


Influence of Body Weight and Water Temperature on Growth in Ragworm Hediste diversicolor
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Statistical Analyses and Simulations
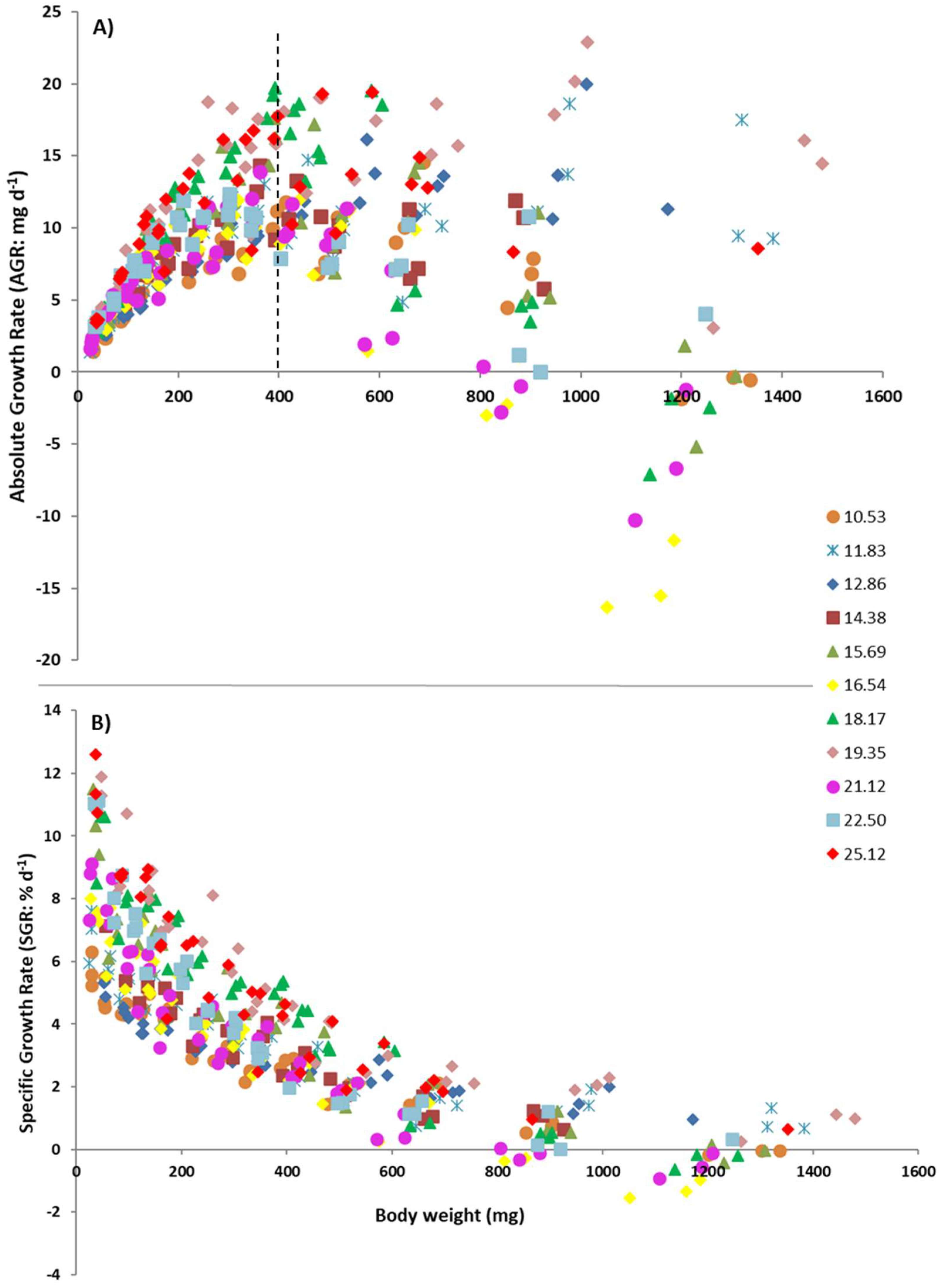
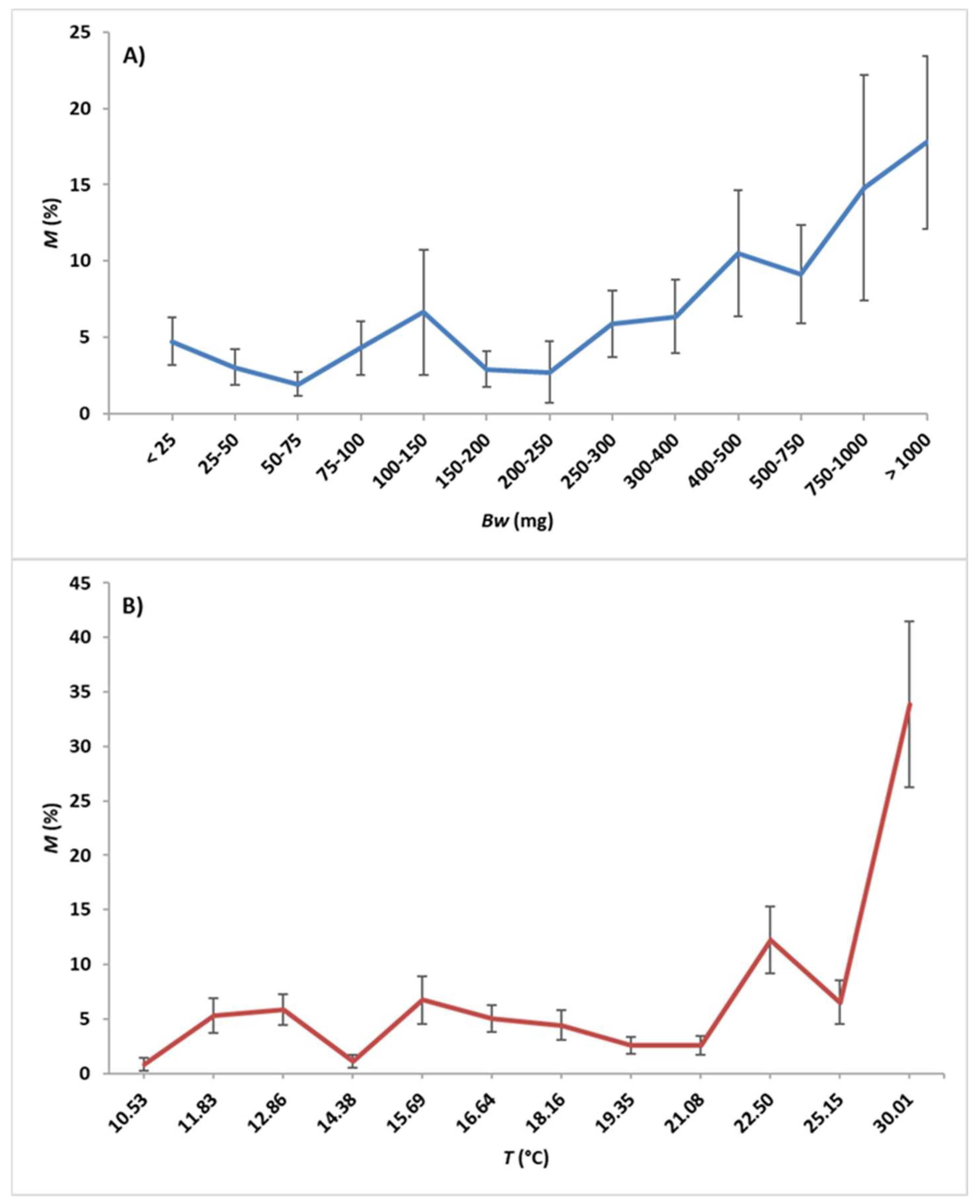
3. Results
3.1. Modeling Relationships between Variables
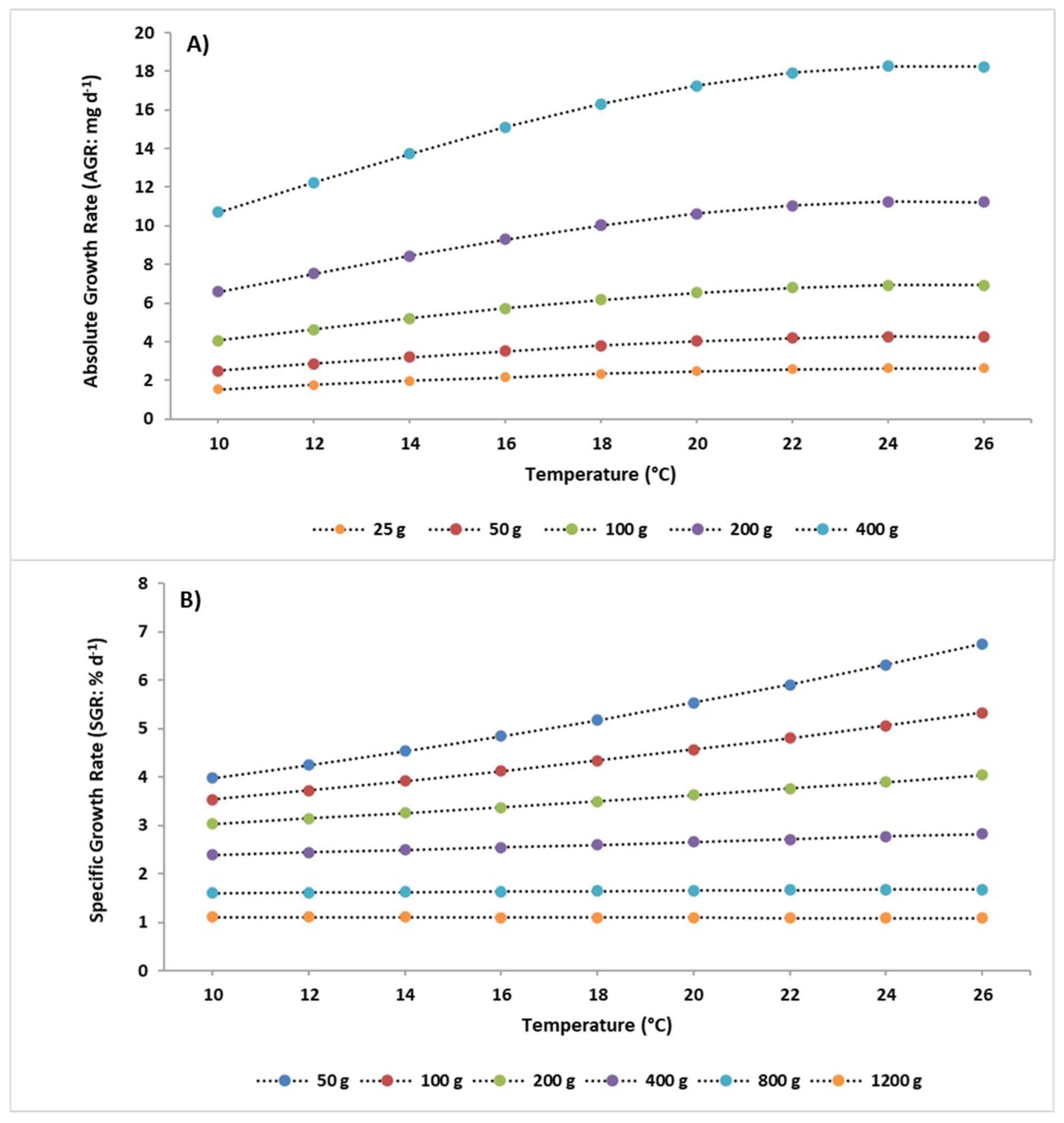
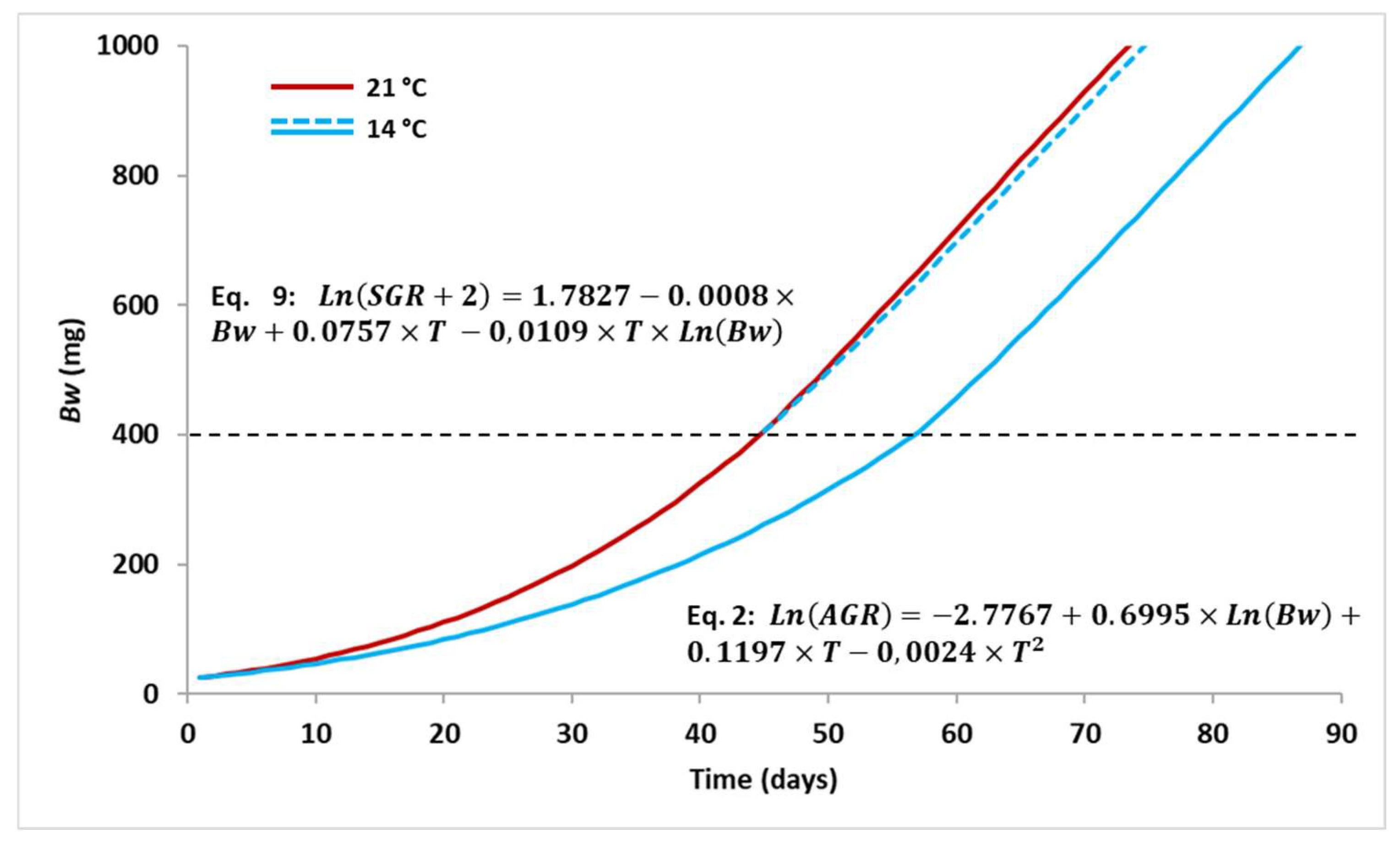
3.2. Simulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scaps, P. A review of the biology, ecology and potential use of the common ragworm Hediste diversicolor (O.F. Müller) (Annelida: Polychaeta). Hydrobiologia 2002, 470, 203–218. [Google Scholar] [CrossRef]
- Fidalgo e Costa, P.; Narciso, L.; Cancela da Fonseca, L. Growth, survival and fatty acid profile of Nereis diversicolor (O. F. Muller, 1776) fed on six different diets. Bull. Mar. Sci. 2000, 67, 337–343. [Google Scholar]
- Santos, A.; Granada, L.; Baptista, T.; Anjos, C.; Simões, T.; Tecelão, C.; Fidalgo e Costa, P.; Costa, J.L.; Pombo, A. Effect of three diets on the growth and fatty acid profile of the common ragworm Hediste diversicolor (O.F. Müller, 1776). Aquaculture 2016, 465, 37–42. [Google Scholar] [CrossRef]
- Marques, B.; Calado, R.; Lillebø, A.I. New species for the biomitigation of a super-intensive marine fish farm effluent: Combined use of polychaete-assisted sand filters and halophyte aquaponics. Sci. Tot. Environ. 2017, 599–600, 1922–1928. [Google Scholar] [CrossRef] [PubMed]
- Pajand, Z.O.; Soltani, M.; Bahmani, M.; Kamali, A. The role oof polychaete Nereis diversicolor in bioremediation of wastewater and its growth performance and fatty acid composition in an integrated culture system with Huso huso (Linnaeus, 1758). Aquac. Res. 2017, 48, 5271–5279. [Google Scholar] [CrossRef]
- Gilbert, F.; Kristensen, E.; Aller, R.C.; Banta, G.T.; Archambault, P.; Belley, R.; Bellucci, L.G.; Calder, L.; Cuny, P.; de Montaudouin, X.; et al. Sediment reworking by the burrowing polychaete Hediste diversicolor modulated by environmental and biological factors across the temperate North Atlantic. A tribute to Gaston Desrosiers. J. Exp. Mar. Biol. Ecol. 2021, 541, 151588. [Google Scholar] [CrossRef]
- Wang, H.; Hagemann, A.; Reitan, K.I.; Ejlertsson, J.; Wollan, H.; Handå, A.; Malzahn, A.M. Potential of the polychaete Hediste diversicolor fed on aquaculture and biogas side streams as an aquaculture food source. Aquac. Environ. Interac. 2019, 11, 551–562. [Google Scholar] [CrossRef] [Green Version]
- García-Alonso, J.; Müller, C.T.; Hardege, J.D. Influence of food regimes and seasonality on fatty acid composition in the ragworm. Aquat. Biol. 2008, 4, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Yousefi-Garakouei, M.; Kamali, A.; Soltani, M. Effects of rearing density on growth, fatty acid profile and bioremediation ability of polychaete Nereis diversicolor in an integrated aquaculture system with rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2019, 50, 725–735. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Santos, A.; Cremades, J.; Calado, R. Performance of polychaete assisted sand filters under contrasting nutrient loads in an integrated multi-trophic aquaculture (IMTA) system. Sci. Rep. 2020, 10, 20871. [Google Scholar] [CrossRef]
- Luis, O.J.; Ponte, A.C. Control of reproduction of the shrimp Penaeus kerathurus held in captivity. J. World Aquac. Soc. 1993, 24, 31–39. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Mosconi, G.; Salvatori, R.; Lanari, D.; Tomassoni, D.; Carnevali, O.; Polzonetti-Magni, A.M. Effect of dietary supplements of mussel and polychaetes on spawning performance of captive sole, Solea solea (Linnaeus, 1758). Animal Reprod. Sci. 2009, 113, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Durou, C.; Poirier, L.; Amiard, J.C.; Budzinski, H.; Gnassia-Barelli, M.; Lemenach, K.; Peluhet, L.; Mouneyrac, C.; Roméo, M.; Amiard-Triquet, C. Biomonitoring in a clean and a multi-contaminated estuary based on biomarkers and chemical analyses in the endobenthic worm Nereis diversicolor. Environ. Poll. 2007, 148, 445–458. [Google Scholar] [CrossRef]
- Barrick, A.; Marion, J.M.; Perrein-Ettajani, H.; Châtel, A.; Mouneyrac, C. Baseline levels of biochemical biomarkers in the endobenthic ragworm Hediste diversicolor as useful tools in biological monitoring of estuaries under anthropogenic pressure. Mar. Poll. Bull. 2018, 129, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Pombo, A.; Baptista, T.; Granada, L.; Ferreira, S.M.F.; Gonçalves, S.C.; Anjos, C.; Erica, S.A.; Chainho, P.; Cancela da Fonseca, L.; Fidalgo e Costa, P.; et al. Insight into aquaculture’s potential of marine annelid worms and ecological concerns: A review. Rev. Aquac. 2020, 12, 107–121. [Google Scholar] [CrossRef]
- Olive, P.J.W. Polychaete aquaculture and polychaete science: A mutual synergism. Hydrobiologia 1999, 402, 175–183. [Google Scholar] [CrossRef]
- Mayer, P.; Estruch, V.; Blasco, J.; Jover, M. Predicting the growth of gilthead sea bream (Sparus aurata L.) farmed in marine cages under real production conditions using temperature- and time-dependent models. Aquac. Res. 2008, 39, 1046–1052. [Google Scholar] [CrossRef]
- Mayer, P.; Estruch, V.; Martí, P.; Jover, M. Use of quantile regression and discriminant analysis to describe growth patterns in farmed gilthead sea bream (Sparus aurata). Aquaculture 2009, 292, 30–36. [Google Scholar] [CrossRef]
- Galasso, H.L.; Richard, M.; Lefebvre, S.; Aliaume, C.; Callier, M.D. Body size and temperature effects on standard metabolic rate for determining metabolic scope for activity of the polychaete Hediste (Nereis) diversicolor. PeerJ 2018, 6, e5675. [Google Scholar] [CrossRef] [Green Version]
- Hernández, J.M.; Gasca-Leyva, E.; León, C.J.; Vergara, J.M. A growth model for gilthead seabream (Sparus aurata). Ecol. Model. 2003, 165, 265–283. [Google Scholar] [CrossRef]
- Bagarrão, R.; Fidalgo e Costa, P.; Baptista, T.; Pombo, A. Reproduction and growth of polychaete Hediste diversicolor (OF Müller, 1776) under different environmental conditions. In Proceedings of the International Meeting on Marine Research 2014, Peniche, Portugal, 10–11 July 2014. [Google Scholar] [CrossRef]
- Nesto, N.; Simonini, R.; Prevedelli, D.; da Ros, L. Effects of diet and density on growth, survival and gametogenesis of Hediste diversicolor (O.F. Müller, 1776) (Nereididae, Polychaeta). Aquaculture 2012, 362–363, 1–9. [Google Scholar] [CrossRef]
- Fidalgo e Costa, P. Reproduction and growth in captivity of the polychaete Nereis diversicolor O. F. Muller, 1776, using two different kinds of sediment: Preliminary assays. Bol. Inst. Esp. Oceanog. 1999, 15, 351–355. [Google Scholar]
- Nielsen, A.M.; Eriksen, N.T.; Lonsmann Iversen, J.J.; Riisgard, H.U. Feeding, growth and respiration in the polychaetes Nereis diversicolor (facultative filter-feeder) and N. virens (omnivorous)—A comparative study. Mar. Ecol. Prog. Ser. 1995, 125, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Rasines, I.; Martín, I.; Aguado-Giménez, F. Contribution to the intensive cultivation of Hediste diversicolor. In Proceedings of the Aquaculture Europe International Conference and Exposition, Funchal, Portugal, 4–7 October 2021. [Google Scholar]
- García-García, B.; Cerezo-Valverde, J.; Aguado-Giménez, F.; García-García, J.; Hernández, M.D. Effect of the interaction between body weight and temperature on growth and maximum daily food intake in sharpsnout sea bream (Diplodus puntazzo). Aquacult. Int. 2010, 19, 131–141. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology. Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Aguado-Giménez, F.; García-García, B. Growth and food intake models in Octopus vulgaris Cuvier (1797): Influence of body weight, temperature, sex and diet. Aquacult. Int. 2002, 10, 361–377. [Google Scholar] [CrossRef]
- Miliou, H.; Fintikaki, M.; Kountouris, T.; Verriopoulos, G. Combined effects of temperature and body weight on growth and protein utilization of the common octopus, Octopus vulgaris. Aquaculture 2005, 249, 245–256. [Google Scholar] [CrossRef]
- Lin, X.; Xie, S.; Su, Y.; Cui, Y. Optimum temperature for the growth performance of juvenile orange spotted grouper (Epinephelus coioides H.). Chin. J. Oceanol. Limnol. 2008, 26, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Ivleva, I.V. The influence of temperature on the transformation of matter in marine invertebrates. In Marine Food Chains; Steele, J.H., Ed.; Oliver and Boyd: Edinburgh, UK, 1970; pp. 96–112. [Google Scholar]
- Arias, A.M.; Drake, P. Distribution and production of the polychaete Nereis diversicolor in a shallow coastal lagoon in the Bay of Cádiz. Cahiers Biol. Mar. 1995, 36, 201–210. [Google Scholar]
- Ozoh, P.T.E. The effect of temperature and salinity on copper body-burden and copper toxicity to Hediste (Nereis) diversicolor. Environ. Monit. Assess. 1992, 21, 11–17. [Google Scholar] [CrossRef]
- Batista, F.M.; Fidalgo e Costa, P.; Ramos, A.; Passos, A.M.; Pousao Ferreira, P.; Cancela da Foseca, L. Production of the ragworm Nereis diversicolor (O.F. Müller, 1776), fed with a diet for gilthead seabream Sparus aurata L.; 1758: Survival, growth, feed utilization and oogenesis. Bol. Inst. Esp. Oceanog. 2003, 19, 447–451. [Google Scholar]
- Fernandes, F.; Jerónimo, D.; Diniz, M.; Calado, R.; Madeira, D. Cellular stress response and thermal tolerance limits of the ragworm Hediste diversicolor under global change scenarios. In Proceedings of the ICYMARE—International Conference for Young Marine Researchers, Bremen, Germany, 24–27 September 2019. [Google Scholar] [CrossRef]
- Machado, D.; França, M.; Pombo, A.; Anjos, C.M.; Ferreira, S.M.; Gonçalves, S.C.; Costa, P.F.; Costa, J.V.; Baptista, T.M. Effect of different diets on growth of Hediste diversicolor (O. F. Müller, 1776) (Nereididae, Polychaeta) juveniles. In Proceedings of the International Meeting on Marine Research 2016, Peniche, Portugal, 14–15 July 2016. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Rey, F.; Koga Li, H.; Domingues, M.R.M.; Calado, R. Optimizing the timeframe to produce polychaetes (Hediste diversicolor) enriched with esssential fatty acids under different combinations of temperatura and salinity. Front. Mar. Sci. 2021, 8, 671545. [Google Scholar] [CrossRef]
- Björnsson, B.; Seteinarsson, A.; Árnason, T. Growth model for Atlantic cod (Gadus morhua): Effects of temperature and body weight on growth rate. Aquaculture 2007, 271, 216–226. [Google Scholar] [CrossRef]
- Dumas, A.; France, J.; Bureau, D. Modelling growth and body composition in fish: Where have we been and where are we going? Aquac. Res. 2010, 41, 161–181. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dependent Variable | Bw | T | ||
|---|---|---|---|---|
| r | p-Value | r | p-Value | |
| M | 0.1883 | ** | 0.3847 | ** |
| All data | ||||
| AGR | 0.0079 | n.s. | 0.0469 | n.s. |
| SGR | 0.8006 | *** | 0.2422 | ** |
| Bw < 400 mg | ||||
| AGR | 0.8420 | *** | 0.2761 | ** |
| SGR | 0.4352 | *** | 0.2145 | *** |
| Bw > 400 mg | ||||
| AGR | 0.3976 | ** | 0.0289 | n.s. |
| SGR | 0.514 | *** | 0.0302 | n.s. |
| AGR | Intercept a S.E. p-Level | Bw b S.E. p-Level | T c S.E. p-Level | T2 d S.E. p-Level | T Bw e S.E. p-Level | R.S.E. R2adj | ANOVA F p-Level |
|---|---|---|---|---|---|---|---|
| Equation (1) | −3.1213 0.3310 *** | 0.7709 0.0584 *** | 0.1396 0.260 *** | −0.0024 0.0005 *** | −0.0042 0.0033 n.s. | 0.1759 90.3% | 564.1 *** |
| Equation (2) | −2.7767 0.1886 *** | 0.6995 0.0154 *** | 0.1197 0.0208 *** | −0.0024 0.0005 *** | — | 0.1761 90.3% | 749.7 *** |
| Equation (3) | 1.3009 0.3736 *** | −0.2181 0.0659 ** | 0.1739 0.0294 *** | −0.0028 0.0006 *** | −0.0072 0.0037 n.s. | 0.1986 70.4% | 144.8 *** |
| Equation (4) | 1.8927 0.2138 *** | −0.3406 0.0175 *** | 0.1398 0.0236 *** | −0.0029 0.0006 *** | — | 0.1997 70.4% | 749.7 *** |
| Equation (1): Ln(AGR) = Ln(a) + b·Ln(Bw) + c·T + d·T2 + e·T·Ln(Bw) Equation (2): Ln(AGR) = Ln(a) + b·Ln(Bw) + c·T + d·T2 Equation (3): Ln(SGR) = Ln(a) + b·Ln(Bw)+c·T + d·T2 + e·T·Ln(Bw) Equation (4): Ln(SGR) = Ln(a) + b·Ln(Bw) + c·T + d·T2 | |||||||
| Intercept a S.E. p-Level | Bw b S.E. p-Level | R.S.E. R2 | ANOVA F p-Level | |
|---|---|---|---|---|
| Equation (5) | 3.7852 0.0652 *** | −0.1335 0.1010 *** | 0.4458 54.6% | 177.7 *** |
| Equation (6) | 6.3971 0.4425 *** | −0.7977 0.0684 *** | 0.3027 48.4% | 138.8 *** |
| Equation (5): Ln(AGR + 17) = Ln(a) + b·Ln(Bw) Equation (6): Ln(SGR + 2) = Ln(a) + b·Ln(Bw) | ||||
| Intercept a S.E. p-Level | Bw b S.E. p-Level | T c S.E. p-Level | T2 d S.E. p-Level | T Bw e S.E. p-Level | R.S.E. R2adj | ANOVA F p-Level | |
|---|---|---|---|---|---|---|---|
| Equation (7) | 2.3010 0.3675 *** | −0.1937 0.0547 *** | 0.1206 0.0308 *** | −0.0006 0.0007 n.s. | −0.0144 0.0031 *** | 0.2639 72.9% | 260.6 *** |
| Equation (8) | 2.4899 0.3057 *** | −0.1933 0.0547 *** | −0.09716 0.0175 *** | — | −0.0144 0.0031 *** | 0.2639 72.9% | 347.3 *** |
| Equation (9) | 1.7827 0.0526 *** | −0.0008 0.0001 *** | 0.0757 0.0075 *** | — | −0.0109 0.0012 *** | 0.2221 80.8% | 542.6 *** |
| Equation (7): Ln(SGR + 2) = Ln(a) + b·Ln(Bw) + c·T + d·T2 + e·T·Ln(Bw) Equation (8): Ln(SGR + 2) = Ln(a) + b·Ln(Bw) + c·T + e·T·Ln(Bw) Equation (9): Ln(SGR + 2) = Ln(a) + b·Bw + c·T + e·T·Ln(Bw) | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguado-Giménez, F.; García-García, B.; Martín, I.E.; Rasines, I. Influence of Body Weight and Water Temperature on Growth in Ragworm Hediste diversicolor. Aquac. J. 2023, 3, 19-31. https://doi.org/10.3390/aquacj3010004
Aguado-Giménez F, García-García B, Martín IE, Rasines I. Influence of Body Weight and Water Temperature on Growth in Ragworm Hediste diversicolor. Aquaculture Journal. 2023; 3(1):19-31. https://doi.org/10.3390/aquacj3010004
Chicago/Turabian StyleAguado-Giménez, Felipe, Benjamin García-García, Ignacio Eduardo Martín, and Inmaculada Rasines. 2023. "Influence of Body Weight and Water Temperature on Growth in Ragworm Hediste diversicolor" Aquaculture Journal 3, no. 1: 19-31. https://doi.org/10.3390/aquacj3010004
APA StyleAguado-Giménez, F., García-García, B., Martín, I. E., & Rasines, I. (2023). Influence of Body Weight and Water Temperature on Growth in Ragworm Hediste diversicolor. Aquaculture Journal, 3(1), 19-31. https://doi.org/10.3390/aquacj3010004