
Modulating Natural Methane Release from Rumen Fermentation through the Use of Ficus glomerata Leaf Tannins in Murrah Buffalo (Bubalus bubalis)

Abstract
:1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Experimental Design and Substrate
3.2. Collection of Rumen Inoculum
3.3. In Vitro Incubation
3.4. Estimation of Fatty Acid Biohydrogenation
3.5. Estimation of Gas and Methane Production
3.6. Volatile Fatty Acid (VFA) Estimation
3.7. In Vitro Dry Matter Degradability and Ammonia Production
3.8. Chemical and Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Getabalew, M.; Alemneh, T.; Akeberegn, D. Methane production in ruminant animals: Implication for their impact on climate change. Concepts Dairy Vet. Sci 2019, 2, 204–211. [Google Scholar]
- Pearce, W.; Holmberg, K.; Hellsten, I.; Nerlich, B. Climate change on Twitter: Topics, communities and conversations about the 2013 IPCC Working Group 1 report. PLoS ONE 2014, 9, e94785. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, M.P. Environmental performance and vulnerability to climate change: A case study of India, Nepal, Bangladesh and Pakistan. In Climate Change and Disaster Risk Management; Filho, W.L., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 149–167. [Google Scholar]
- Cottle, D.J.; Nolan, J.V.; Wiedemann, S.G. Ruminant enteric methane mitigation: A review. Anim. Prod. Sci. 2011, 51, 491–514. [Google Scholar] [CrossRef]
- Nawab, A.; Li, G.; An, L.; Nawab, Y.; Zhao, Y.; Xiao, M.; Tang, S.; Sun, C. The potential effect of dietary tannins on enteric methane emission and ruminant production, as an alternative to antibiotic feed additives—A review. Ann. Anim. Sci. 2020, 20, 355–388. [Google Scholar] [CrossRef]
- Schofield, P.; Mbugua, D.; Pell, A. Analysis of condensed tannins: A review. Anim. Feed. Sci. Technol. 2001, 91, 21–40. [Google Scholar] [CrossRef]
- Waghorn, G. Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production—Progress and challenges. Anim. Feed. Sci. Technol. 2008, 147, 116–139. [Google Scholar] [CrossRef]
- Naumann, H.; Sepela, R.; Rezaire, A.; Masih, S.E.; Zeller, W.E.; Reinhardt, L.A.; Robe, J.T.; Sullivan, M.L.; Hagerman, A.E. Relationships between structures of condensed tannins from Texas legumes and methane production during in vitro rumen digestion. Molecules 2018, 23, 2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef]
- Dey, A.; Dutta, N.; Sharma, K.; Pattanaik, A. Evaluation of condensed tannins from tropical tree leaves and its impact on in vitro nitrogen degradability of groundnut cake. Anim. Nutr. Feed. Technol. 2006, 6, 215–222. [Google Scholar]
- Ashwin, K.; Pattanaik, A.K.; Howarth, G.S. Polyphenolic bioactives as an emerging group of nutraceuticals for promotion of gut health: A review. Food Biosci. 2021, 44, 101376. [Google Scholar] [CrossRef]
- Ranjhan, S. Dairy feeding systems. In Smallholder Dairying in the Tropics; ILRI (International Livestock Research Institute): Nairobi, Kenya, 1999; pp. 117–132. [Google Scholar]
- Bhatta, R.; Saravanan, M.; Baruah, L.; Prasad, C. Effects of graded levels of tannin-containing tropical tree leaves on in vitro rumen fermentation, total protozoa and methane production. J. Appl. Microbiol. 2015, 118, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.; McGinn, S.; Martinez, T.; McAllister, T. Use of condensed tannin extract from quebracho trees to reduce methane emissions from cattle. J. Anim. Sci. 2007, 85, 1990–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, B.; Pinchak, W.; Anderson, R.; Fulford, J.; Puchala, R. Effects of condensed tannins supplementation level on weight gain and in vitro and in vivo bloat precursors in steers grazing winter wheat. J. Anim. Sci. 2006, 84, 2546–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Barry, T.N.; Manley, T.R. Interrelationships between the concentrations of total condensed tannin, free condensed tannin and lignin in Lotus sp. and their possible consequences in ruminant nutrition. J. Sci. Food Agric. 1986, 37, 248–254. [Google Scholar] [CrossRef]
- Frutos, P.; Hervas, G.; Giráldez, F.J.; Mantecón, A. Tannins and ruminant nutrition. Span. J. Agric. Res. 2004, 2, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Bhatta, R.; Uyeno, Y.; Tajima, K.; Takenaka, A.; Yabumoto, Y.; Nonaka, I.; Enishi, O.; Kurihara, M. Difference in the nature of tannins on in vitro ruminal methane and volatile fatty acid production and on methanogenic archaea and protozoal populations. J. Dairy Sci. 2009, 92, 5512–5522. [Google Scholar] [CrossRef] [Green Version]
- Toral, P.G.; Monahan, F.; Hervás, G.; Frutos, P.; Moloney, A. Modulating ruminal lipid metabolism to improve the fatty acid composition of meat and milk. Challenges and opportunities. Animal 2018, 12, s272–s281. [Google Scholar] [CrossRef] [Green Version]
- Hassan, F.-u.; Arshad, M.A.; Ebeid, H.M.; Rehman, M.S.-u.; Khan, M.S.; Shahid, S.; Yang, C. Phytogenic additives can modulate rumen microbiome to mediate fermentation kinetics and methanogenesis through exploiting diet–microbe interaction. Front. Vet. Sci. 2020, 7, 575801. [Google Scholar] [CrossRef]
- Carreño, D.; Hervás, G.; Toral, P.G.; Belenguer, A.; Frutos, P. Ability of different types and doses of tannin extracts to modulate in vitro ruminal biohydrogenation in sheep. Anim. Feed. Sci. Technol. 2015, 202, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Guerreiro, O.; Alves, S.P.; Costa, M.; Duarte, M.F.; Jerónimo, E.; Bessa, R.J.B. Effects of Increasing Doses of Condensed Tannins Extract from Cistus Ladanifer L. on In Vitro Ruminal Fermentation and Biohydrogenation. Animals 2021, 11, 761. [Google Scholar] [CrossRef]
- Makkar, H.P. Quantification of Tannins in Tree and Shrub Foliage: A Laboratory Manual; Springer Science & Business Media: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Menke, K.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Mandal, G.; Roy, A.; Patra, A. Effects of feeding plant additives rich in saponins and essential oils on the performance, carcass traits and conjugated linoleic acid concentrations in muscle and adipose tissues of Black Bengal goats. Anim. Feed. Sci. Technol. 2014, 197, 76–84. [Google Scholar] [CrossRef]
- Conway, E. Ammonia. General Method. In Microdiffusion Analysis and Volumetric Error; Crosby Lockwood and Son Ltd.: London, UK, 1962; pp. 98–100. [Google Scholar]
- Association of Official Analytical Chemistry (AOAC). Official Methods of Analysis, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- SPSS. Statistical Packages for Social Sciences, version 17.0; SPSS Inc.: Chicago, IL, USA, 2008.
- Snedecor, G.; Cochran, W. Statistical Methods, 8th ed.; East West Press Pvt. Ltd.: New Delhi, India, 1994; p. 313. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Attributes | Oat Hay | Concentrate Mixture 1 | Ficus glomerata Leaf Meal |
|---|---|---|---|
| Chemical composition | |||
| Organic matter | 93.30 ± 1.12 | 91.07 ± 1.01 | 88.50 ± 1.23 |
| Crude protein | 7.80 ± 0.56 | 19.50 ± 0.67 | 14.85 ± 0.35 |
| Ether extract | 3.07 ± 0.60 | 4.12 ± 0.45 | 4.54 ± 0.24 |
| Total ash | 6.70 ± 0.72 | 8.93 ± 0.63 | 11.50 ± 0.82 |
| Neutral detergent fiber | 58.40 ± 1.35 | 44.60 ± 1.23 | 38.70 ± 1.67 |
| Acid detergent fiber | 43.70 ± 1.73 | 21.06 ± 0.96 | 31.52 ± 1.11 |
| Phenolic fractions | |||
| Total phenolics 2 | - | - | 16.92 ± 0.52 |
| Total tannin phenolics 2 | 15.71 ± 0.41 | ||
| Non-tannin phenolics 2 | 1.22 ± 0.06 | ||
| Condensed tannins 3 | 10.64 ± 0.33 |
| Attributes | Control | FG-0.25 | FG-0.50 | FG-1.0 | SEM | p Value |
|---|---|---|---|---|---|---|
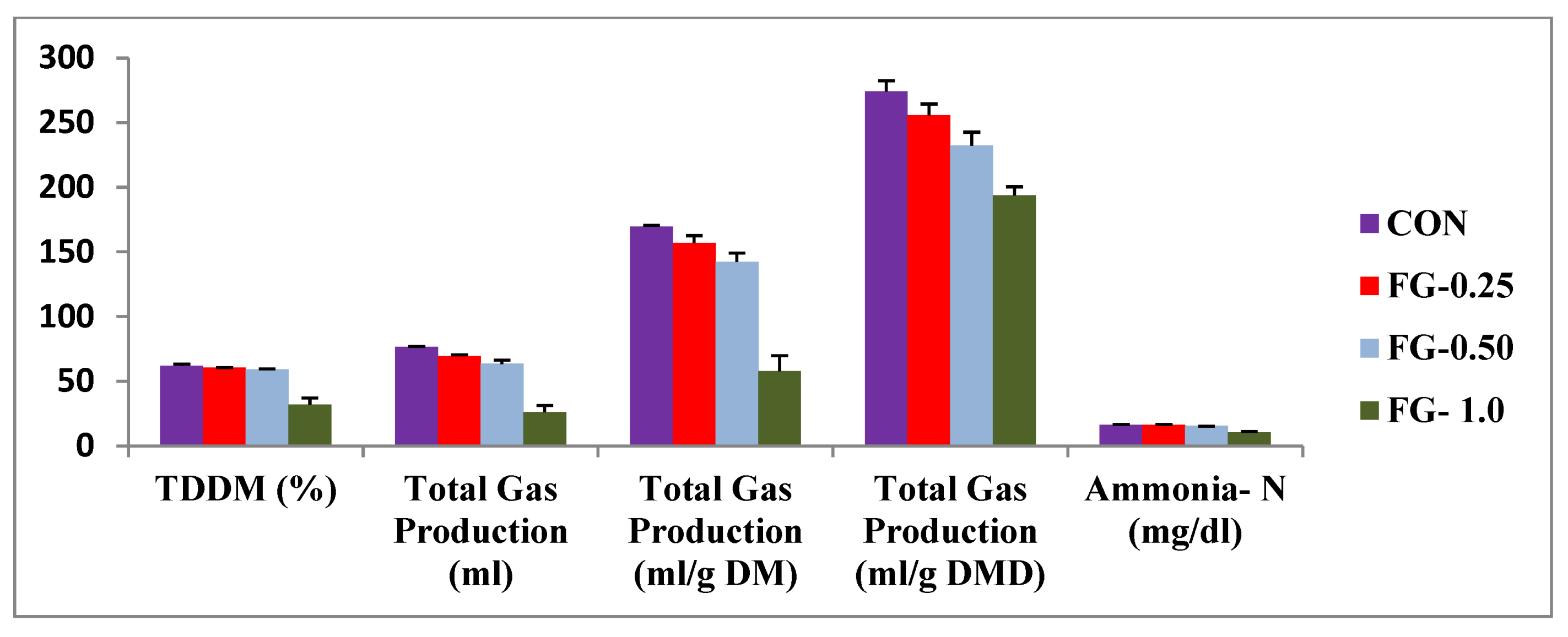
| Total gas production (mL) | 76.55 c ± 0.47 | 69.32 bc ± 1.32 | 63.48 b ± 3.06 | 26.16 a ± 5.27 | 6.85 | <0.001 |
| Total gas production (mL/g DM) | 169.52 c ± 1.14 | 157.07 bc ± 5.57 | 142.36 b ± 6.85 | 57.95 a ± 11.83 | 15.35 | <0.001 |
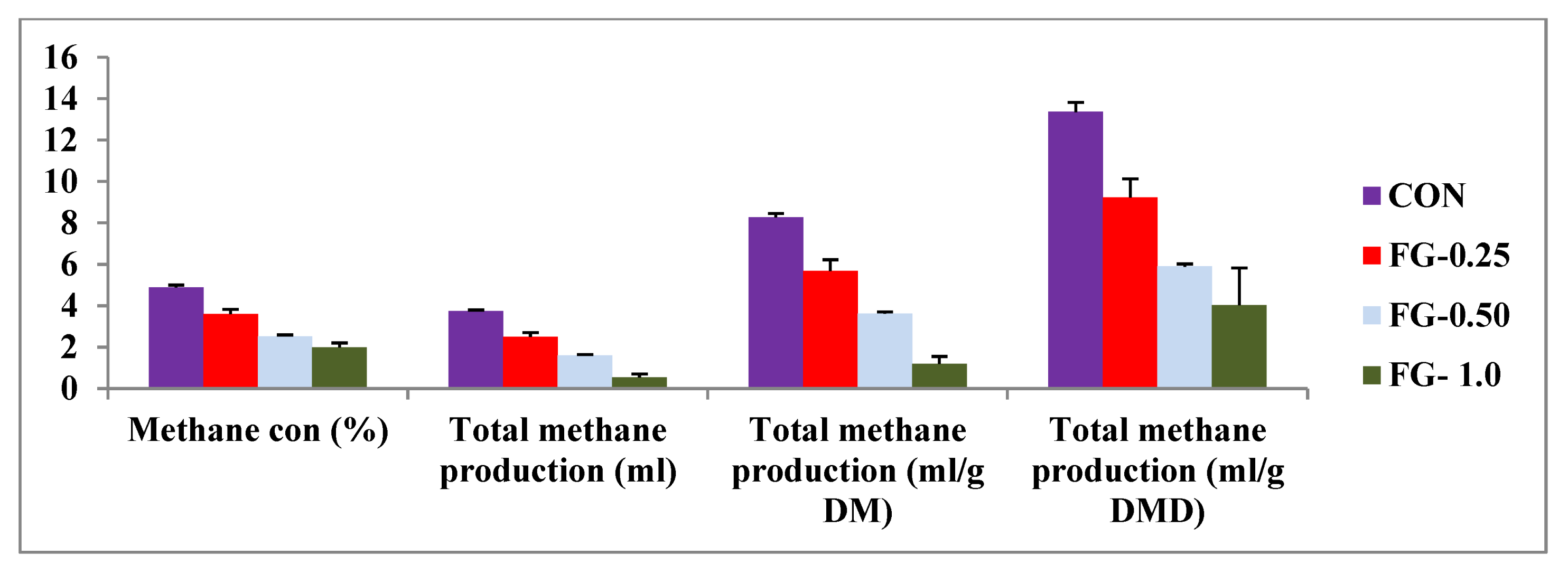
| Methane concentration (%) | 4.88 c ± 0.12 | 3.60 b ± 0.23 | 2.53 a ± 0.06 | 2.0 a ± 0.21 | 0.41 | <0.001 |
| Total methane production (mL) | 3.73 d ± 0.07 | 2.50 c ± 0.20 | 1.60 b ± 0.04 | 0.54 a ± 0.16 | 0.43 | <0.001 |
| Total methane production (mL/g DM) | 8.27 d ± 0.18 | 5.67 c ± 0.56 | 3.61 b ± 0.09 | 1.19 a ± 0.36 | 0.96 | <0.001 |
| Total methane production (mL/g DMD) | 13.36 c ± 0.46 | 9.22 b ± 0.91 | 5.89 ab ± 0.13 | 4.03 a ± 1.80 | 1.37 | 0.003 |
| TDDM (%) | 61.96 c ± 1.46 | 60.45 c ± 0.03 | 56.28 b ± 0.21 | 31.82 a ± 5.32 | 4.49 | 0.001 |
| Ammonia N (mg/dL) | 16.33 c ± 0.47 | 16.10 b ± 0.70 | 15.40 b ± 0.0 | 10.50 a ± 0.70 | 0.84 | 0.002 |
| Acetate (mM/dL) | 3.46 c ± 0.08 | 3.40 c ± 0.07 | 2.96 b ± 0.02 | 2.19 a ± 0.07 | 0.10 | 0.002 |
| Propionate (mM/dL) | 0.88 b ± 0.02 | 1.02 c ± 0.04 | 0.84 b ± 0.06 | 0.57 a ± 0.07 | 0.04 | 0.002 |
| Butyrate (mM/dL) | 0.25 b ± 0.01 | 0.23 b ± 0.02 | 0.24 b ± 0.01 | 0.15 a ± 0.01 | 0.02 | 0.001 |
| A:P ratio | 3.93 c ± 0.02 | 3.33 a ± 0.01 | 3.52 b ± 0.05 | 3.84 c ± 0.04 | 1.02 | 0.001 |
| Fatty Acids | Control | FG-0.25 | FG-0.50 | FG-1.0 | SEM | p Value |
|---|---|---|---|---|---|---|
| Saturated fatty acids (SFA) | ||||||
| C4:0 | 36.75 bc ± 0.22 | 37.11 c ± 0.05 | 36.29 b ± 0.19 | 0.0 a ± 0.0 | 4.79 | <0.001 |
| C6:0 | 3.92 c ± 0.09 | 3.34 b ± 0.06 | 3.42 b ± 0.04 | 0.0 a ± 0.0 | 0.47 | <0.001 |
| C8:0 | 0.0 a ± 0.0 | 3.55 b ± 1.78 | 9.30 c ± 0.46 | 2.72 ab ± 0.18 | 1.09 | 0.001 |
| C10:0 | 3.82 b ± 0.09 | 4.04 c ± 0.08 | 4.20 c ± 0.07 | 0.0 a ± 0.0 | 0.53 | <0.001 |
| C11:0 | 4.48 c ± 0.09 | 4.71 c ± 0.12 | 4.00 b ± 0.07 | 0.0 a ± 0.0 | 0.58 | <0.001 |
| C12:0 | 5.74 a ± 0.09 | 5.64 a ± 0.07 | 5.22 a ± 0.01 | 10.95 b ± 0.49 | 0.72 | <0.001 |
| C13:0 | 2.13 a ± 0.09 | 2.31 a ± 0.23 | 2.78 a ± 0.09 | 13.17 b ± 0.88 | 1.42 | <0.001 |
| C14:0 | 6.74 a ± 0.04 | 5.15 a ± 0.11 | 5.27 a ± 0.05 | 43.76 b ± 4.8 | 5.07 | <0.001 |
| C15:0 | 2.71 a ± 0.10 | 2.13 a ± 1.4 | 0.37 a ± 0.37 | 10.48 b ± 0.80 | 1.27 | <0.001 |
| C16:0 | 16.47 d ± 0.39 | 13.52 c ± 0.09 | 10.29 b ± 0.13 | 0.0 a ± 0.0 | 1.87 | <0.001 |
| C18:0 | 15.27 b ± 0.14 | 5.40 b ± 0.15 | 0.6.32 c ± 0.22 | 2.86 a ± 0.08 | 0.39 | <0.001 |
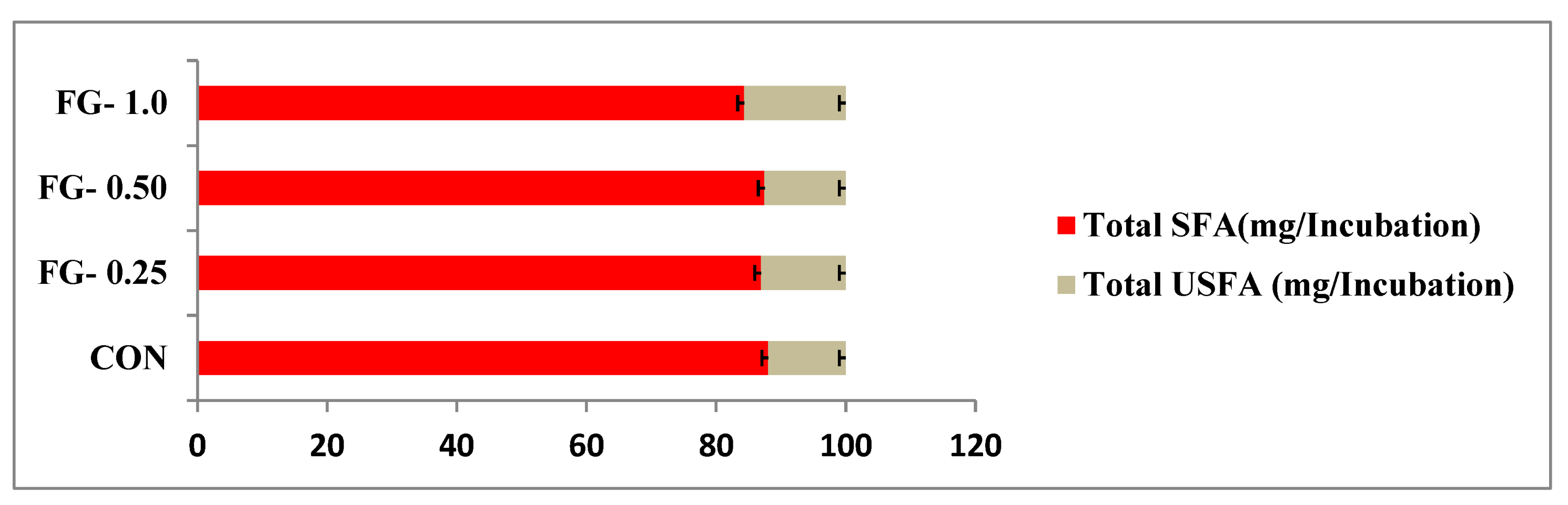
| Total SFA | 88.05 ± 0.29 | 87.93 ± 0.52 | 87.49 ± 0.19 | 84.32 ± 3.64 | 0.89 | 0.53 |
| C14:1 | 11.94 d ± 0.29 | 10.89 c ± 0.19 | 9.05 b ± 0.12 | 7.69 a ± 0.44 | 0.50 | <0.001 |
| Trans-vaccenic acid (C18:1) | 0.0 a ± 0.0 | 2.89 ab ± 0.39 | 3.46 ab ± 0.29 | 7.99 b ± 3.49 | 1.16 | 0.063 |
| Total unsaturated fatty acids | 11.94 ± 0.29 | 13.07 ± 0.52 | 12.51 ± 0.19 | 15.68 ± 3.67 | 0.89 | 0.53 |
| Dose Incubated (mL/60 mL BRF) | SFA Inhibited | t-Vaccenic Acid (C18:1) Enhanced | Gas Production Inhibited | Methane Production Inhibited | TDDM Inhibited |
|---|---|---|---|---|---|
| 0.25 | 0.14 | 100 | 9.44 | 32.98 | 2.01 |
| 0.50 | 0.64 | 100 | 17.07 | 57.10 | 8.73 |
| 1.0 | 4.24 | 100 | 65.83 | 85.52 | 48.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.K.; Dey, A.; Singh, M. Modulating Natural Methane Release from Rumen Fermentation through the Use of Ficus glomerata Leaf Tannins in Murrah Buffalo (Bubalus bubalis). Methane 2023, 2, 319-328. https://doi.org/10.3390/methane2030021
Singh RK, Dey A, Singh M. Modulating Natural Methane Release from Rumen Fermentation through the Use of Ficus glomerata Leaf Tannins in Murrah Buffalo (Bubalus bubalis). Methane. 2023; 2(3):319-328. https://doi.org/10.3390/methane2030021
Chicago/Turabian StyleSingh, Ram Kumar, Avijit Dey, and Mala Singh. 2023. "Modulating Natural Methane Release from Rumen Fermentation through the Use of Ficus glomerata Leaf Tannins in Murrah Buffalo (Bubalus bubalis)" Methane 2, no. 3: 319-328. https://doi.org/10.3390/methane2030021
APA StyleSingh, R. K., Dey, A., & Singh, M. (2023). Modulating Natural Methane Release from Rumen Fermentation through the Use of Ficus glomerata Leaf Tannins in Murrah Buffalo (Bubalus bubalis). Methane, 2(3), 319-328. https://doi.org/10.3390/methane2030021