

The Lipidome of the Gastrointestinal Tract in Lactating Holstein Cows


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Handling and Experimental Design
2.2. Sample Collection
2.3. Lipid Extraction and Lipidomics
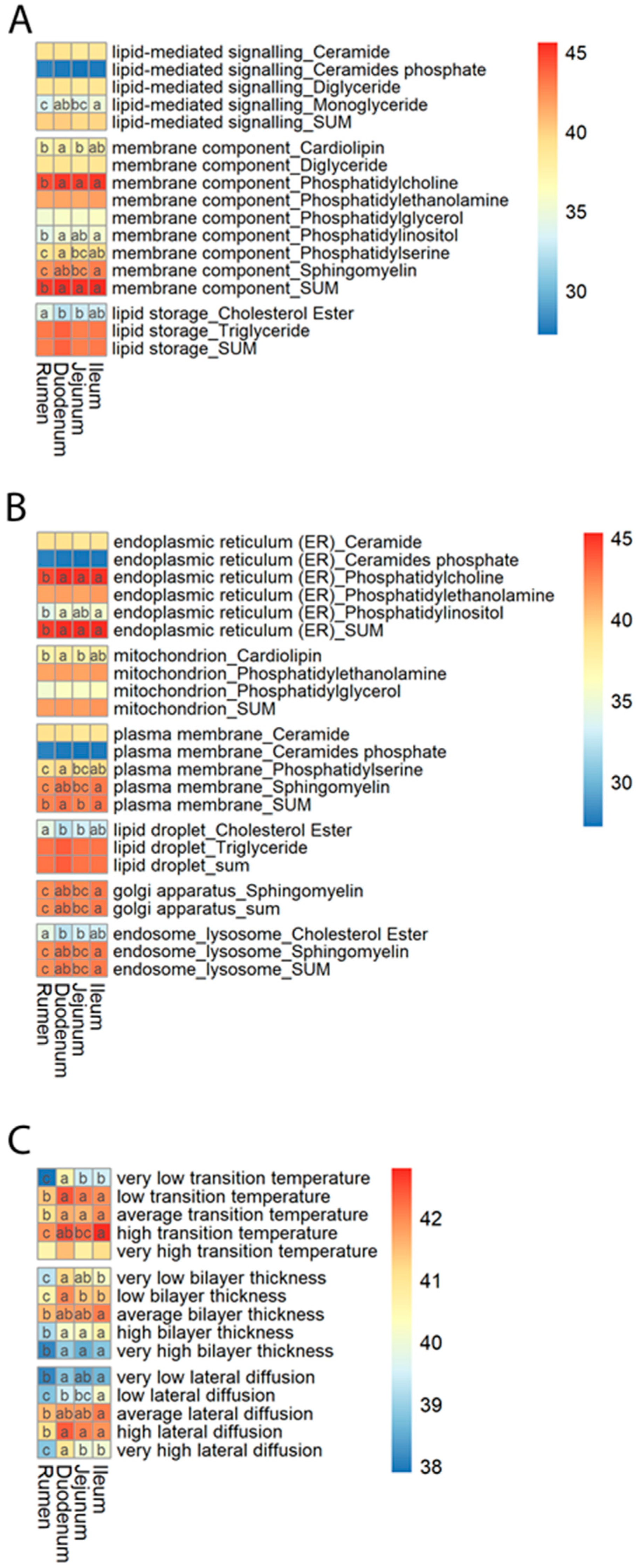
2.4. Functional Mapping
2.5. Statistical Analysis
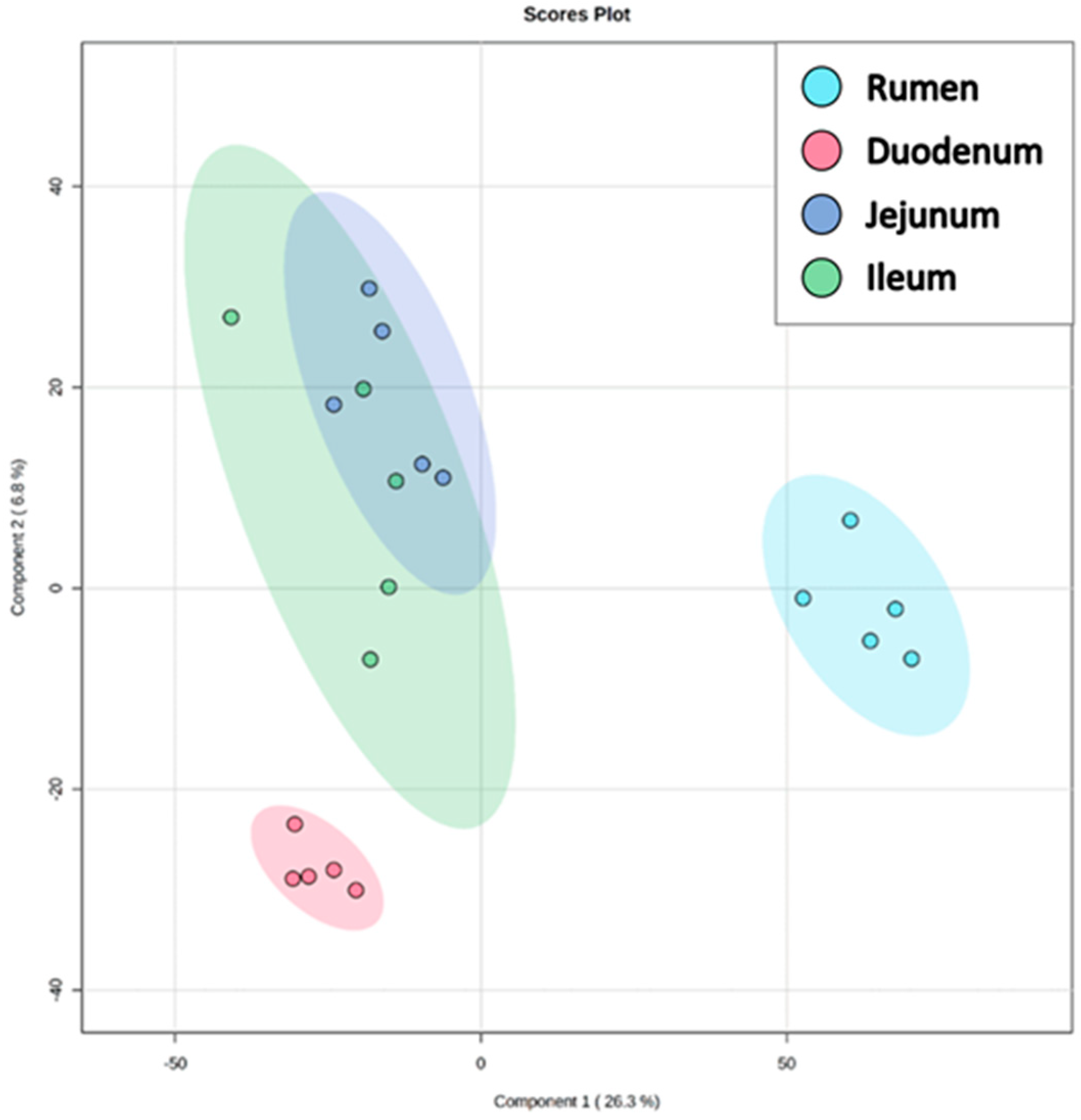
3. Results and Discussion
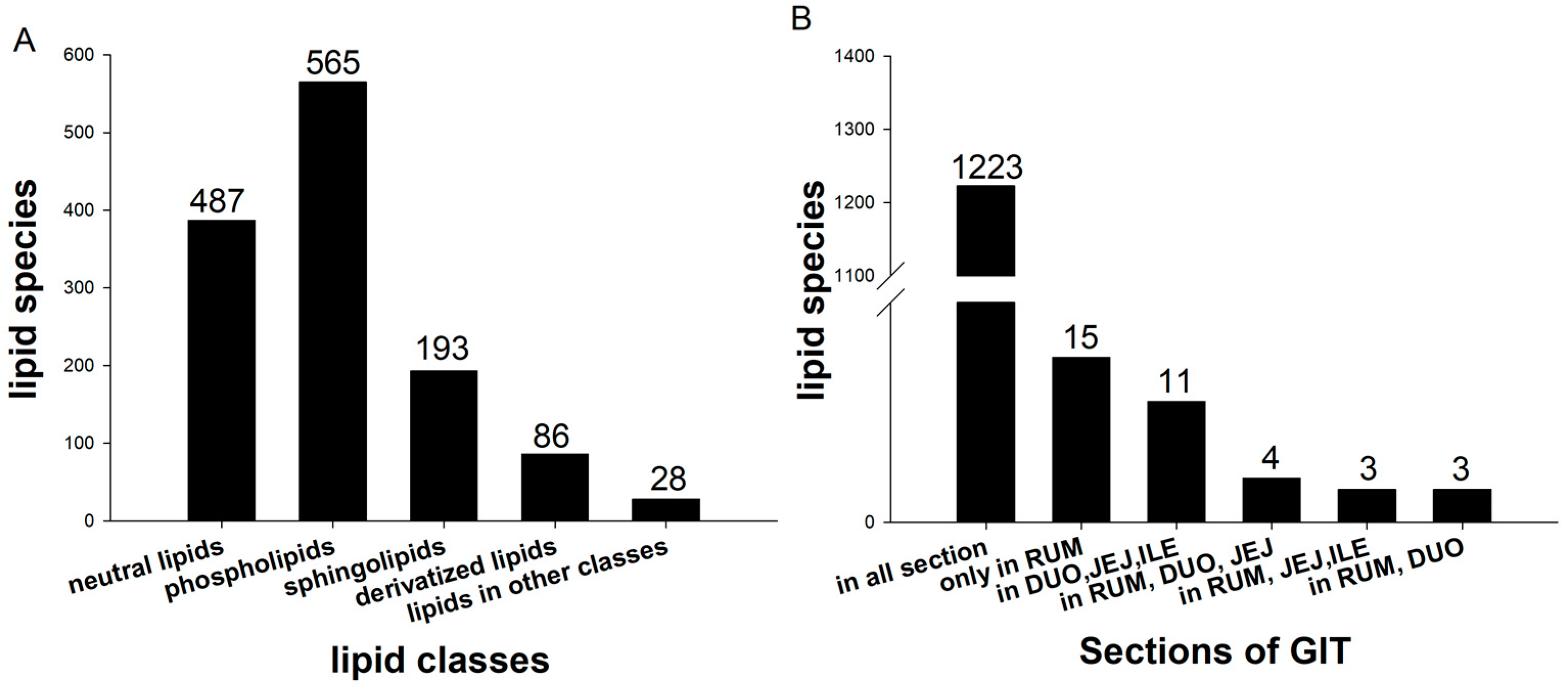
3.1. Overall Lipid Classification and Distribution
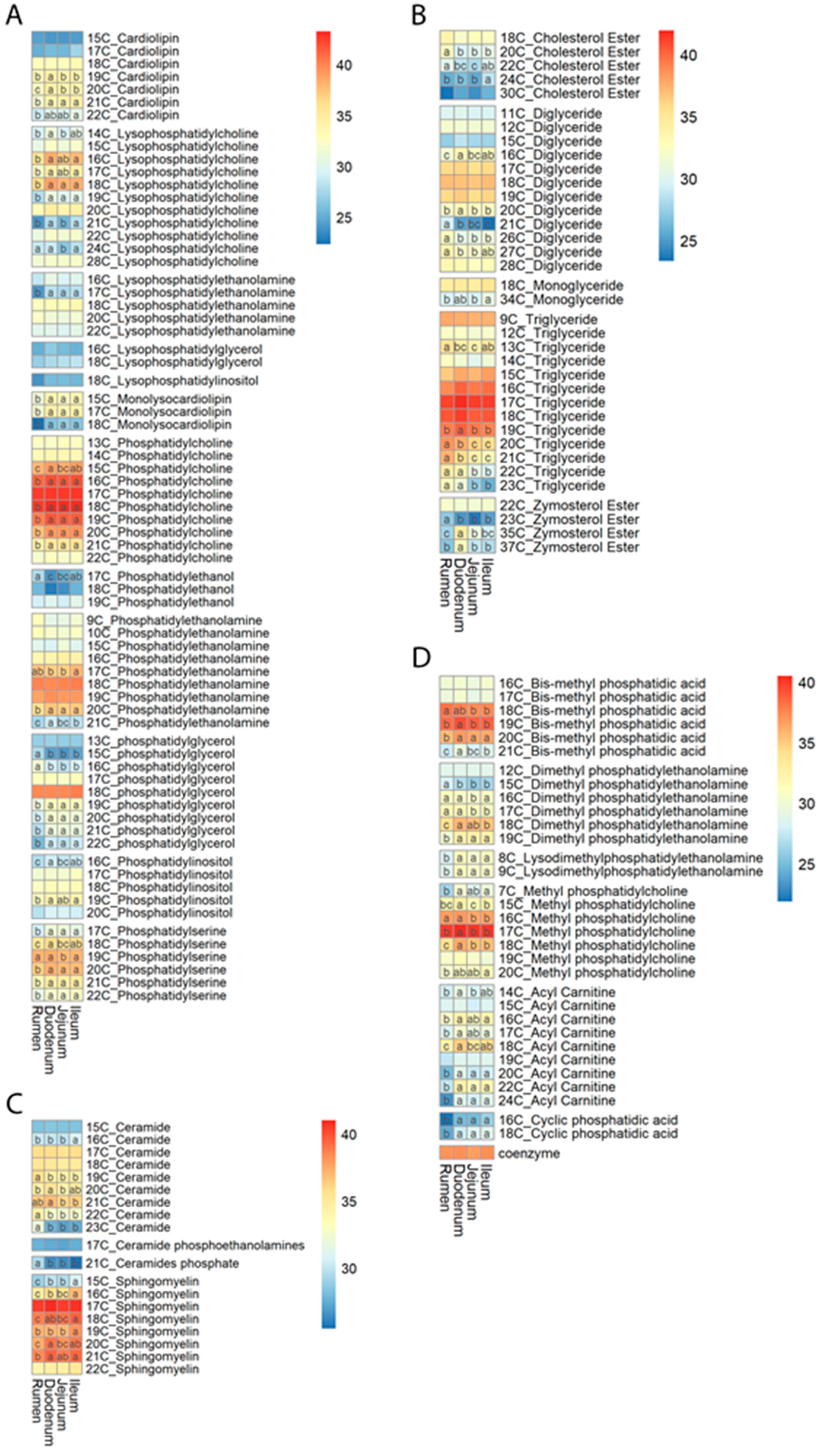
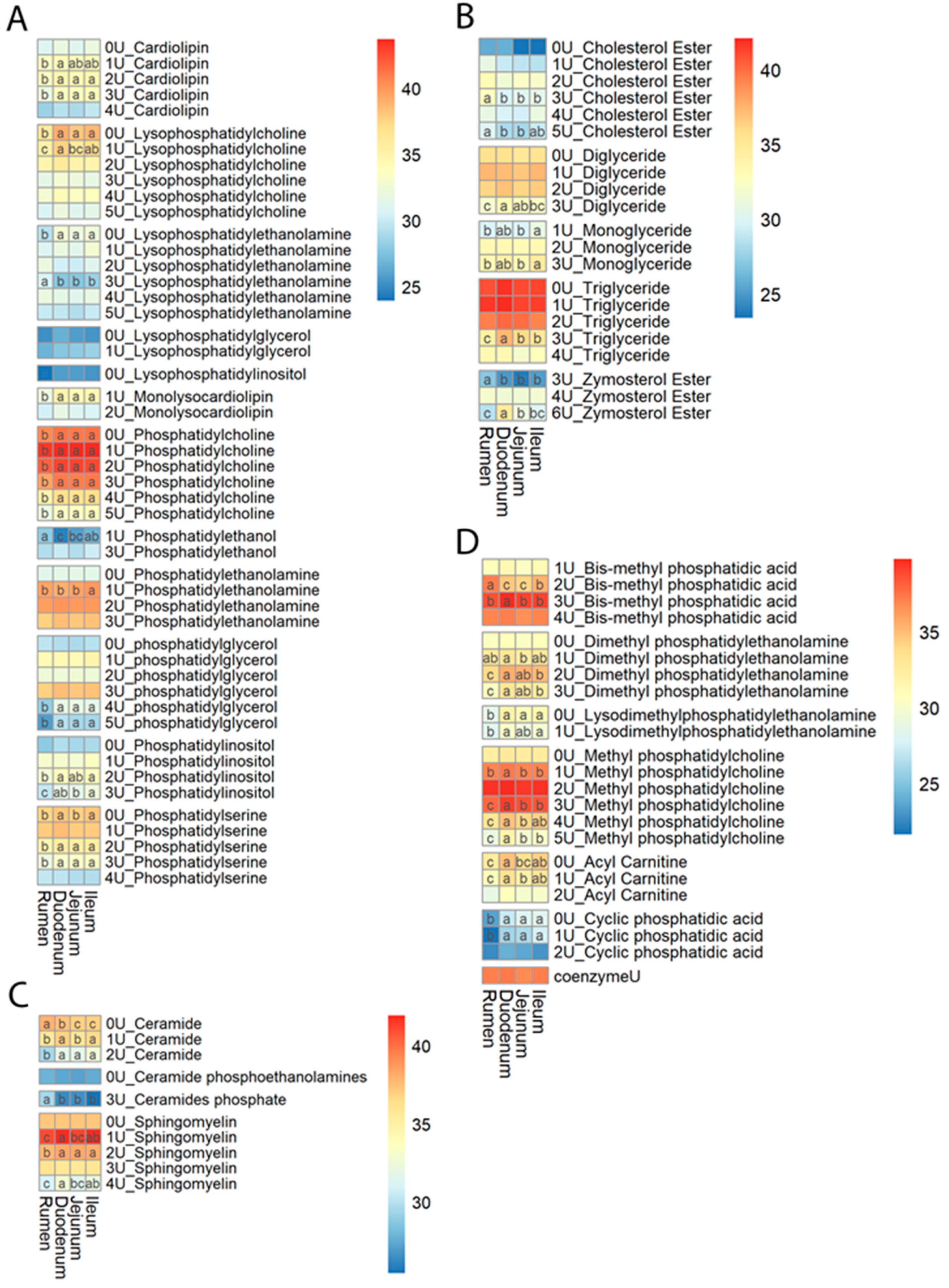
3.2. Lipid Content Profiles
3.3. Chain Length and Unsaturation of Fatty Acids
3.4. Lipid Ontology
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bloom, M.; Evans, E. Physical properties of the fluid lipid-bilayer component of cell membranes: A perspective. Q. Rev. Biophys. 1991, 24, 293–397. [Google Scholar] [CrossRef]
- English, A.R.; Zurek, N.; Voeltz, G.K. Peripheral ER structure and function. Curr. Opin. Cell Biol. 2009, 21, 596–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, T. Cell Biology, 4th ed.; Elyse O’Grady: Philadelphia, The Netherlands, 2023; pp. 237–250. [Google Scholar]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Kullenberg, D.; Taylor, L.A.; Schneider, M.; Massing, U. Health effects of dietary phospholipids. Lipids Health Dis. 2012, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Ghanem, M.; Lewis, G.F.; Xiao, C.T. Recent advances in cytoplasmic lipid droplet metabolism in intestinal enterocyte. BBA-Mol. Cell Biol. Lipids 2022, 1867, 159197. [Google Scholar] [CrossRef]
- Olzmann, J.A.; Carvalho, P. Dynamics and functions of lipid droplets. Nat. Rev. Mol. Cell Biol. 2019, 20, 137–155. [Google Scholar] [CrossRef]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’H, J.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Raajendiran, A.; Tsiloulis, T.; Watt, M.J. Adipose tissue development and the molecular regulation of lipid metabolism. Essays Biochem. 2016, 60, 437–450. [Google Scholar]
- Merrill, A.H., Jr.; Sullards, M.C.; Wang, E.; Voss, K.A.; Riley, R.T. Sphingolipid metabolism: Roles in signal transduction and disruption by fumonisins. Environ. Health Perspect. 2001, 109 (Suppl. 2), 283–289. [Google Scholar] [PubMed]
- Hla, T.; Dannenberg, A.J. Sphingolipid signaling in metabolic disorders. Cell Metab. 2012, 16, 420–434. [Google Scholar] [CrossRef] [Green Version]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. BBA-Mol. Basis Dis. 2012, 1822, 1442–1452. [Google Scholar] [CrossRef] [Green Version]
- Kvidera, S.K.; Horst, E.A.; Sanz Fernandez, M.V.; Abuajamieh, M.; Ganesan, S.; Gorden, P.J.; Green, H.B.; Schoenberg, K.M.; Trout, W.E.; Keating, A.F.; et al. Characterizing effects of feed restriction and glucagon-like peptide 2 administration on biomarkers of inflammation and intestinal morphology. J. Dairy Sci. 2017, 100, 9402–9417. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.Y.; Gu, H.; Jiang, L.H.; Xu, W.; Liu, C.Q.; Li, Y.; Qian, X.Y.; Li, D.D.; Li, Z.L.; Hu, J.; et al. Cocaine modifies brain lipidome in mice. Mol. Cell Neurosci. 2017, 85, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.Y.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L.; et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, M.R.; Jeucken, A.; Wassenaar, T.A.; van de Lest, C.H.A.; Brouwers, J.F.; Helms, J.B. LION/web: A web-based ontology enrichment tool for lipidomic data analysis. Gigascience 2019, 8, giz061. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.Q.; Zhou, G.Y.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J.G. Using MetaboAnalyst 5.0 for LC-HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef]
- Klopp, R.N.; Ferreira, C.R.; Casey, T.M.; Boerman, J.P. Relationship of cow and calf circulating lipidomes with colostrum lipid composition and metabolic status of the cow. J. Dairy Sci. 2022, 105, 1768–1787. [Google Scholar] [CrossRef]
- Rico, J.E.; Saed Samii, S.; Zang, Y.; Deme, P.; Haughey, N.J.; Grilli, E.; McFadden, J.W. Characterization of the Plasma Lipidome in Dairy Cattle Transitioning from Gestation to Lactation: Identifying Novel Biomarkers of Metabolic Impairment. Metabolites 2021, 11, 290. [Google Scholar] [CrossRef] [PubMed]
- Brasitus, T.A.; Dahiya, R.; Dudeja, P.K. Rat proximal small intestinal Golgi membranes: Lipid composition and fluidity. Biochim. et Biophys. Acta (BBA)-Lipids Lipid Metab. 1988, 958, 218–226. [Google Scholar] [CrossRef]
- Omodeo-Sale, F.; Lindi, C.; Marciani, P.; Cavatorta, P.; Sartor, G.; Masotti, L.; Esposito, G. Postnatal maturation of rat intestinal membrane: Lipid composition and fluidity. Comp. Biochem. Physiol. Part A Physiol. 1991, 100, 301–307. [Google Scholar] [CrossRef]
- Keelan, M.; Walker, K.; Thomson, A.B.R. Intestinal brush border membrane marker enzymes, lipid composition and villus morphology: Effect of fasting and diabetes mellitus in rats. Comp. Biochem. Physiol. Part A Physiol. 1985, 82, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Brasitus, T.A.; Dudeja, P.K.; Dahiya, R.; Halline, A. Dexamethasone-induced alterations in lipid composition and fluidity of rat proximal-small-intestinal brush-border membranes. Biochem. J. 1987, 248, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.M.; Hostetler, B.; Ling, S.; Mone, M.; Watkins, J.B. Intestinal membrane lipid composition and fluidity during development in the rat. Am. J. Physiol. 1985, 248, G200–G207. [Google Scholar] [CrossRef] [PubMed]
- Forstner, G.G.; Tanaka, K.; Isselbacher, K.J. Lipid composition of the isolated rat intestinal microvillus membrane. Biochem. J. 1968, 109, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Kaur, J. Chronic Ethanol Feeding Affects Intestinal Mucus Lipid Composition and Glycosylation in Rats. Ann. Nutr. Metab. 2002, 46, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Duranthon, V.; Frémont, L.; Léger, C.L. Effect of essential fatty acid deficiency on lipid composition of basolateral plasma membrane of pig intestinal mucosal cells. Lipids 1991, 26, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Daveloose, D.; Linard, A.; Arfi, T.; Viret, J.; Christon, R. Simultaneous changes in lipid composition, fluidity and enzyme activity in piglet intestinal brush border membrane as affected by dietary polyunsaturated fatty acid deficiency. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1993, 1166, 229–237. [Google Scholar] [CrossRef]
- Lopez-Pedrosa, J.M.; Torres, M.I.; Fernández, M.I.; Ríos, A.; Gil, A. Severe Malnutrition Alters Lipid Composition and Fatty Acid Profile of Small Intestine in Newborn Piglets. J. Nutr. 1998, 128, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.M.; Bostwick, H.E.; Danziger, M.D.; Newman, L.J.; Medow, M.S. Ontogeny of basolateral membrane lipid composition and fluidity in small intestine. Am. J. Physiol. 1989, 257, G138–G144. [Google Scholar] [CrossRef]
- Hauser, H.; Howell, K.; Dawson, R.M.C.; Bowyer, D.E. Rabbit small intestinal brush border membrane. Prep. Lipid Compos. Biochim. Biophys. Acta (BBA)-Biomembr. 1980, 602, 567–577. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Role of Cardiolipin in Mitochondrial Function and Dynamics in Health and Disease: Molecular and Pharmacological Aspects. Cells 2019, 8, 728. [Google Scholar] [CrossRef] [Green Version]
- Sparagna, G.C.; Lesnefsky, E.J. Cardiolipin remodeling in the heart. J. Cardiovasc. Pharmacol. 2009, 53, 290–301. [Google Scholar] [CrossRef]
- Ren, M.; Phoon, C.K.; Schlame, M. Metabolism and function of mitochondrial cardiolipin. Prog. Lipid Res. 2014, 55, 1–16. [Google Scholar] [CrossRef]
- Claypool, S.M. Cardiolipin, a critical determinant of mitochondrial carrier protein assembly and function. Biochim. Biophys. Acta 2009, 1788, 2059–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBride, B.W.; Kelly, J.M. Energy cost of absorption and metabolism in the ruminant gastrointestinal tract and liver: A review. J. Anim. Sci. 1990, 68, 2997–3010. [Google Scholar] [CrossRef]
- Ceciliani, F.; Audano, M.; Addis, M.F.; Lecchi, C.; Ghaffari, M.H.; Albertini, M.; Tangorra, F.; Piccinini, R.; Caruso, D.; Mitro, N.; et al. The untargeted lipidomic profile of quarter milk from dairy cows with subclinical intramammary infection by non-aureus staphylococci. J. Dairy Sci. 2021, 104, 10268–10281. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, S.; Zhao, H.; Li, L.; Li, Y.; Tu, Y.; Jiang, L.; Zhao, G. Lipidomic profiling using GC and LC-MS/MS revealed the improved milk quality and lipid composition in dairy cows supplemented with citrus peel extract. Food Res. Int. 2022, 161, 111767. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Wang, L.; Du, H.; Yang, G.; Fu, T.; Lian, H.; Sun, Y.; Liu, S.; Zhang, L.; Gao, T. Metabolomic and Lipidomic Approaches to Evaluate the Effects of Eucommia ulmoides Leaves on Milk Quality and Biochemical Properties. Front. Vet. Sci. 2021, 8, 644967. [Google Scholar] [CrossRef]
- Law, S.H.; Chan, M.L.; Marathe, G.K.; Parveen, F.; Chen, C.H.; Ke, L.Y. An Updated Review of Lysophosphatidylcholine Metabolism in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineiro, R.; Falasca, M. Lysophosphatidylinositol signalling: New wine from an old bottle. Biochim. Biophys. Acta 2012, 1821, 694–705. [Google Scholar] [CrossRef] [Green Version]
- Makide, K.; Kitamura, H.; Sato, Y.; Okutani, M.; Aoki, J. Emerging lysophospholipid mediators, lysophosphatidylserine, lysophosphatidylthreonine, lysophosphatidylethanolamine and lysophosphatidylglycerol. Prostaglandins Other Lipid Mediat. 2009, 89, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, I.; Imaizumi, K.; Sugano, M. Absorption and transport of base moieties of phosphatidylcholine and phosphatidylethanolamine in rats. Biochim. Biophys. Acta 1987, 921, 245–253. [Google Scholar] [PubMed]
- Goodman, D.S. Cholesterol ester metabolism. Physiol. Rev. 1965, 45, 747–839. [Google Scholar] [CrossRef]
- Chang, T.Y.; Li, B.L.; Chang, C.C.; Urano, Y. Acyl-coenzyme A:cholesterol acyltransferases. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E1–E9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, M.A.; Vandervoort, G.; AlZahal, O.; Hook, S.E.; Matthews, J.C.; McBride, B.W. Rumen epithelial adaptation to high-grain diets involves the coordinated regulation of genes involved in cholesterol homeostasis. Physiol. Genom. 2011, 43, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Aschenbach, J.R.; Penner, G.B.; Stumpff, F.; Gabel, G. Ruminant Nutrition Symposium: Role of fermentation acid absorption in the regulation of ruminal pH. J. Anim. Sci. 2011, 89, 1092–1107. [Google Scholar] [CrossRef] [Green Version]
- Omoigui, S. Cholesterol synthesis is the trigger and isoprenoid dependent interleukin-6 mediated inflammation is the common causative factor and therapeutic target for atherosclerotic vascular disease and age-related disorders including osteoporosis and type 2 diabetes. Med. Hypotheses 2005, 65, 559–569. [Google Scholar]
- Goodlad, R.A. Some effects of diet on the mitotic index and the cell cycle of the ruminal epithelium of sheep. Exp. Physiol. 1981, 66, 487–499. [Google Scholar] [CrossRef] [Green Version]
- Japelt, R.B.; Jakobsen, J. Vitamin D in plants: A review of occurrence, analysis, and biosynthesis. Front. Plant Sci. 2013, 4, 136. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, Y.; Gaucher, E.A. Evolution of bacterial steroid biosynthesis and its impact on eukaryogenesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2101276118. [Google Scholar] [CrossRef]
- Li, Y.; Nicholson, R.J.; Summers, S.A. Ceramide signaling in the gut. Mol. Cell Endocrinol. 2022, 544, 111554. [Google Scholar] [CrossRef]
- Uchida, Y. Ceramide signaling in mammalian epidermis. Biochim. Biophys. Acta 2014, 1841, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Morad, S.A.; Cabot, M.C. Ceramide-orchestrated signalling in cancer cells. Nat. Rev. Cancer 2013, 13, 51–65. [Google Scholar] [CrossRef]
- Becker, K.A.; Tummler, B.; Gulbins, E.; Grassme, H. Accumulation of ceramide in the trachea and intestine of cystic fibrosis mice causes inflammation and cell death. Biochem. Biophys. Res. Commun. 2010, 403, 368–374. [Google Scholar] [CrossRef]
- Qu, Q.; Zeng, F.; Liu, X.; Wang, Q.J.; Deng, F. Fatty acid oxidation and carnitine palmitoyltransferase I: Emerging therapeutic targets in cancer. Cell Death Dis. 2016, 7, e2226. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, X.; Xiong, X.; Yin, Y. Energy metabolism in intestinal epithelial cells during maturation along the crypt-villus axis. Sci. Rep. 2016, 6, 31917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvi, P.S.; Cowles, R.A. Butyrate and the Intestinal Epithelium: Modulation of Proliferation and Inflammation in Homeostasis and Disease. Cells 2021, 10, 1775. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.C. Lipid Metabolism in the Rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef] [PubMed]
- Vlaeminck, B.; Fievez, V.; Demeyer, D.; Dewhurst, R.J. Effect of forage:concentrate ratio on fatty acid composition of rumen bacteria isolated from ruminal and duodenal digesta. J. Dairy Sci. 2006, 89, 2668–2678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bainbridge, M.L.; Cersosimo, L.M.; Wright, A.D.G.; Kraft, J. Rumen bacterial communities shift across a lactation in Holstein, Jersey and Holstein × Jersey dairy cows and correlate to rumen function, bacterial fatty acid composition and production parameters. FEMS Microbiol. Ecol. 2016, 92, 59. [Google Scholar] [CrossRef] [Green Version]
- Contreras, G.A.; O’Boyle, N.J.; Herdt, T.H.; Sordillo, L.M. Lipomobilization in periparturient dairy cows influences the composition of plasma nonesterified fatty acids and leukocyte phospholipid fatty acids. J. Dairy Sci. 2010, 93, 2508–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.G. Lipid phase transitions and phase diagrams I. Lipid phase transitions. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1977, 472, 237–281. [Google Scholar] [CrossRef]
- Roberts, G.C.K. European Biophysical Societies Association. In Encyclopedia of Biophysics, 1st ed.; Springer: Berlin, Germany, 2013; pp. 1841–1854. [Google Scholar]
- Oshima, A.; Nakashima, H.; Sumitomo, K. Evaluation of Lateral Diffusion of Lipids in Continuous Membranes between Freestanding and Supported Areas by Fluorescence Recovery after Photobleaching. Langmuir 2019, 35, 11725–11734. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class Group | Class Name | Rumen% | Duodenum% | Jejunum% | Ileum% | SEM | p-Value | DIJ * vs. Rumen |
|---|---|---|---|---|---|---|---|---|
| Phospholipids | Cardiolipin | 0.118 | 0.154 | 0.151 | 0.152 | 0.000150 | 0.20 | 0.04 |
| Monolysocardiolipin | 0.022 b | 0.089 a | 0.072 a | 0.078 a | 0.000194 | 0.01 | <0.01 | |
| Phosphatidylcholine | 52.4 | 54.0 | 60.2 | 58.2 | 0.024640 | 0.07 | 0.06 | |
| Phosphatidylethanolamine | 6.009 | 4.180 | 5.112 | 5.170 | 0.004848 | 0.08 | 0.04 | |
| Phosphatidylethanol | 0.003 | 0.002 | 0.002 | 0.003 | 0.000004 | 0.18 | 0.32 | |
| Phosphatidylserine | 1.591 | 1.502 | 1.388 | 1.410 | 0.001011 | 0.49 | 0.20 | |
| Phosphatidylglycerol | 0.102 | 0.088 | 0.104 | 0.112 | 0.000150 | 0.42 | 0.95 | |
| Phosphatidylinositol | 0.059 | 0.067 | 0.067 | 0.077 | 0.000129 | 0.77 | 0.42 | |
| Lysophosphatidylcholine | 0.562 b | 1.953 a | 1.578 a | 1.703 a | 0.004832 | 0.05 | 0.01 | |
| Lysophosphatidylethanolamine | 0.060 | 0.041 | 0.045 | 0.058 | 0.000159 | 0.55 | 0.34 | |
| Lysophosphatidylglycerol | 0.001 | 0.001 | 0.001 | 0.001 | 0.000003 | 0.37 | 0.11 | |
| Lysophosphatidylinositol | <0.001 | <0.001 | 0.001 | <0.001 | 0.000002 | 0.41 | 0.43 | |
| Neutral lipids | Monoglyceride | 0.074 | 0.065 | 0.058 | 0.093 | 0.000100 | 0.10 | 0.85 |
| Diglyceride | 1.026 a | 0.684 b | 0.645 b | 0.707 b | 0.000684 | <0.01 | <0.01 | |
| Triglyceride | 20.6 | 20.6 | 15.9 | 16.4 | 0.031940 | 0.59 | 0.43 | |
| Cholesterol Ester | 0.072 a | 0.012 c | 0.023 bc | 0.028 b | 0.000050 | <0.01 | <0.01 | |
| Zymosterol Ester | 0.016 b | 0.091 a | 0.021 b | 0.012 b | 0.000115 | <0.01 | 0.07 | |
| Sphingolipids | Ceramides | 1.037 a | 0.612 b | 0.500 b | 0.533 b | 0.000581 | <0.01 | <0.01 |
| Ceramides phosphate | 0.003 a | <0.001 b | <0.001 b | <0.001 b | 0.000003 | <0.01 | <0.01 | |
| Ceramide phosphoethanolamines | 0.001 | <0.001 | 0.001 | 0.001 | 0.000002 | 0.17 | 0.03 | |
| Sphingomyelin | 9.348 | 9.501 | 8.615 | 10.210 | 0.013140 | 0.86 | 0.95 | |
| Derivatized lipids (bioti nylation, diazomethane) | Bis-methyl phosphatidic acid | 2.457 | 2.169 | 1.924 | 1.614 | 0.002693 | 0.16 | 0.08 |
| Dimethyl phosphatidylethanolamine | 0.086 b | 0.147 a | 0.150 a | 0.113 b | 0.000113 | <0.01 | <0.01 | |
| Lysodimethylphosphatidylethanolamine | 0.003 b | 0.013 a | 0.010 a | 0.010 a | 0.000028 | 0.02 | <0.01 | |
| Methyl phosphatidylcholine | 3.807 a | 3.580 ab | 3.104 bc | 2.886 c | 0.001793 | 0.01 | 0.01 | |
| Fatty acyl and other lipids | Acyl Carnitine | 0.032 c | 0.094 a | 0.052 bc | 0.082 ab | 0.000143 | 0.02 | 0.01 |
| Coenzyme | 0.492 a | 0.344 b | 0.282 b | 0.349 b | 0.000338 | <0.01 | <0.01 | |
| Cyclic phosphatidic acid | 0.0001 | 0.0011 | 0.0016 | 0.0014 | 0.000005 | 0.07 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.; Loor, J.J. The Lipidome of the Gastrointestinal Tract in Lactating Holstein Cows. Ruminants 2023, 3, 76-91. https://doi.org/10.3390/ruminants3010007
Jiang Q, Loor JJ. The Lipidome of the Gastrointestinal Tract in Lactating Holstein Cows. Ruminants. 2023; 3(1):76-91. https://doi.org/10.3390/ruminants3010007
Chicago/Turabian StyleJiang, Qianming, and Juan J. Loor. 2023. "The Lipidome of the Gastrointestinal Tract in Lactating Holstein Cows" Ruminants 3, no. 1: 76-91. https://doi.org/10.3390/ruminants3010007
APA StyleJiang, Q., & Loor, J. J. (2023). The Lipidome of the Gastrointestinal Tract in Lactating Holstein Cows. Ruminants, 3(1), 76-91. https://doi.org/10.3390/ruminants3010007