

Presence and Persistence of Listeria monocytogenes in the Danish Ready-to-Eat Food Production Environment

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates and Other Relevant Information
2.2. Whole Genome Sequencing
2.3. Characterization of Isolates in Terms of Virulence, Resistance and Plasmids
2.4. SNP Analyses
2.5. Human Listeriosis Cases during the Study Period
3. Results
3.1. Summary of the Bacterial Isolates
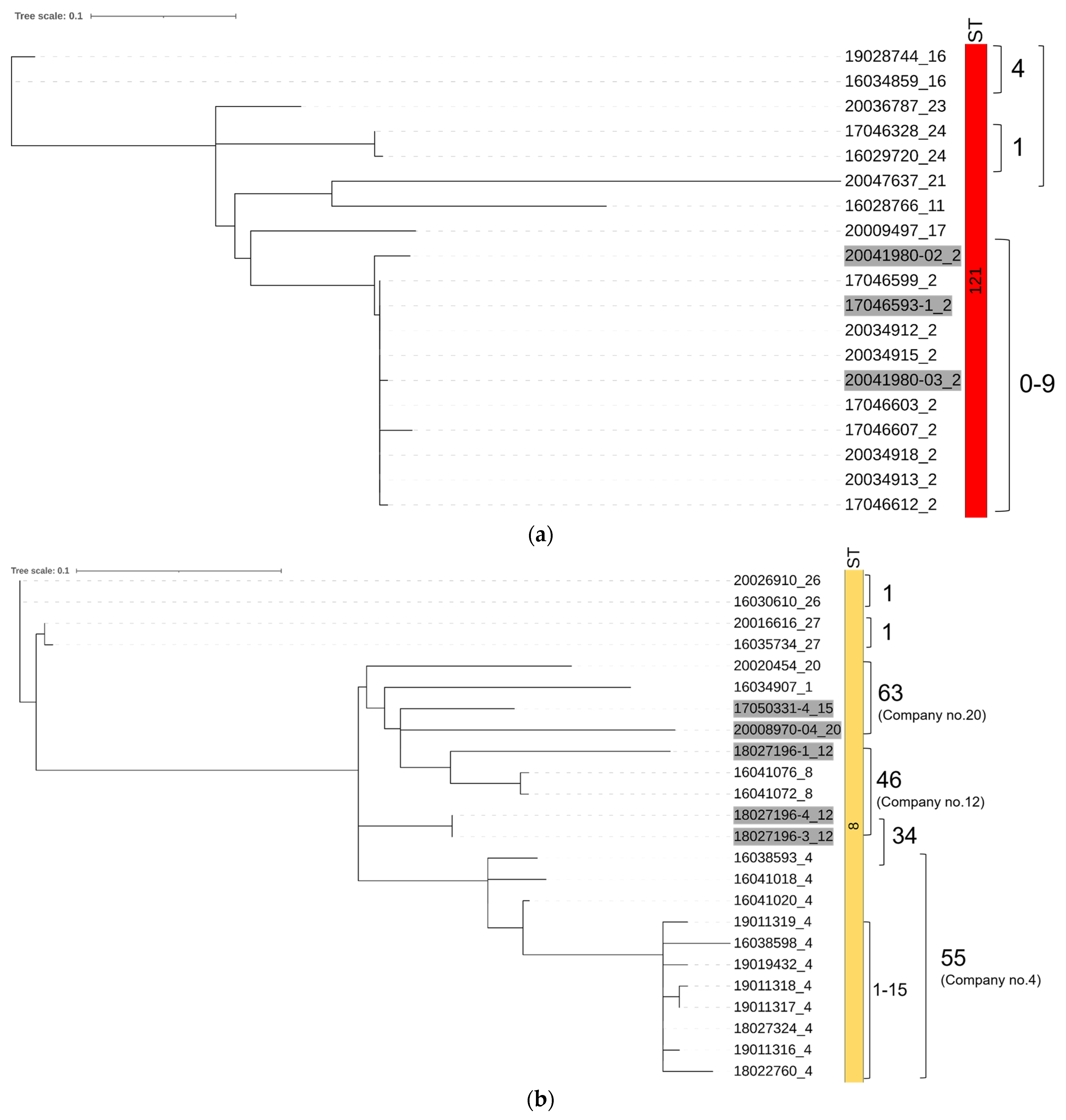
3.2. Distribution of STs and SNP Analyses
3.3. Characterisation of the Selected Virulence, Resistant Genes and Plasmids
3.4. Possible Links to Danish Human Listeriosis Cases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA and ECDC. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar] [CrossRef]
- Annual Report on Zoonoses in Denmark 2019; National Food Institute, Technical University of Denmark: Kgs. Lyngby, Denmark, 2020; Available online: https://www.food.dtu.dk/english/-/media/institutter/foedevareinstituttet/publikationer/pub-2020/rapport-annual-report-2019-final.pdf?la=da&hash=1578450DA14996C6315FF54EF50B800179AE51F9&hash=1578450DA14996C6315FF54EF50B800179AE51F9 (accessed on 19 November 2020).
- Pires, S.M.; Jakobsen, L.S.; Ellis-Iversen, J.; Pessoa, J.; Ethelberg, S. Burden of Disease Estimates of Seven Pathogens Commonly Transmitted Through Foods in Denmark, 2017. Foodborne Pathog. Dis. 2020, 17, 322–339. [Google Scholar] [CrossRef]
- Petran, R.L.; Zottola, E.A. A Study of Factors Affecting Growth and Recovery of Listeria monocytogenes Scott A. J. Food Sci. 1989, 54, 458–460. [Google Scholar] [CrossRef]
- Nolan, D.A.; Chamblin, D.C.; Troller, J.A. Minimal water activity levels for growth and survival of Listeria monocytogenes and Listeria innocua. Int. J. Food Microbiol. 1992, 16, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, P.O.; Bourgeois, R.; Murray, R.G. Psychrotrophic properties of Listeria monocytogenes. Can. J. Microbiol. 1972, 18, 543–551. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef]
- Churchill, K.J.; Sargeant, J.M.; Farber, J.M.; O’connor, A.M. Prevalence of Listeria monocytogenes in select ready-to-eat foods—Deli meat, soft cheese, and packaged salad: A systematic review and meta-analysis. J. Food Prot. 2019, 82, 344–357. [Google Scholar] [CrossRef]
- Vogel, B.F.; Huss, H.H.; Ojeniyi, B.; Ahrens, P.; Gram, A. Elucidation of Listeria monocytogenes Contamination Routes in Cold-Smoked Salmon Processing Plants Detected by DNA-Based Typing Methods. Appl. Environ. Microbiol. 2001, 67, 2586–2595. [Google Scholar] [CrossRef]
- Lassen, S.G.; Ethelberg, S.; Björkman, J.T.; Jensen, T.; Sørensen, G.; Jensen, A.K.; Müller, L.; Nielsen, E.M.; Mølbak, K. Two listeria outbreaks caused by smoked fish consumption—Using whole-genome sequencing for outbreak investigations. Clin. Microbiol. Infect. 2016, 22, 620–624. [Google Scholar] [CrossRef]
- Jensen, A.K.; Nielsen, E.M.; Björkman, J.T.; Jensen, T.; Müller, L.; Persson, S.; Bjerager, G.; Perge, A.; Krause, T.G.; Kiil, K.; et al. Whole-genome Sequencing Used to Investigate a Nationwide Outbreak of Listeriosis Caused by Ready-to-eat Delicatessen Meat, Denmark, 2014. Clin. Infect. Dis. 2016, 63, 64–70. [Google Scholar] [CrossRef]
- Commision regulation (EC) No 2073/2005, of November 2015, on Microbiological Criteria for Foodstuffs. Off. J. Eur. Union 2005, L338, 1–26.
- Commission regulation (EC) No 1441/2007, of 5 December 2007, Amending Regulation (EC) No 2073/2005 on Microbiological Criteria for Foodstuffs. Off. J. Eur. Union 2007, 322, 12–29.
- FAO/WHO. Risk Assessment of Listeria monocytogenes in Ready-to-Eat Foods: Technical Report; Microbiological Risk Assessment Series No.5; FAO: Rome, Italy, 2004. [Google Scholar]
- Maury, M.M.; Tsai, Y.-H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Wagner, E.; Møretrø, T.; Heir, E.; Moen, B.; Rychli, K.; Langsrud, S. Pervasive Listeria monocytogenes Is Common in the Norwegian Food System and Is Associated with Increased Prevalence of Stress Survival and Resistance Determinants. Appl. Environ. Microbiol. 2022, 88, 1–57. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Langsrud, S.; Møretrø, T. In-Depth Longitudinal Study of Listeria monocytogenes ST9 Isolates from the Meat Processing Industry: Resolving Diversity and Transmission Patterns Using Whole-Genome Sequencing. Appl. Environ. Microbiol. 2020, 86, 1–18. [Google Scholar] [CrossRef]
- Jacquet, C.; Doumith, M.; Gordon, J.I.; Martin, P.M.V.; Cossart, P.; Lecuit, M.A. Molecular Marker for Evaluating the Pathogenic Potential of Foodborne Listeria monocytogenes. J. Infect. Dis. 2004, 189, 2094–2100. [Google Scholar] [CrossRef]
- Chen, Y.; Ross, W.H.; Whiting, R.C.; Van Stelten, A.; Nightingale, K.K.; Wiedmann, M.; Scott, V.N. Variation in Listeria monocytogenes Dose Responses in Relation to Subtypes Encoding a Full-Length or Truncated Internalin A. Appl. Environ. Microbiol. 2011, 77, 1171–1180. [Google Scholar] [CrossRef]
- FAO; WHO. Listeria monocytogenes in Ready-to-Eat (RTE) Foods: Attribution, Characterization and Monitoring-Meeting Report; Microbiological Risk Assessment Series No.38; FAO: Rome, Italy, 2022. [Google Scholar]
- Muchaamba, F.; Eshwar, A.K.; Stevens, M.J.A.; Stephan, R.; Tasara, T. Different Shades of Listeria monocytogenes: Strain, Serotype, and Lineage-Based Variability in Virulence and Stress Tolerance Profiles. Front. Microbiol. 2022, 12, 3736. [Google Scholar] [CrossRef]
- Carpentier, B.; Barre, L. Guidelines on Sampling the Food Processing Area and Equipment for the Detection of Listeria monocytogenes Version 3. 2012. Available online: http://www.ansespro.fr/eurl-listeria/ (accessed on 31 August 2021).
- Clausen, P.T.L.C.; Aarestrup, F.M.; Lund, O. Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinform. 2018, 19, 307. [Google Scholar] [CrossRef]
- Hasman, H.; Saputra, D.; Sicheritz-Ponten, T.; Lund, O.; Svendsen, C.A.; Frimodt-Møller, N.; Aarestrup, F.M. Rapid Whole-Genome Sequencing for Detection and Characterization of Microorganisms Directly from Clinical Samples. J. Clin. Microbiol. 2014, 52, 139–146. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of Methods for Genomic Taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontéen, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef]
- Seemann, T. Abricate. Available online: https://github.com/tseemann/abricate (accessed on 1 October 2021).
- Carattoli, A.; Zankari, E.; Garcìa-Fernandez, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids. Antimicrob using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Annual Report on Zoonoses in Denmark 2017; National Food Institute, Technical University of Denmark: Kongens Lyngby, Denmark, 2018; Available online: https://www.food.dtu.dk/english/-/media/institutter/foedevareinstituttet/publikationer/pub-2018/rapport-annual-report-on-zoonoses-2017.pdf?la=da&hash=E19140DF81A8C7D613C79D3BEA7673D24D844CD5&hash=E19140DF81A8C7D613C79D3BEA7673D24D844CD5 (accessed on 6 July 2021).
- Annual Report on Zoonoses in Denmark 2016; National Food Institute, Technical University of Denmark: Kongens Lyngby, Denmark, 2017; Available online: https://www.food.dtu.dk/english/-/media/institutter/foedevareinstituttet/publikationer/pub-2017/annual-report-on-zoonoses-in-denmark-2016-ver3.pdf?la=da&hash=54DC8AEA6C2E5377054CD7B6BC14B89A311F73A0&hash=54DC8AEA6C2E5377054CD7B6BC14B89A311F73A0 (accessed on 31 May 2021).
- Annual Report on Zoonoses in Denmark 2018; National Food Institute, Technical University of Denmark: Kongens Lyngby, Denmark, 2019; Available online: https://www.food.dtu.dk/english/-/media/institutter/foedevareinstituttet/publikationer/pub-2019/annual-report-on-zoonoses-2018-v2-final.pdf?la=da&hash=CEFDA3E9ACC848EA5AA0977C65625DC65F1DB7FD&hash=CEFDA3E9ACC848EA5AA0977C65625DC65F1DB7FD (accessed on 10 February 2021).
- Annual Report on Zoonoses in Denmark 2020; National Food Institute, Technical University of Denmark: Kongens Lyngby, Denmark, 2021; Available online: https://www.food.dtu.dk/english/-/media/institutter/foedevareinstituttet/publikationer/pub-2021/rapport-annual-report-on-zoonoses-2020.pdf?la=da&hash=72A8B11B4FC5E19F20AEED3B7FCB338CE9C8E7F0&hash=72A8B11B4FC5E19F20AEED3B7FCB338CE9C8E7F0 (accessed on 6 July 2021).
- Institut Pasteur InlA Premature Stop Codons. Available online: https://bigsdb.pasteur.fr/_nuxt/img/inlA_PMSC.4bb4563.pdf (accessed on 21 May 2021).
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole genome-based population biology and epidemiological surveillance of Listeria monocytogenes. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef]
- Rousseaux, S.; Olier, M.; Lemaître, J.P.; Piveteau, P.; Guzzo, J. Use of PCR-Restriction Fragment Length Polymorphism of inlA for Rapid Screening of Listeria monocytogenes Strains Deficient in the Ability to Invade Caco-2 Cells. Appl. Environ. Microbiol. 2004, 70, 2180–2185. [Google Scholar] [CrossRef]
- Olier, M.; Pierre, F.; Rousseaux, S.; Lemaître, J.-P.; Rousset, A.; Piveteau, P.; Guzzo, J. Expression of Truncated Internalin A Is Involved in Impaired Internalization of Some Listeria monocytogenes Isolates Carried Asymptomatically by Humans. Infect. Immun. 2003, 71, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Ebner, R.; Stephan, R.; Althaus, D.; Brisse, S.; Maury, M.; Tasara, T. Phenotypic and genotypic characteristics of Listeria monocytogenes strains isolated during 2011–2014 from different food matrices in Switzerland. Food Control. 2015, 57, 321–326. [Google Scholar] [CrossRef]
- Zilelidou, E.; Karmiri, C.-V.; Zoumpopoulou, G.; Mavrogonatou, E.; Kletsas, D.; Tsakalidou, E.; Papadimitriou, K.; Drosinos, E.; Skandamis, P. Listeria monocytogenes Strains Underrepresented during Selective Enrichment with an ISO Method Might Dominate during Passage through Simulated Gastric Fluid and In Vitro Infection of Caco-2 Cells. Appl. Environ. Microbiol. 2016, 82, 6846–6858. [Google Scholar] [CrossRef]
- Wang, Y.; Pettengill, J.; Pightling, A.; Timme, R.; Allard, M.; Strain, E.; Rand, H. Genetic Diversity of Salmonella and Listeria Isolates from Food Facilities. J. Food Prot. 2018, 81, 2082–2089. [Google Scholar] [CrossRef]
- Løvdal, T.; Brandal, L.T.; Sundaram, A.Y.M.; Naseer, U.; Roth, B.; Lunestad, B.T. Small-Scale Comparative Genomic Analysis of Listeria monocytogenes Isolated from Environments of Salmon Processing Plants and Human Cases in Norway. Hygiene 2021, 1, 43–55. [Google Scholar] [CrossRef]
- Fagerlund, A.; Idland, L.; Heir, E.; Møretrø, T.; Aspholm, M.; Lindbäck, T.; Langsrud, S. Whole-Genome Sequencing analysis of Listeria monocytogenes from rural, urban, and farm environments in Norway: Genetic diversity, persistence, and relation to clinical and food isolates. Appl. Environ. Microbiol. 2022, 88, e02136-21. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes Persistence in Food-Associated Environments: Epidemiology, Strain Characteristics, and Implications for Public Health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef]
- Harrand, A.S.; Jagadeesan, B.; Baert, L.; Wiedmann, M.; Orsi, R.H. Evolution of Listeria monocytogenes in a Food Processing Plant Involves Limited Single-Nucleotide Substitutions but Considerable Diversification by Gain and Loss of Prophages. Appl. Environ. Microbiol. 2020, 86, e02493-19. [Google Scholar] [CrossRef]
- Statens Serum Institute Overvågning i Tal, Grafer Og Kort (Monitoring in Numbers, Grafs, and Maps). Available online: https://statistik.ssi.dk/ (accessed on 13 December 2022).
- Maia, R.L.; Teixeira, P.; Mateus, T.L. Risk communication strategies (on listeriosis) for high-risk groups. Trends Food Sci. Technol. 2019, 84, 68–70. [Google Scholar] [CrossRef]
- Dalgaard, P.; Mejlholm, O. Modeling Growth of Listeria and Lactic Acid Bacteria in Food Environments. Methods Mol. Biol. 2019, 1918, 247–264. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | 2016 | 2020 | ||
|---|---|---|---|---|
| Product Type of the Company | Meat | Fish | Meat | Fish |
| No. of samples | 251 | 175 | 181 | 170 |
| No. of companies | 22 | 17 | 18 | 16 |
| No. of samples per company | 6–25 | 10–20 | ||
| No. of samples positive for L. monocytogenes by culture | 15 (6.0) a | 17 (9.7) | 10 (5.5) | 10 (5.9) |
| No. of companies positive for L. monocytogenes by culture | 8 (36.3) | 9 (52.9) | 7 (38.9) | 4 (25.0) |
| 2017 | 2018 | 2019 | 2020 | ||||
| Sample source | Env | Prod | Env | Prod | Env | Prod | Prod |
| No. isolates with WGS data | 6 | 4 | 2 | 3 | 6 | 5 | 9 |
| No. companies with L. monocytogenes | 2 | 3 | 1 | 1 | 2 | 4 | 3 |
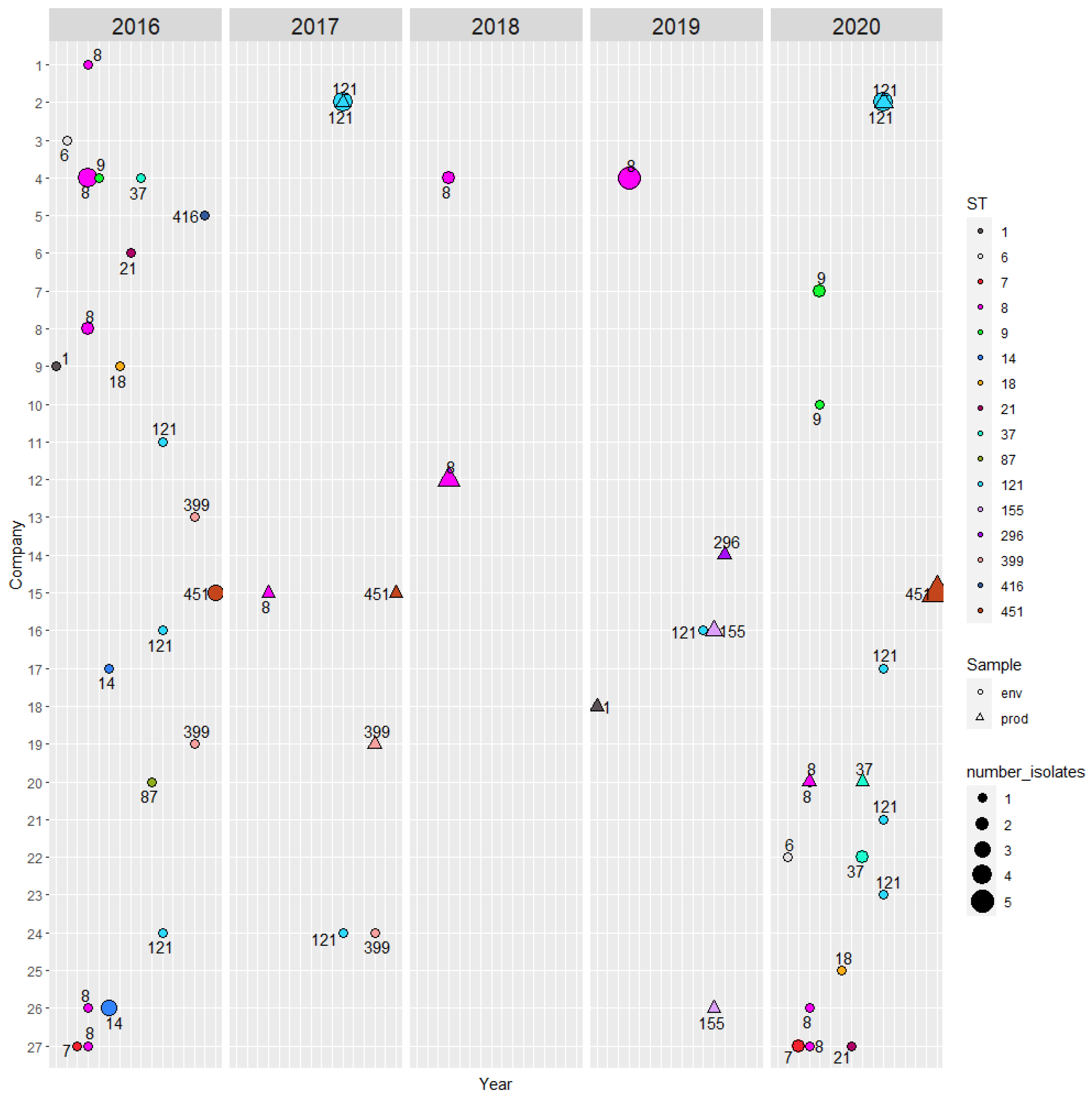
| ST | Total No. Isolates | Lineage | Company ID | Company Type | No. Isolates | ST | Total No. Isolates | Lineage | Company ID | Company Type | No. Isolates |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ST1 | 2 | I | 9 18 | Meat Fish | 1 1 | ST37 | 4 | II | 4 20 22 | Fish Meat Meat | 1 1 2 |
| ST6 | 2 | I | 3 22 | Meat Meat | 1 1 | ST87 | 1 | I | 20 | Meat | 1 |
| ST7 | 3 | II | 27 | Meat | 3 | ST121 | 19 | II | 2 11 16 17 21 23 24 | Fish Fish Meat Meat Meat Meat Fish | 11 1 2 1 1 1 2 |
| ST8 | 24 | II | 1 4 5 8 12 20 26 27 | Fish Fish Meat Meat Meat Meat Meat Meat | 1 11 2 3 1 2 2 2 | ST155 | 3 | II | 16 26 | Meat Meat | 2 1 |
| ST9 | 4 | II | 4 7 10 | Fish Meat Meat | 1 2 1 | ST296 | 1 | I | 14 | Meat | 1 |
| ST14 | 4 | II | 17 26 | Meat Meat | 1 3 | ST399 | 4 | II | 13 18 24 | Meat Fish Fish | 1 2 1 |
| ST18 | 2 | II | 9 25 | Fish Fish | 1 1 | ST416 | 1 | I | 5 | Meat | 1 |
| ST21 | 2 | II | 6 27 | Meat Meat | 1 1 | ST451 | 9 | II | 15 | Meat | 9 |
| Year | ST Type (Lineage a) | Source |
|---|---|---|
| 2016 | ST4 (I) | Cold cuts of meat |
| 2016 | ST6 (I) | Unknown |
| 2017 | ST1 (I) | Unknown |
| 2017 | ST1247 (II) | Unknown |
| 2017 | ST8 (II) | Imported smoked salmon |
| 2017 | ST55 (I) | Unknown |
| 2017 | ST6 (I) | Imported sweet corn |
| 2018 | ST20 (II) | Unknown |
| 2018 | ST8 (II) | Unknown |
| 2019 | ST1 (I) | Salads |
| 2020 | ST7#7 (II) | Unknown |
| 2020 | ST394#1 (II) | Hot-smoked trout |
| 2020 | ST451#2 (II) b | Hot-smoked fish products |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takeuchi-Storm, N.; Hansen, L.T.; Nielsen, N.L.; Andersen, J.K. Presence and Persistence of Listeria monocytogenes in the Danish Ready-to-Eat Food Production Environment. Hygiene 2023, 3, 18-32. https://doi.org/10.3390/hygiene3010004
Takeuchi-Storm N, Hansen LT, Nielsen NL, Andersen JK. Presence and Persistence of Listeria monocytogenes in the Danish Ready-to-Eat Food Production Environment. Hygiene. 2023; 3(1):18-32. https://doi.org/10.3390/hygiene3010004
Chicago/Turabian StyleTakeuchi-Storm, Nao, Lisbeth Truelstrup Hansen, Niels Ladefoged Nielsen, and Jens Kirk Andersen. 2023. "Presence and Persistence of Listeria monocytogenes in the Danish Ready-to-Eat Food Production Environment" Hygiene 3, no. 1: 18-32. https://doi.org/10.3390/hygiene3010004
APA StyleTakeuchi-Storm, N., Hansen, L. T., Nielsen, N. L., & Andersen, J. K. (2023). Presence and Persistence of Listeria monocytogenes in the Danish Ready-to-Eat Food Production Environment. Hygiene, 3(1), 18-32. https://doi.org/10.3390/hygiene3010004