogenesis, and parthenogenesis haploids in cucurbit species. Plant Cell Rep. 2016, 35, 1991–2019. [Google Scholar] [CrossRef]
- Massel, K.; Lam, Y.; Wong, A.C.; Hickey, L.T.; Borrell, A.K.; Godwin, I.D. Hotter, Drier, CRISPR: The latest edit on climate change. Theor. Appl. Genet. 2021, 134, 1691–1709. [Google Scholar] [CrossRef]
- Mobini, S.H.; Warkentin, T.D. A simple and efficient method of in vivo rapid generation technology in pea (Pisum sativum L.). In Vitro Cell Dev. Biol.-Plant 2016, 52, 530–536. [Google Scholar] [CrossRef]
- Naqvi, R.Z.; Siddiqui, H.A.; Mahmood, M.A.; Najeebullah, S.; Ehsan, A.; Azhar, M.; Farooq, M.; Amin, I.; Asad, S.; Mukhtar, Z.; et al. Smart breeding approaches in post-genomics era for developing climate-resilient food crops. Front. Plant Sci. 2022, 13, 972164. [Google Scholar] [CrossRef]
- Pandey, S.; Singh, A.; Parida, S.K.; Prasad, M. Combining speed breeding with traditional and genomics-assisted breeding for crop improvement. Plant Breed. 2022, 141, 301–313. [Google Scholar] [CrossRef]
- Pfeiffer, N.E. Microchemical and morphological studies of effect of light on plants. Bot. Gaz. 1926, 81, 173–195. [Google Scholar] [CrossRef]
- Radha, T.; Mathew, L. Fruit Crops; New India Publishing: New Delhi, India, 2007; Volume 3. [Google Scholar]
- Rajan, S.; Singh, G. Approaches and Strategies for Precision Farming in Mango. In Precision Farming in Horticulture; Singh, H.P., Singh, G., Samuel, J.C., Pathak, R.K., Eds.; Central Institute of Subtropical Horticulture: Lucknow, India, 2003; pp. 124–144. [Google Scholar]
- Shamshiri, R.R.; Kalantari, F.; Ting, K.C.; Thorp, K.R.; Hameed, I.A.; Weltzien, C.; Ahmad, D.; Shad, Z.M. Advances in greenhouse automation and controlled environment agriculture: A transition to plant factories and urban agriculture. Int. J. Agric. Biol. Eng. 2018, 11, 1–22. [Google Scholar] [CrossRef]
- Riaz, A.; Athiyannan, N.; Periyannan, S.; Afanasenko, O.; Mitrofanova, O.; Aitken, E.A.B.; Lagudah, E.; Hickey, L.T. Mining Vavilov’s treasure chest of wheat diversity for adult plant resistance to Puccinia triticina. Plant Dis. 2017, 101, 317–323. [Google Scholar] [CrossRef]
- Ribalta, F.M.; Pazos-Navarro, M.; Nelson, K.; Edwards, K.; Ross, J.J.; Bennett, R.G.; Munday, C.; Erskine, W.; Ochatt, S.J.; Croser, J.S. Precocious floral initiation and identification of exact timing of embryo physiological maturity facilitate germination of immature seeds to truncate the lifecycle of pea. Plant Growth Regul. 2017, 81, 345–353. [Google Scholar] [CrossRef]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2021, 44, 645–664. [Google Scholar] [CrossRef] [PubMed]
- Samantara, K.; Bohra, A.; Mohapatra, S.R.; Prihatini, R.; Asibe, F.; Singh, L.; Reyes, V.P.; Tiwari, A.; Maurya, A.K.; Croser, J.S.; et al. Breeding more crops in less time: A perspective on speed breeding. Biology 2022, 11, 275. [Google Scholar] [CrossRef]
- Tripp, R.; Louwaars, N.; Eaton, D. Plant variety protection in developing countries. a report from the field. Food Policy 2007, 32, 354–371. [Google Scholar] [CrossRef]
- Samineni, S.; Sen, M.; Sajja, S.B.; Gaur, P.M. Rapid generation advance (RGA) in Chickpea to produce up to seven generations per year and enable speed breeding. Crop J. 2020, 8, 164–169. [Google Scholar] [CrossRef]
- Shahzad, A.; Ullah, S.; Dar, A.A.; Sardar, M.F.; Mehmood, T.; Tufail, M.A.; Shakoor, A.; Haris, M. Nexus on climate change: Agriculture and possible solution to cope future climate change stresses. Environ. Sci. Pollut. Res. 2021, 28, 14211–14232. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kumar, A.; Dhakte, P.; Raturi, G.; Vishwakarma, G.; Barbadikar, K.M.; Das, B.K.; Shivaraj, S.M.; Sonah, H.; Deshmukh, R. Speed breeding opportunities and challenges for crop improvement. J. Plant Growth Regul. 2023, 42, 46–59. [Google Scholar] [CrossRef]
- Slafer, G.A.; Rawson, H.M. Phyllochron in wheat as affected by photoperiod under two temperature regimes. Funct. Plant Biol. 1997, 24, 151–158. [Google Scholar] [CrossRef]
- Shaw, H.J. The Consuming Geographies of Food: Diet, Food Deserts and Obesity; Routledge: London, UK, 2014; p. 210. [Google Scholar]
- Sinha, P.; Singh, V.K.; Bohra, A.; Kumar, A.; Reif, J.C.; Varshney, R.K. Genomics and breeding innovations for enhancing genetic gain for climate resilience and nutrition traits. Theor. Appl. Genet. 2021, 134, 1829–1843. [Google Scholar] [CrossRef]
- Stam, P. Construction of Integrated genetic linkage maps by means of a new computer package: Join Map. Plant J. 1993, 3, 739–744. [Google Scholar] [CrossRef]
- Steinwand, M.A.; Ronald, P.C. Crop biotechnology and the future of food. Nat. Food 2020, 1, 273–283. [Google Scholar] [CrossRef]
- Stetter, M.G.; Zeitler, L.; Steinhaus, A.; Kroener, K.; Biljecki, M.; Schmid, K.J. Crossing methods and cultivation conditions for rapid production of segregating populations in three grain Amaranth species. Front. Plant Sci. 2016, 7, 816. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J.; Biswas, S.; Tsakirpaloglou, N.; Septiningsih, E.M. Functional allele validation by gene editing to leverage the wealth of genetic resources for crop improvement. Int. J. Mol. Sci. 2022, 23, 6565. [Google Scholar] [CrossRef]
- Schneider, K. Mapping populations and principles of genetic mapping. In The Handbook of Plant Genome Mapping: Genetic and Physical Mapping; Meksem, K., Kahl, G., Eds.; Wiley-VCH: Weinheim, Germany, 2005; pp. 1–21. [Google Scholar] [CrossRef]
- McClung, C.R.; Lou, P.; Hermand, V.; Kim, J.A. The importance of ambient temperature to growth and the induction of flowering. Front. Plant Sci. 2016, 7, 1266. [Google Scholar] [CrossRef]
- Wanga, M.A.; Shimelis, H.; Mashilo, J.; Laing, M.D. Opportunities and challenges of speed breeding: A review. Plant Breed. 2021, 140, 185–194. [Google Scholar] [CrossRef]
- Dupuis, I.; Dumas, C. Influence of temperature stress on in vitro fertilization and heat shock protein synthesis in maize (Zea mays L.) reproductive tissues. Plant Physiol. 1990, 94, 665–670. [Google Scholar] [CrossRef]
- Kim, H.Y.; Horie, T.; Nakagawa, H.; Wada, K. Effects of elevated CO2 concentration and high temperature on growth and yield of rice: II. The effect on yield and its components of Akihikari rice. Jpn. J. Crop Sci. 1996, 65, 644–651. [Google Scholar] [CrossRef]
- Vadez, V.; Hash, T.; Bidinger, F.R.; Kholova, J. Phenotyping pearl millet for adaptation to drought. Front. Physiol. 2012, 3, 386. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; De Groot, S.; Soole, K.; Langridge, P. Early Flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.-D.; Hatta, M.A.M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding: A powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhang, P.; Wang, H.; Lu, Z.; Liu, C.J.; Liu, H.; Yan, G.J. How to advance up to seven generations of canola (Brassica napus L.) per annum for the production of pure line populations? Euphytica 2016, 209, 113–119. [Google Scholar] [CrossRef]
- Zheng, Z.; Wang, H.B.; Chen, G.D.; Yan, G.J.; Liu, C.J. A procedure allowing up to eight generations of wheat and nine generations of barley per annum. Euphytica 2013, 191, 311–316. [Google Scholar] [CrossRef]
- Schoen, A.; Wallace, S.; Holbert, M.F.; Brown-Guidera, G.; Harrison, S.; Murphy, P.; Sanantonio, N.; Van Sanford, D.; Boyles, R.; Mergoum, M.; et al. Reducing the generation time in winter wheat cultivars using speed breeding. Crop Sci. 2023, 63, 2079–2090. [Google Scholar] [CrossRef]
- Mobini, S.; Khazaei, H.; Warkentin, T.D.; Vandenberg, A. Shortening the generation cycle in Faba bean (Vicia faba) by application of cytokinin and cold stress to assist speed breeding. Plant Breed. 2020, 139, 1181–1189. [Google Scholar] [CrossRef]
- Lulsdorf, M.M.; Banniza, S. Rapid Generation cycling of an F2 population derived from a cross between Lens vulinaris Medik. and Lens ervoides (Brign.) grande after aphanomyces root rot selection. Plant Breed. 2018, 137, 486–491. [Google Scholar] [CrossRef]
- Liu, H.; Zwer, P.; Wang, H.; Liu, C.; Lu, Z.; Wang, Y.; Yan, G. A fast generation cycling system for oat and triticale breeding. Plant Breed. 2016, 135, 574–579. [Google Scholar] [CrossRef]
- González-Barrios, P.; Bhatta, M.; Halley, M.; Sandro, P.; Gutiérrez, L. Speed breeding and early panicle harvest accelerates oat (Avena sativa L.) breeding cycles. Crop Sci. 2021, 61, 320–330. [Google Scholar] [CrossRef]
- Edet, O.U.; Ishii, T. Cowpea speed breeding using regulated growth chamber conditions and seeds of oven-dried immature pods potentially accommodates eight generations per year. Plant Methods 2022, 18, 106. [Google Scholar] [CrossRef]
- Saxena, K.; Saxena, R.; Hickey, L.; Varshney, R. Can a speed breeding approach accelerate genetic gain in pigeonpea? Euphytica 2019, 215, 1–7. [Google Scholar] [CrossRef]
- Trnka, M.; Feng, S.; Semenov, M.A.; Olesen, J.E.; Kersebaum, K.C.; Rötter, R.P.; Semerádová, D.; Klem, K.; Huang, W.; Ruiz-Ramos, M.; et al. Mitigation efforts will not fully alleviate the increase in water scarcity occurrence probability in wheat-producing areas. Sci. Adv. 2019, 5, eaau2406. [Google Scholar] [CrossRef]
- Uğur, T.; GÖREN, H.K. Speed breeding: An innovative approach for accelerated genetic improvement in agricultural crops and integrating with molecular-based approaches. In Pioneer and Contemporary Studies in Agriculture, Forest and Water Issues; Duvar Yayınları: Izmir, Turkey, 2023; pp. 241–264. [Google Scholar] [CrossRef]
- Christopher, J.; Richard, C.; Chenu, K.; Christopher, M.; Borrell, A.; Hickey, L. Integrating rapid phenotyping and speed breeding to improve stay-green and root adaptation of wheat in changing, water-limited, Australian environments. Procedia Environ. Sci. 2015, 29, 175–176. [Google Scholar] [CrossRef]
- Rana, M.M.; Takamatsu, T.; Baslam, M.; Kaneko, K.; Itoh, K.; Harada, N.; Sugiyama, T.; Ohnishi, T.; Kinoshita, T.; Takagi, H.; et al. Salt tolerance improvement in rice through efficient SNP marker-assisted selection coupled with speed-breeding. Int. J. Mol. Sci. 2019, 20, 2585. [Google Scholar] [CrossRef]
- Varshney, R.K.; Bohra, A.; Yu, J.; Graner, A.; Zhang, Q.; Sorrells, M.E. Designing future crops: Genomics-assisted breeding comes of age. Trends Plant Sci. 2021, 26, 631–649. [Google Scholar] [CrossRef]
- Voss-Fels, K.P.; Cooper, M.; Hayes, B.J. Accelerating crop genetic gains with genomic selection. Theor. Appl. Genet. 2019, 132, 669–686. [Google Scholar] [CrossRef]
- Waltz, E. CRISPR-edited crops free to enter market, skip regulation. Nat. Biotechnol. 2016, 34, 582–583. [Google Scholar] [CrossRef] [PubMed]
- Eş, I.; Gavahian, M.; Marti-Quijal, F.J.; Lorenzo, J.M.; Khaneghah, A.M.; Tsatsanis, C.; Kampranis, S.C.; Barba, F.J. The application of the CRISPR-Cas9 Genome editing machinery in food and agricultural science: Current status, future perspectives, and associated challenges. Biotechnol. Adv. 2019, 37, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Yang, Y.; Qin, R.; Li, H.; Qiu, C.; Li, L.; Wei, P.; Yang, J. Rapid improvement of grain weight via highly efficient CRISPR/Cas9-mediated multiplex genome editing in rice. J. Genet. Genom. 2016, 43, 529–532. [Google Scholar] [CrossRef]
- Shailani, A.; Joshi, R.; Singla-Pareek, S.L.; Pareek, A. Stacking for future: Pyramiding genes to improve drought and salinity tolerance in rice. Physiol. Plant 2021, 172, 1352–1362. [Google Scholar] [CrossRef]
- Wenden, B.; Rameau, C. Systems biology for plant breeding: The example of flowering time in pea. Comptes Rendus Biol. 2009, 332, 998–1006. [Google Scholar] [CrossRef]
- Foucher, F.; Morin, J.; Courtiade, J.; Cadioux, S.; Ellis, N.; Banfield, M.J.; Rameau, C. Determinate and Late Flowering are two Terminal Flower1/Centroradialis homologs that control two distinct phases of flowering initiation and development in pea. Plant Cell 2003, 15, 2742–2754. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Knowles, C.L.; Vander Schoor, J.K.; Liew, L.C.; Jones, S.E.; Lambert, M.J.; Weller, J.L. Pea LATE BLOOMER1 is a GIGANTEA ortholog with roles in photoperiodic flowering, deetiolation, and transcriptional regulation of circadian clock gene homologs. Plant Physiol. 2007, 144, 648–661. [Google Scholar] [CrossRef]
- Warnasooriya, S.N.; Brutnell, T.P. Enhancing the productivity of grasses under high-density planting by engineering light responses: From model systems to feedstocks. J. Exp. Bot. 2014, 65, 2825–2834. [Google Scholar] [CrossRef]
- FAO. Water at a Glance—The Relationship between Water, Agriculture, Food Security and Poverty; FAO: Rome, Italy, 2008. [Google Scholar]
- Srivastav, A.L.; Dhyani, R.; Ranjan, M.; Madhav, S.; Sillanpää, M. Climate-resilient strategies for sustainable management of water resources and agriculture. Environ. Sci. Pollut. Res. 2021, 28, 41576–41595. [Google Scholar] [CrossRef] [PubMed]
- United Nations Conference on Trade and Development May. 2011. Available online: https://unctad.org/system/files/official-document/tdr2011_en.pdf (accessed on 10 September 2023).
- Saenz, A. Transforming Shipping Containers into Local Farms—Podponics Brings Produce to the City. Singularity Hub. 2011. Available online: https://singularityhub.com/2011/08/30/transforming-shipping-containers-into-local-farms-podponics-brings-produce-to-the-city/#sm.000100v3z66e9fdyt062k15j3s238 (accessed on 10 September 2023).
- CropBox. Available online: https://cropbox.co/ (accessed on 10 September 2023).
- Ribaut, J.; De Vicente, M.; Delannay, X. Molecular breeding in developing countries: Challenges and perspectives. Curr. Opin. Plant Biol. 2010, 13, 213–218. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Aspect | Description | Advantages | Limitations |
|---|---|---|---|
| Modification of Variables | Modification of key growth variables such as photoperiod, temperature, soil moisture, population density, and CO2 levels [55,56]. | Facilitates multiple breeding cycles annually, expedites breeding processes, and assesses populations in diverse environments [57,58]. Enhances genetic gain by optimizing conditions for crop development. | Lack of advanced controlled environment facilities can raise costs and limit accessibility, especially in resource-poor nations. Difficulty in maintaining steady electricity supply and temperature [11,60]. |
| Photoperiod Modification | Adjustment of photoperiod regimes to induce early blooming in crops like wheat, barley, chickpeas, and grain amaranth [11,24]. | Enables early flowering and seed set, suitable for regions with unstable electricity supplies. Utilizes low-energy LED and solar power systems for photoperiod control. | Excessive photoperiod can slow down plant growth and elevate stress hormone levels, requiring precise management Need for validation of field crop phenotyping due to potential bias in controlled environments. |
| Temperature Regulation | Modulation of soil and air temperatures to optimize plant progression and maturation [59,61]. | Promotes favorable timing of key growth transitions, vital for seed set, maturity, and flowering. Innovative technologies, like solar-powered air-conditioning, offer cost-effective solutions [62]. | Growing plants beyond their physiological limits can be harmful and result in genetic loss. Maintenance of temperature within specific limits is crucial. |
| Soil Moisture Management | Effective management of soil moisture, reducing it after flowering to facilitate rapid grain filling and maturation [3]. | Versatile strategies adaptable to various settings, whether open field or controlled environments. Indoor cultivation provides precise control of moisture levels [63]. | Challenges in controlling pests and diseases, tracking individuals for gene discovery purposes. |
| HDP | Low-cost strategy involving higher plant density, promoting early flowering, faster maturation, and reduced crop cycle length [64,65,66]. | Accelerates breeding processes, reduces cycle length, and maintains large populations for advanced selections. Valuable tool in modern agriculture. | Varying responses in different crop species and genotypes to modified CO2 levels. Requirements for appropriate facilities and safety protocols [67,68]. |
| Other Acceleration Methods | Methods like plant nutrition, hormonal therapies, organ tissue culture, drying, and chilling of seeds used to expedite development and trigger flowering [10,69]. | Offers additional tools for accelerating the breeding process and enhancing genetic gain. Diverse responses to plant growth regulators in controlled environments [70]. | Risk of genetic loss due to challenging management of these accelerated methods. |
| Crop | Speed Breeding Conditions | Generation Time | Number of Generations per Year | Reference |
|---|---|---|---|---|
| Soybean | 10 h. photoperiod (Blue light enriched) and use of light-emitting diodes (LEDs). | 77 | 5 | [34] |
| Peanut | 24 h. light and 28 ± 3 °C max. 17 ± 3 °C min. temperature. | 89 | 4 | [36] |
| Pea | 20 h. photoperiod, 21 °C/16 °C light/dark temp., 500 μM m−2 s−1 light intensity and hydroponic system. | 68.4 | 5.3 | [49] |
| Chickpea | 22 h. light, (25 ± 1) °C temperature and immature seed harvest. | 50–52.7 in early maturing accessions 55.4–58.6 in medium maturity accessions | 7, 6.2 and 6 in early, medium, and late maturity accessions | [61] |
| Wheat | 22 h. light, 22/17 °C temperature and immature seed harvest. | 65.4 | 5.6 | [78] |
| Canola | 22 h. light, 22/17 °C temperature and immature seed harvest. | 98.2 | 3.7 | [78] |
| Barley | 22 h. light, 22/17 °C temperature and immature seed harvest. | 68.4 | 5.3 | [78] |
| Canola | 20 h. photoperiod, 25/22 (±1) °C. | 62–71 | 5.1–5.9 | [79] |
| Winter wheat | 22 h. light, 25 °C/22 °C temperature. | 87 | 4 | [81] |
| Faba bean | 20 h. photoperiod, 21 °C light/16 °C dark temperature, 10–5 M BAP application. | 89 | 4 | [82] |
| Lentil | 20 h. photoperiod, 100 µM gibberellin application and immature seed harvest. | 56 | 5 | [83] |
| Oat, Triticale | 20 h. photoperiod, 25/22 °C day/night temperature, 65/85% day/night RH, In vitro culture of immature embryos. | 41–61 | 6–7.6 | [84] |
| Oat | 22 h. photoperiod, 20/16 °C. | 51 | 5 | [85] |
| Cowpea | 11-day-old pod seeds oven-dried at 39 °C. | - | 8 | [86] |
| Pigeon pea | 2870–2900 rpm and a blower wheel delivering air @ 2980–9330 m3/h. 28–32 °C. | 50–56 | 4 | [87] |
| Aspect | Description | Advantages | Limitations |
|---|---|---|---|
| Diverse Mapping Populations | Efficient creation of genetically diverse populations. | Facilitates the development of crop varieties with broad genetic diversity. | Genotypic variations in plant species may lead to challenges in ensuring the stability and uniformity of crop performance across different environments. |
| GM Crop Development | Shortened timelines for GM crop creation. | Accelerates the development of genetically modified crop varieties with desired traits. | Ethical considerations related to GM crops, including safety and environmental concerns, may arise. Regulatory approvals and public acceptance can be time-consuming. |
| Trait Stacking | Streamlined development of resilient crop varieties. | Enables the incorporation of multiple desirable traits into a single crop variety, enhancing resilience. | Consistency of crop performance across different environments may be challenging to achieve. Managing the interactions between stacked traits requires careful consideration. |
| Enhancing research across disciplines | Integrating SB to accelerate research in diverse fields. | Expediting experiments, data collection, and hypothesis testing, thus aiding in a deeper understanding of plant responses to environmental stressors, refining cultivation practices, and exploring plant physiology. It enables informed decision-making and innovative problem-solving across diverse research areas. | The need for controlled environments, often associated with speed breeding, can be a financial burden and a logistical challenge, potentially limiting its widespread use in diverse research disciplines. Furthermore, the suitability of this method varies among crop species, and effectively managing rapid growth cycles and plant phenology may require specialized expertise, which can pose barriers in some research fields. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potts, J.; Jangra, S.; Michael, V.N.; Wu, X. Speed Breeding for Crop Improvement and Food Security. Crops 2023, 3, 276-291. https://doi.org/10.3390/crops3040025
Potts J, Jangra S, Michael VN, Wu X. Speed Breeding for Crop Improvement and Food Security. Crops. 2023; 3(4):276-291. https://doi.org/10.3390/crops3040025
Chicago/Turabian StylePotts, Jesse, Sumit Jangra, Vincent N. Michael, and Xingbo Wu. 2023. "Speed Breeding for Crop Improvement and Food Security" Crops 3, no. 4: 276-291. https://doi.org/10.3390/crops3040025
APA StylePotts, J., Jangra, S., Michael, V. N., & Wu, X. (2023). Speed Breeding for Crop Improvement and Food Security. Crops, 3(4), 276-291. https://doi.org/10.3390/crops3040025