


Stress and Disease Resistance in Crayfish (Procambarus clarkii) Breed “Huachizhen-1”


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. DNA Extraction, Polymerase Chain Reaction (PCR) Amplification, and Electrophoresis
2.3. Stress Resistance of Crayfish
2.4. Pathogen Challenge for Crayfish
2.5. Data Analysis
3. Results
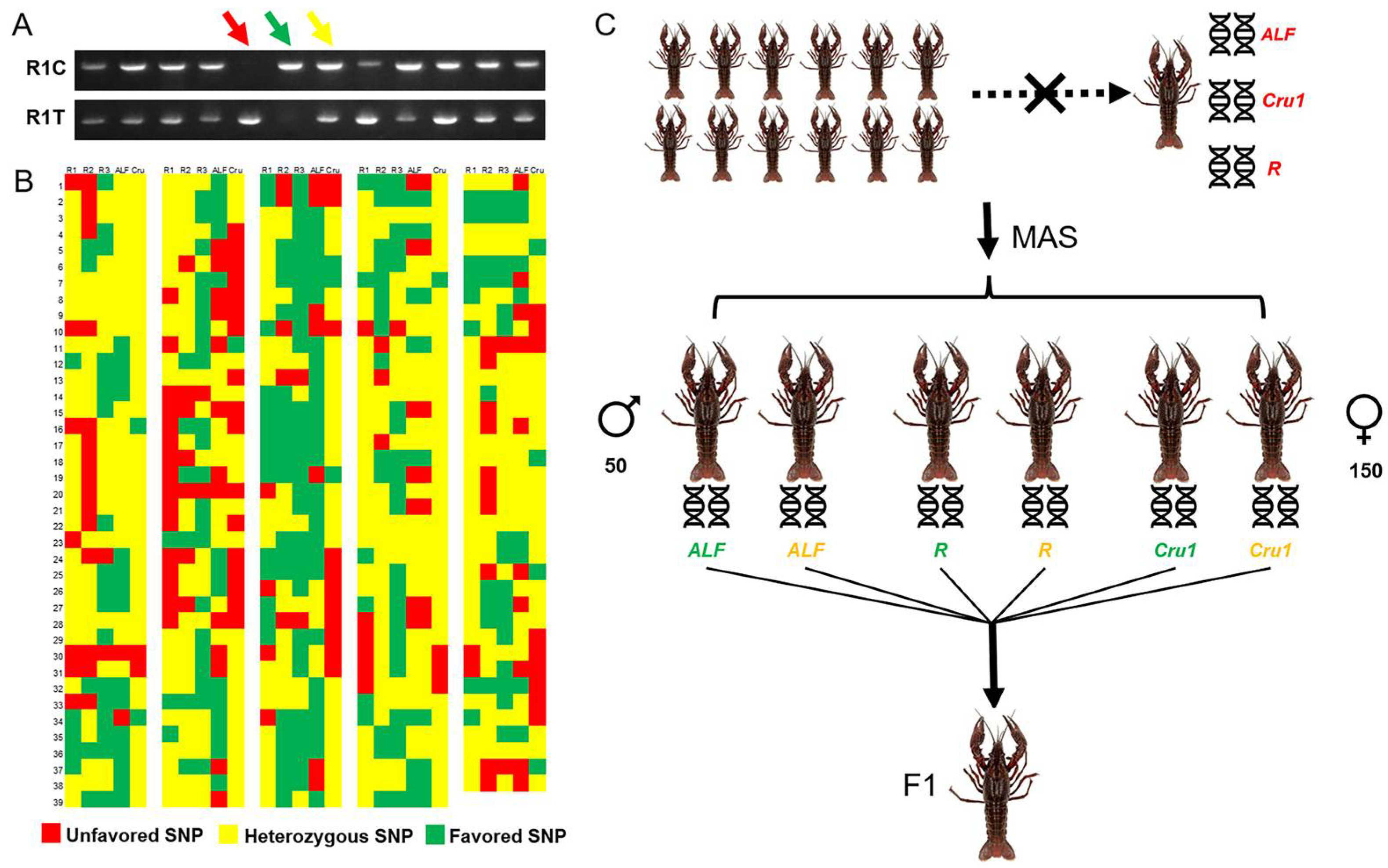
3.1. Selection of Parents for Breeding “Huachizhen-1” Crayfish
3.2. No Significant Difference in Genetic Diversity between the SG and USG Populations
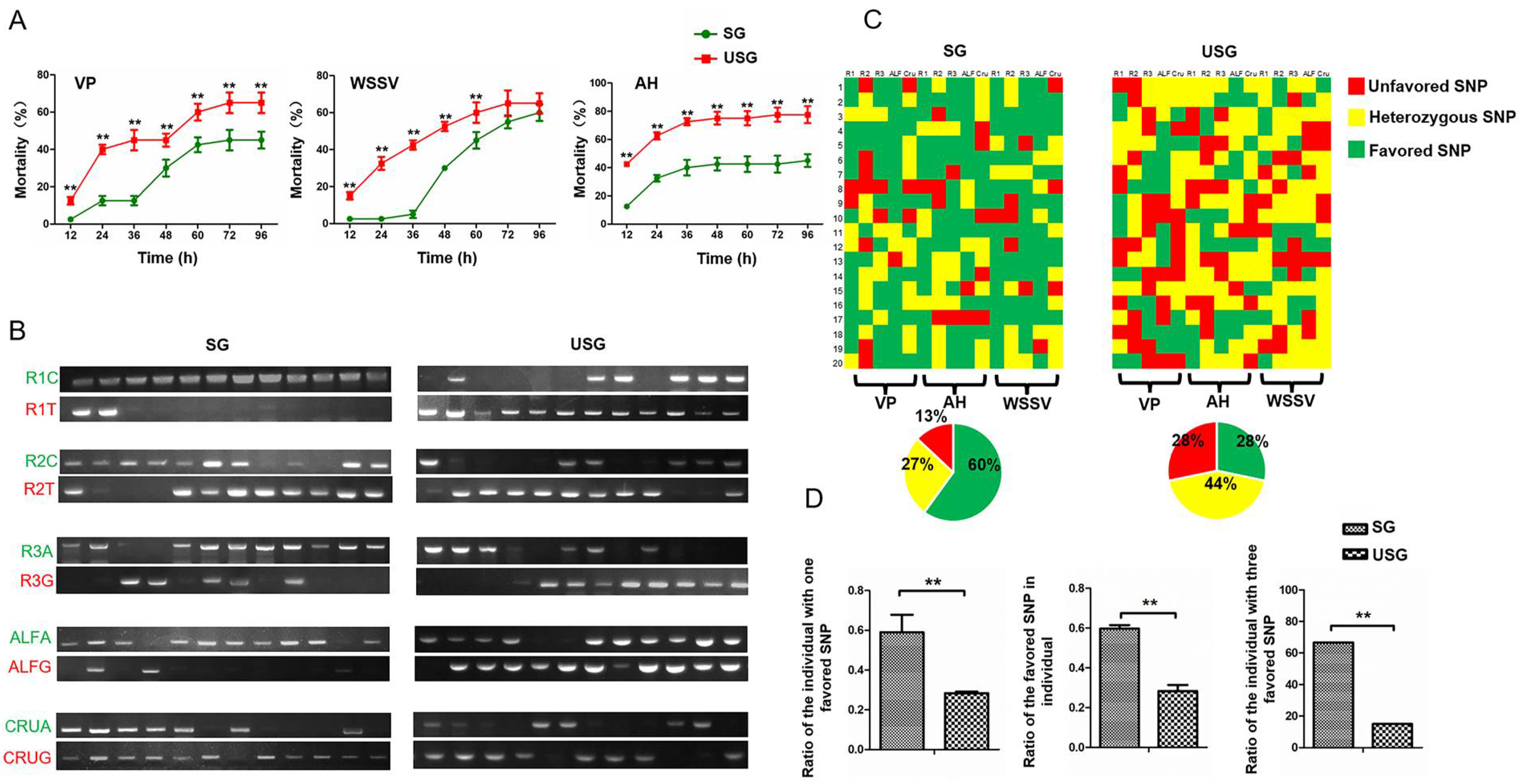
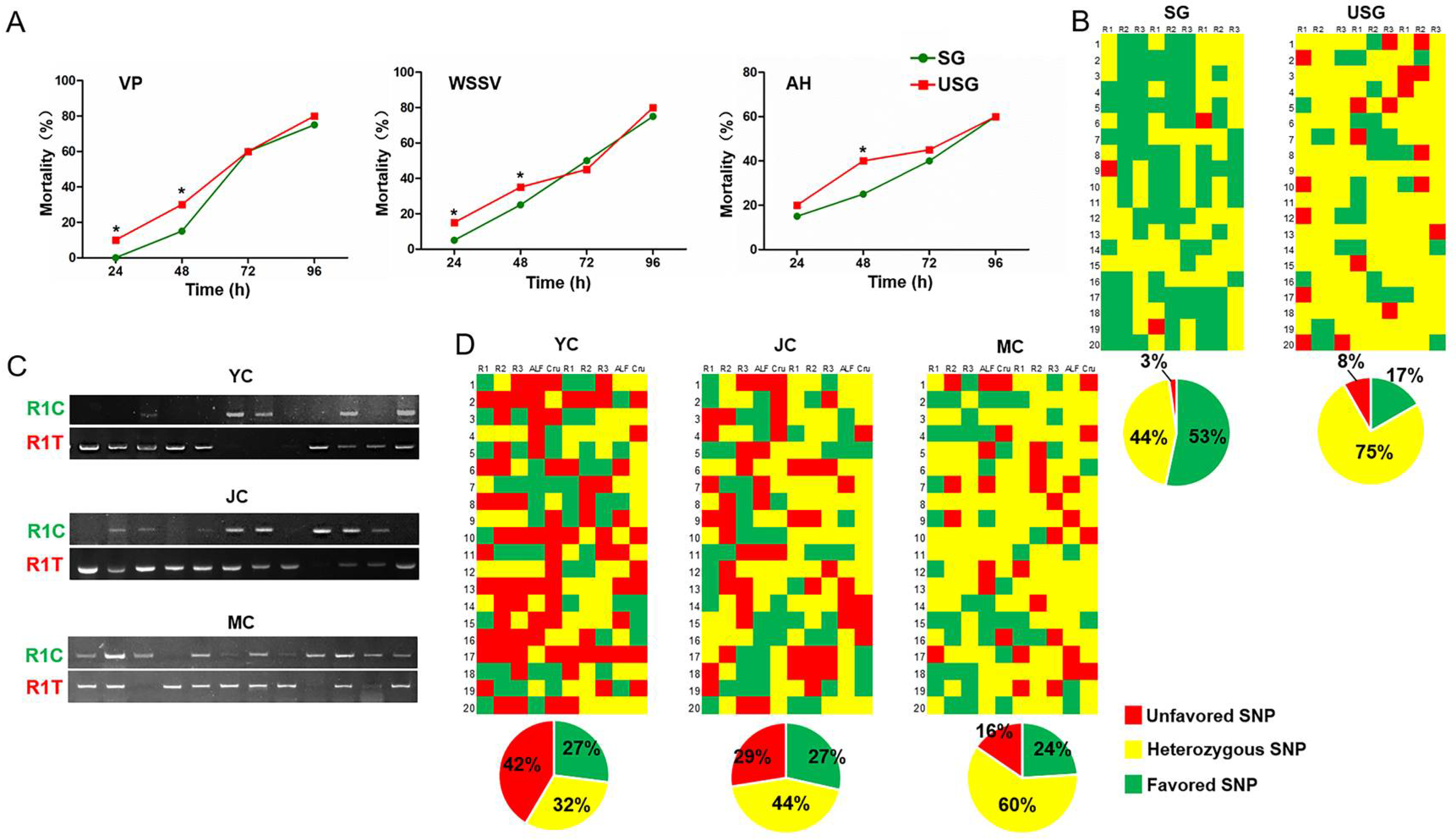
3.3. Crayfish in SG Showed Stronger Stress Resistance Than Those in USG
3.4. Crayfish in SG Showed Stronger Disease Resistance Than Those in USG
3.5. Elimination of Crayfish with Unfavoured Genotype and Retention of Those with the Heterozygous Genotype under Natural Conditions
4. Discussion
4.1. Immune Genes Can Be Applied in Stress- and Disease-Resistance Breeding in Crayfish
4.2. Stress- and Disease-Resistant Crayfish Bred Using MAS
4.3. Hybridization Breeding Can Improve Stress and Disease Resistance in Crayfish
4.4. Application and Prospects of “Huachizhen-1”
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barbaresi, S.; Fani, R.; Gherardi, F.; Mengoni, A.; Souty-Grosset, C. Genetic variability in European populations of an invasive American crayfish: Preliminary results. Biol. Invasions 2003, 5, 269–274. [Google Scholar] [CrossRef]
- Yue, G.H.; Feng, J.B.; Xia, J.H.; Cao, S.Y.; Wang, C.M. Inferring the invasion mechanisms of the red swamp crayfish in China using mitochondrial DNA sequences. Aquac. Fish. 2021, 6, 35–41. [Google Scholar] [CrossRef]
- Li, Y.; Guo, X.; Chen, L.; Bai, X.; Wei, X.; Zhou, X.; Huang, S.; Wang, W. Inferring invasion history of red swamp crayfish (Procambarus clarkii) in China from mitochondrial control region and nuclear intron sequences. Int. J. Mol. Sci. 2015, 16, 14623–14639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Peng, G.H.; Zhang, H.Y.; Peng, B.; Bai, X.F. Sex differences in the disease resistance of red swamp crayfish (Procambarus clarkii). Aquaculture 2022, 548, 737712. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, H.; Huang, Z.; Ye, X.; Zhang, L.; Li, Q.; Zhao, Z.; Su, X.; Liu, G.; Du, J.; et al. Differential transcriptomic analysis of crayfish (Procambarus clarkii) from a rice coculture system challenged by Vibrio parahaemolyticus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 36, 100741. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.N.; Huang, L.; Wang, S.Y.; Li, Y.T.; Tang, Y.Y.; Zhang, D.Z.; Tang, B.P.; Yang, H.; He, J.X.; Ding, F.; et al. Transcriptome analysis of differentially expressed genes in the red swamp crayfish Procambarus clarkii challenged with Aeromonas hydrophila. Fish Shellfish. Immunol. 2021, 119, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Zhu, F.; Xu, Y. WSSV proteins and DNA genome released by ultrasonic rupture can infect crayfish as effectively as intact virions. J. Virol. Methods 2020, 283, 113917. [Google Scholar] [CrossRef] [PubMed]
- Longshaw, M. Diseases of crayfish: A review. J. Invertebr. Pathol. 2011, 106, 54–70. [Google Scholar] [CrossRef]
- Chang, Y.H.; Kumar, R.; Ng, T.H.; Wang, H.C. What vaccination studies tell us about immunological memory within the innate immune system of cultured shrimp and crayfish. Dev. Comp. Immunol. 2018, 80, 53–66. [Google Scholar] [CrossRef]
- Oficialdegui, F.J.; Clavero, M.; Sánchez, M.I.; Green, A.J.; Boyero, L.; Michot, T.C.; Klose, K.; Kawai, T.; Lejeusne, C. Unravelling the global invasion routes of a worldwide invader, the red swamp crayfish (Procambarus clarkii). Freshw. Biol. 2019, 64, 1382–1400. [Google Scholar] [CrossRef]
- Sun, J.X.; Peng, G.H.; Xiong, L.J.; Tan, C.; Li, Y.; Bai, X.F. Genome-wide SSR marker development and application in genetic diversity analysis of red swamp crayfish Procambarus clarkii in China. Crustaceana 2021, 94, 189–205. [Google Scholar] [CrossRef]
- Xing, Z.J.; Jian, H.C.; Lu, W. Genetic diversity analysis of eight Procambarus clarkii stocks in Jiangsu Province based on microsatellites. Acta Shanghai Ocean. Univ. 2014, 23, 656–662. [Google Scholar]
- Wendy, C.P.L.; Mohammad, A.L.; Mohd, Y.R.; Mohd, R.I.; Adam, P. Advances to improve the eating and cooking qualities of rice by marker-assisted breeding. Crit. Rev. Biotechnol. 2016, 36, 87–98. [Google Scholar]
- Mwamahonje, A.; Eleblu, J.S.Y.; Ofori, K.; Deshpande, S.; Feyissa, T.; Tongoona, P. Drought tolerance and application of marker-assisted selection in Sorghum. Biology 2021, 10, 1249. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Gao, T.; Xu, Y.; Li, X.; Li, J.; Lin, H.; Yan, W.; Pan, J.; Tang, J. A chromosome-level reference genome of red swamp crayfish Procambarus clarkii provides insights into the gene families regarding growth or development in crustaceans. Genomics 2021, 113, 3274–3284. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Lee, S.Y.; Kim, Y.A.; Andrén, T.; Söderhäll, I. Antibacterial peptides in hemocytes and hematopoietic tissue from freshwater crayfish Pacifastacus leniusculus: Characterization and expression pattern. Dev. Comp. Immunol. 2007, 31, 441–455. [Google Scholar] [CrossRef]
- Zhang, X.W.; Han, K.K.; Yang, C.H.; Xia, X.H.; Ren, Q.; Zhang, H.W. A stomach-specific lectin plays a crucial role in the innate immunity of red swamp crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2020, 103, 285–292. [Google Scholar] [CrossRef]
- Ren, X.; Peng, G.H.; Peng, B.; Tan, Y.F.; Bai, X.F. Robust strategy for disease resistance and increasing production breeding in red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2022, 122, 57–66. [Google Scholar] [CrossRef]
- Chen, D.; Wang, H. Redclaw crayfish (Cherax quadricarinatus) responds to Vibrio parahaemolyticus infection by activating toll and immune deficiency signaling pathways and transcription of associated immune response genes. Fish Shellfish Immunol. 2022, 127, 611–622. [Google Scholar] [CrossRef]
- Du, Z.Q.; Li, B.; Shen, X.L.; Wang, K.; Du, J.; Yu, X.D.; Yuan, J.J. A new antimicrobial peptide isoform, Pc-crustin 4 involved in antibacterial innate immune response in fresh water crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2019, 94, 861–870. [Google Scholar] [CrossRef]
- Yin, C.M.; Pan, X.Y.; Cao, X.T.; Li, T.; Zhang, Y.H.; Lan, J.F. A crayfish ALF inhibits the proliferation of microbiota by binding to RPS4 and MscL of E. coli. Dev. Comp. Immunol. 2021, 121, 104106. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.F.; Peng, G.H.; Xiong, L.J.; Peng, B.; Wu, Y.B.; Song, C.W.; Bai, X.F. Genetic diversity and structure analysis of 13 red swamp crayfish (Procambarus clarkii) populations in Yangtze River basin. J. Huazhong Agric. Univ. 2020, 39, 33–39. [Google Scholar]
- Yeh, F.C.; Boyle, T.J.B. Population genetic analysis of codmminant and dominant markers and quantitative traits. Belg. J. Bot. 1997, 129, 129–157. [Google Scholar]
- Paaby, A.B.; Rockman, M.V. The many faces of pleiotropy. Trends Genet. 2013, 29, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, M.R.; Valpuesta, J.M. Hsp70 chaperone: A master player in protein homeostasis. F1000Research 2018, 7, 1497. [Google Scholar] [CrossRef] [Green Version]
- Fan, K.P.; Hua, X.T.; Liu, Y.F.; Zhang, Z.Q.; Li, X.H.; Liu, Y.; Liu, P.F. HSP70 gene expression responses to the temperature stress in pufferfish (Takifugu rubripes). Biosci. Biotechnol. Biochem. 2021, 85, 1088–1096. [Google Scholar] [CrossRef]
- Roufayel, R.; Kadry, S. Molecular chaperone HSP70 and key regulators of apoptosis-a review. Curr. Mol. Med. 2019, 19, 315–325. [Google Scholar] [CrossRef]
- Zhang, L.; Song, Z.W.; Zhong, S.; Gan, J.H.; Liang, H.W.; Yu, Y.L.; Wu, G.; He, L. Acute hypoxia and reoxygenation induces oxidative stress, glycometabolism, and oxygen transport change in red swamp crayfish (Procambarus clarkii): Application of transcriptome profiling in assessment of hypoxia. Aquac. Rep. 2022, 23, 101029. [Google Scholar] [CrossRef]
- Lin, F.Y.; Gao, Y.; Wang, H.; Zhang, Q.X.; Zeng, C.L.; Liu, H.P. Identification of an anti-lipopolysacchride factor possessing both antiviral and antibacterial activity from the red claw crayfish Cherax quadricarinatus. Fish Shellfish Immunol. 2016, 57, 213–221. [Google Scholar] [CrossRef]
- Zhao, B.R.; Zheng, Y.; Gao, J.; Wang, X.W. Maturation of an antimicrobial peptide inhibits aeromonas hydrophila infection in crayfish. J. Immunol. 2020, 204, 487–497. [Google Scholar] [CrossRef]
- Hasan, N.; Choudhary, S.; Naaz, N.; Sharma, N.; Laskar, R.A. Recent advancements in molecular marker-assisted selection and applications in plant breeding programmes. J. Genet. Eng. Biotechnol. 2021, 19, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.; Barbosa, A.C.O.; Diniz, R.P.; Oliveira, E.J.; Ferreira, C.F. Molecular marker assisted selection for increasing inbreeding in S1 populations of cassava. An. Acad. Bras. Cienc. 2018, 90, 3853–3869. [Google Scholar] [CrossRef] [PubMed]
- Longin, C.F.; Mühleisen, J.; Maurer, H.P.; Zhang, H.; Gowda, M.; Reif, J.C. Hybrid breeding in autogamous cereals. Theor. Appl. Genet. 2012, 125, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.; Gho, C.; Leafgren, R.; Tang, T.; Messina, C. Breeding drought-tolerant maize hybrids for the US corn-belt: Discovery to product. J. Exp. Bot. 2014, 65, 6191–6204. [Google Scholar] [CrossRef]
- Li, Y.H. Invasion Genetics of Procambarus Clarkii in China; Huazhong Agricultural University: Hubei, China, 2013. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | PIC | Ho | He |
|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | |
| SG | 0.36 ± 0.15 | 0.43 ± 0.27 | 0.41 ± 0.18 |
| USG | 0.37 ± 0.14 | 0.45 ± 0.22 | 0.44 ± 0.18 |
| p value | 0.89 | 0.97 | 0.82 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, X.; Xiong, L.; Tan, Y.; Liu, X.; Zhu, X.; Bai, X. Stress and Disease Resistance in Crayfish (Procambarus clarkii) Breed “Huachizhen-1”. Aquac. J. 2023, 3, 7-17. https://doi.org/10.3390/aquacj3010002
Ren X, Xiong L, Tan Y, Liu X, Zhu X, Bai X. Stress and Disease Resistance in Crayfish (Procambarus clarkii) Breed “Huachizhen-1”. Aquaculture Journal. 2023; 3(1):7-17. https://doi.org/10.3390/aquacj3010002
Chicago/Turabian StyleRen, Xin, Lijing Xiong, Yunfei Tan, Xiaoyu Liu, Xi Zhu, and Xufeng Bai. 2023. "Stress and Disease Resistance in Crayfish (Procambarus clarkii) Breed “Huachizhen-1”" Aquaculture Journal 3, no. 1: 7-17. https://doi.org/10.3390/aquacj3010002
APA StyleRen, X., Xiong, L., Tan, Y., Liu, X., Zhu, X., & Bai, X. (2023). Stress and Disease Resistance in Crayfish (Procambarus clarkii) Breed “Huachizhen-1”. Aquaculture Journal, 3(1), 7-17. https://doi.org/10.3390/aquacj3010002