

Effects of Cashew Nut Shell Extract on Ruminal Fermentation and Nutrient Digestibility under Continuous Culture


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatments
2.2. Continuous Culture Conditions
2.3. Sample Collection and Analysis
2.4. Statistical Analysis
3. Results and Discussion
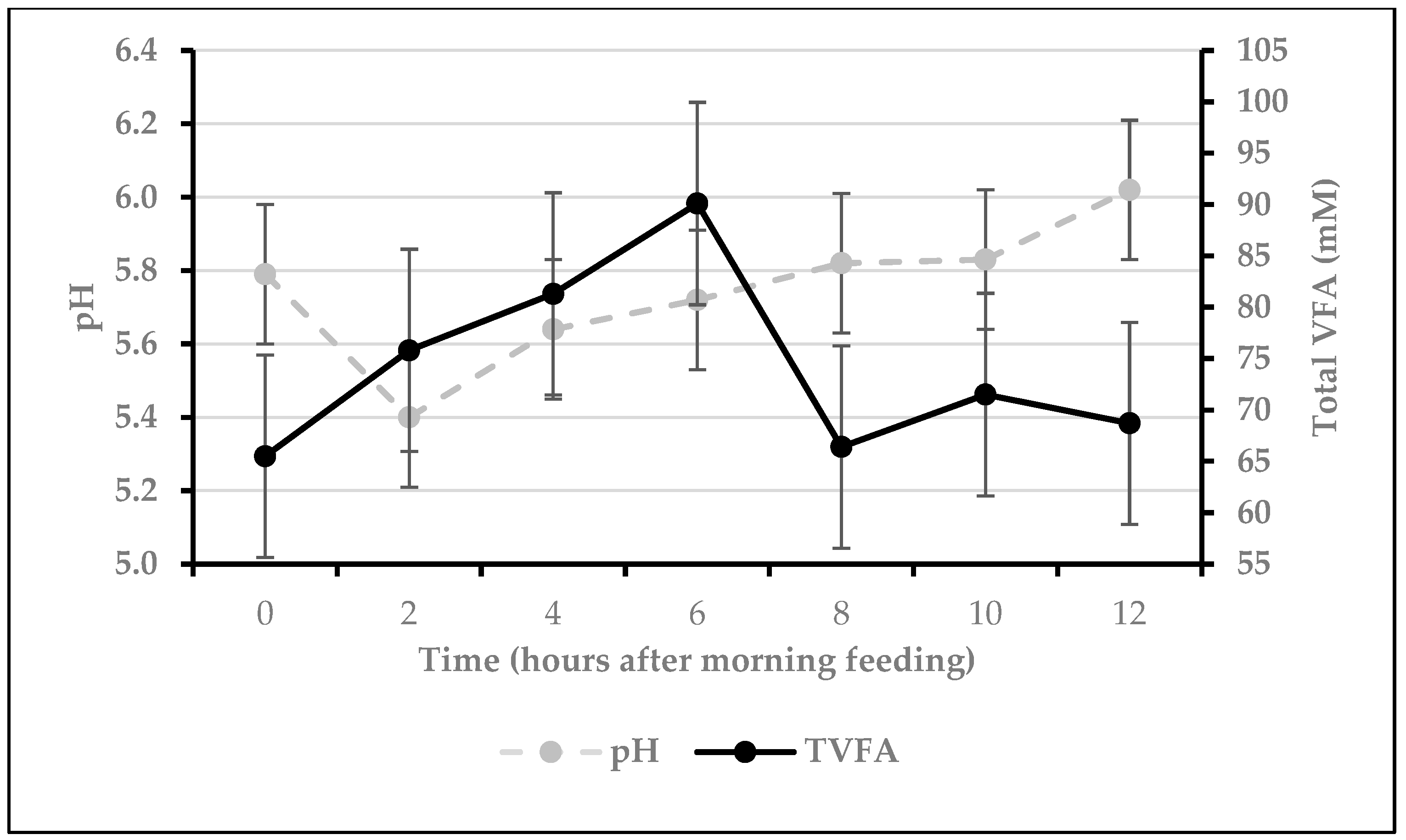
3.1. Culture Fermentation Measurements
3.2. Apparent Digestibility of Nutrients
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergen, W.G.; Bates, D.B. Ionophores: Their effect on production efficiency and mode of action. J. Anim. Sci. 1984, 58, 1465–1483. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Strobel, H.J. Effect of ionophores on ruminal fermentation. Appl. Environ. Microbiol. 1989, 55, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greathead, H. Plants and plant extracts for improving animal productivity. Proc. Nutr. Soc. 2003, 62, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calsamiglia, S.; Busquet, M.; Cardozo, P.W.; Castillejos, L.; Ferret, A. Invited review: Essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 2007, 90, 2580–2595. [Google Scholar] [CrossRef] [Green Version]
- Menon, A.R.R.; Pillai, C.K.S.; Sudha, J.D.; Mathew, A.G. Cashew nut shell liquid-its polymeric and other industrial products. J. Sci. Ind. Res. 1985, 44, 324–338. [Google Scholar]
- Watanabe, Y.; Suzuki, R.; Koike, S.; Nagashima, K.; Mochizuki, M.; Forster, R.J.; Kobayashi, Y. In vitro evaluation of cashew nut shell liquid as a methane-inhibiting and propionate-enhancing agent for ruminants. J. Dairy Sci. 2010, 93, 5258–5267. [Google Scholar] [CrossRef] [Green Version]
- Kubo, I.; Muroi, H.; Himejima, M.; Yamagiwa, Y.; Mera, H.; Tokushima, K.; Ohta, S.; Kamikawa, T. Structure-antibacterial activity relationships of anacardic acids. J. Agric. Food Chem. 1993, 41, 1016–1019. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Forano, E.; Martin, C.; Newbold, C.J. Microbial ecosystem and methanogenesis in ruminants. Animal 2010, 4, 1024–1036. [Google Scholar] [CrossRef] [Green Version]
- Shinkai, T.; Enishi, O.; Mitsumori, M.; Higuchi, K.; Kobayashi, Y.; Takenaka, A.; Nagashima, K.; Mochizuki, M. Mitigation of methane production from cattle by feeding cashew nut shell liquid. J. Dairy Sci. 2012, 95, 5308–5316. [Google Scholar] [CrossRef] [Green Version]
- Branco, A.F.; Giallongo, F.; Frederick, T.; Weeks, H.; Oh, J.; Hristov, A.N. Effect of technical cashew nut shell liquid on rumen methane emission and lactation performance of dairy cows. J. Dairy Sci. 2015, 98, 4030–4040. [Google Scholar] [CrossRef] [Green Version]
- Compton, C.; Peña, O.M.; Velasquez, C.; Lascano, G.J.; Mechor, G.D.; Jenkins, T.C.; Aguerre, M.J. A comparison of ionophore sources showed differences in volatile fatty acid changes but equal effects on digestibility in continuous cultures of ruminal microorganisms. J. Dairy Sci. 2022, 105 (Suppl. S1), 71. [Google Scholar]
- Fuentes, M.C.; Calsamiglia, S.; Cardozo, P.W.; Vlaeminck, B. Effect of pH and level of concentrate in the diet on the production of biohydrogenation intermediates in a dual-flow continuous culture. J. Dairy Sci. 2009, 92, 4456–4466. [Google Scholar] [CrossRef] [Green Version]
- Slyter, L.L.; Bryant, M.P.; Wolin, M.J. Effect of pH on population and fermentation in a continuously cultured rumen ecosystem. Appl. Microbiol. 1966, 14, 573–578. [Google Scholar] [CrossRef]
- Teather, R.M.; Sauer, F.D. A naturally compartmented rumen simulation system for the continuous culture of rumen bacteria and protozoa. J. Dairy Sci. 1988, 71, 666–673. [Google Scholar] [CrossRef]
- Jenkins, T.C.; Bridges, W.C., Jr.; Harrison, J.H.; Young, K.M. Addition of potassium carbonate to continuous cultures of mixed ruminal bacteria shifts volatile fatty acids and daily production of biohydrogenation intermediates. J. Dairy Sci. 2014, 97, 975–984. [Google Scholar] [CrossRef] [Green Version]
- Lascano, G.J.; Alende, M.; Koch, L.E.; Jenkins, T.C. Changes in fermentation and biohydrogenation intermediates in continuous cultures fed low and high levels of fat with increasing rates of starch degradability. J. Dairy Sci. 2016, 99, 6334–6341. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 18th ed.; AOAC International: Rockville, MD, USA, 2006. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 17th ed.; AOAC International: Rockville, MD, USA, 2000. [Google Scholar]
- Hall, M.B. Determination of starch, including maltooligosaccharides, in animal feeds: Comparison of methods and a method recommended for AOAC collaborative study. J. AOAC Int. 2009, 92, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Suzuki, Y.; Hayashi, S.; Suzuki, Y.; Koike, S.; Kobayashi, Y. Potency of cashew nut shell liquid in rumen modulation under different dietary conditions and indication of its surfactant action against rumen bacteria. J. Anim. Sci. Technol. 2017, 59, 27. [Google Scholar] [CrossRef] [Green Version]
- Goetz, B.M.; Horst, E.A.; Mayorga, E.J.; Abeyta, M.A.; Rodriguez-Jimenez, S.; Carta, S.; Hikita, C.; Watanabe, T.; Baumgard, L.H. Effects of a cashew nut shell extract on energetic metabolism and inflammatory biomarkers in transition dairy cows. J. Dairy Sci. 2020, 103 (Suppl. S1), T147. [Google Scholar]
- Sarmikasoglou, E.; Johnson, M.; Vinyard, J.; Sumadong, P.; Lobo, R.R.; Cordero, J.A.; Bahman, A.; Ravelo, A.; Halima, S.; Hikita, C.; et al. Effects of cashew nut shell extract and monensin on microbial fermentation in a dual-flow continuous culture. J. Dairy Sci. 2022, 105 (Suppl. S1), 274. [Google Scholar]
- Vallimont, J.E.; Bargo, F.; Cassidy, T.W.; Luchini, N.D.; Broderick, G.A.; Varga, G.A. Effects of replacing dietary starch with sucrose on ruminal fermentation and nitrogen metabolism in continuous culture. J. Dairy Sci. 2004, 87, 4221–4229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, D.; Karnati, S.K.R.; Wagner, B.; Firkins, J.L.; Eastridge, M.L.; Aldrich, J.M. Essential oil and monensin affect ruminal fermentation and the protozoal population in continuous culture. J. Dairy Sci. 2018, 101, 5069–5508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arce-Cordero, J.A.; Monteiro, H.F.; Lelis, A.L.; Lima, L.R.; Restelatto, R.; Brandao, V.L.N.; Leclerc, H.; Vyas, D.; Faciola, A.P. Copper sulfate and sodium selenite lipid-microencapsulation modifies ruminal microbial fermentation in a dual-flow continuous culture system. J. Dairy Sci. 2020, 103, 7068–7080. [Google Scholar] [CrossRef]
- Goetz, B.M.; Horst, E.A.; Mayorga, E.J.; Abeyta, M.A.; Rodriguez-Jimenez, S.; Hikita, C.; Watanabe, T.; Lourenco, J.M.; Callaway, T.R.; Baumgard, L.H. Effects of a cashew nut shell extract on nutrient digestibility and rumen microbiome in transition dairy cows. J. Dairy Sci. 2021, 104 (Suppl. S1), P443. [Google Scholar]


{kind=link}
| Item | Control Diet (CO) |
|---|---|
| Ingredient, % of DM | |
| Corn silage 1 | 37.0 |
| Alfalfa hay 2 | 15.0 |
| Ground corn | 23.7 |
| Solvent soybean meal | 9.5 |
| Expeller soybean meal 3 | 4.0 |
| Soyhulls | 8.0 |
| Energy Booster 100 | 1.0 |
| Mineral premix 4 | 1.8 |
| Item | CNSE (ppm) | |||
|---|---|---|---|---|
| 0 | 100 | 200 | 300 | |
| DM, % | 92.1 | 92.4 | 92.3 | 91.6 |
| OM, % DM | 94.2 | 93.6 | 94.1 | 93.8 |
| CP, % DM | 16.7 | 17.2 | 17.2 | 16.9 |
| NDF, % DM | 29.2 | 30.0 | 28.9 | 28.9 |
| ADF, % DM | 19.1 | 18.5 | 19.7 | 18.5 |
| Starch, % DM | 29.9 | 29.7 | 29.1 | 29.4 |
| Item | CNSE (ppm) | SEM 2 | p-Value 1 | ||||
|---|---|---|---|---|---|---|---|
| 0 | 100 | 200 | 300 | L | Q | ||
| pH | 5.78 | 5.72 | 5.75 | 5.80 | 0.20 | 0.51 | 0.47 |
| Total VFA, mM | 76.8 | 70.5 | 72.3 | 71.2 | 10.9 | 0.46 | 0.51 |
| VFA, mol/100 mol | |||||||
| Acetate | 37.5 | 39.2 | 35.8 | 38.1 | 1.57 | 0.96 | 0.51 |
| Propionate | 27.8 | 26.6 | 28.6 | 28.5 | 8.12 | 0.67 | 0.92 |
| Butyrate | 17.8 | 17.6 | 18.4 | 16.7 | 1.81 | 0.61 | 0.57 |
| Isovalerate | 0.82 | 1.12 | 1.09 | 0.82 | 0.44 | 0.82 | 0.26 |
| Valerate | 8.70 | 8.71 | 9.30 | 8.52 | 1.56 | 0.93 | 0.58 |
| A:P 3 | 1.24 | 1.29 | 1.17 | 1.21 | 1.29 | 0.64 | 0.78 |
| Item | CNSE (ppm) | SEM 2 | p-Value 1 | ||||
|---|---|---|---|---|---|---|---|
| 0 | 100 | 200 | 300 | L | Q | ||
| DM, % | 54.2 | 59.2 | 53.4 | 56.2 | 4.27 | 0.57 | 0.57 |
| OM, % | 63.3 | 66.2 | 62.6 | 65.5 | 3.33 | 0.74 | 0.85 |
| NDF, % | 57.8 | 62.1 | 55.7 | 59.9 | 4.72 | 0.71 | 0.71 |
| Starch, % | 97.8 | 97.5 | 96.8 | 97.4 | 0.65 | 0.47 | 0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Compton, C.; Peña, O.M.; Hikita, C.; Watanabe, T.; Jenkins, T.C.; Lascano, G.J.; Aguerre, M.J. Effects of Cashew Nut Shell Extract on Ruminal Fermentation and Nutrient Digestibility under Continuous Culture. Ruminants 2023, 3, 92-99. https://doi.org/10.3390/ruminants3010008
Compton C, Peña OM, Hikita C, Watanabe T, Jenkins TC, Lascano GJ, Aguerre MJ. Effects of Cashew Nut Shell Extract on Ruminal Fermentation and Nutrient Digestibility under Continuous Culture. Ruminants. 2023; 3(1):92-99. https://doi.org/10.3390/ruminants3010008
Chicago/Turabian StyleCompton, Chandler, Omar M. Peña, Chie Hikita, Tomonori Watanabe, Thomas C. Jenkins, Gustavo J. Lascano, and Matias J. Aguerre. 2023. "Effects of Cashew Nut Shell Extract on Ruminal Fermentation and Nutrient Digestibility under Continuous Culture" Ruminants 3, no. 1: 92-99. https://doi.org/10.3390/ruminants3010008
APA StyleCompton, C., Peña, O. M., Hikita, C., Watanabe, T., Jenkins, T. C., Lascano, G. J., & Aguerre, M. J. (2023). Effects of Cashew Nut Shell Extract on Ruminal Fermentation and Nutrient Digestibility under Continuous Culture. Ruminants, 3(1), 92-99. https://doi.org/10.3390/ruminants3010008