
Functional Interdependence in Coupled Dissipative Structures: Physical Foundations of Biological Coordination

 , ,
, , 
Abstract
:1. Introduction
1.1. Coordinative Structures
1.2. Self-Organization
1.3. A Simple Self-Organizing System: E-SOFI
1.4. The Present Study
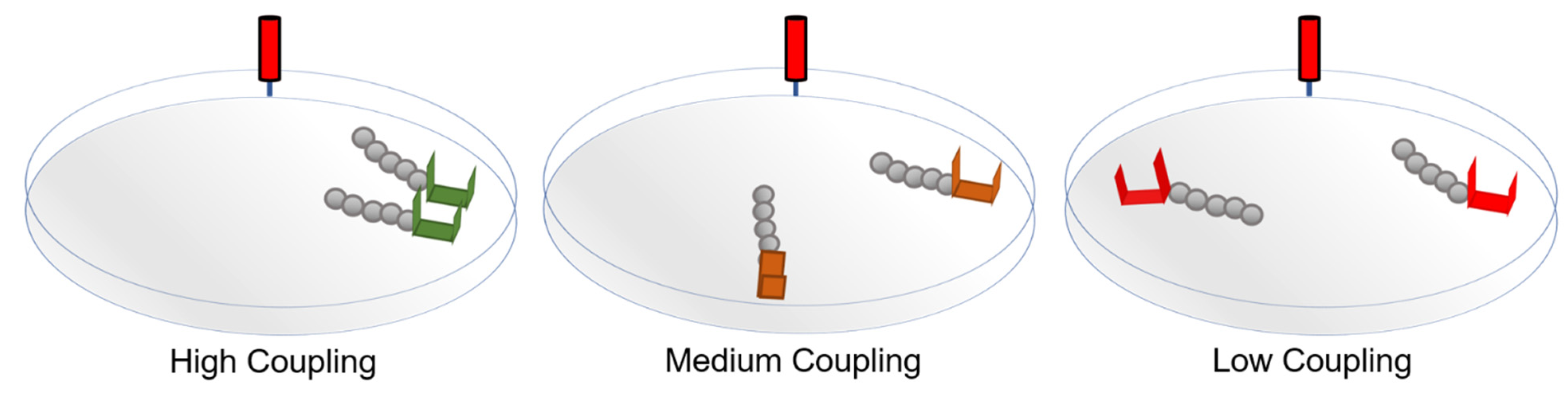
2. Materials and Methods
2.1. Materials and Procedure
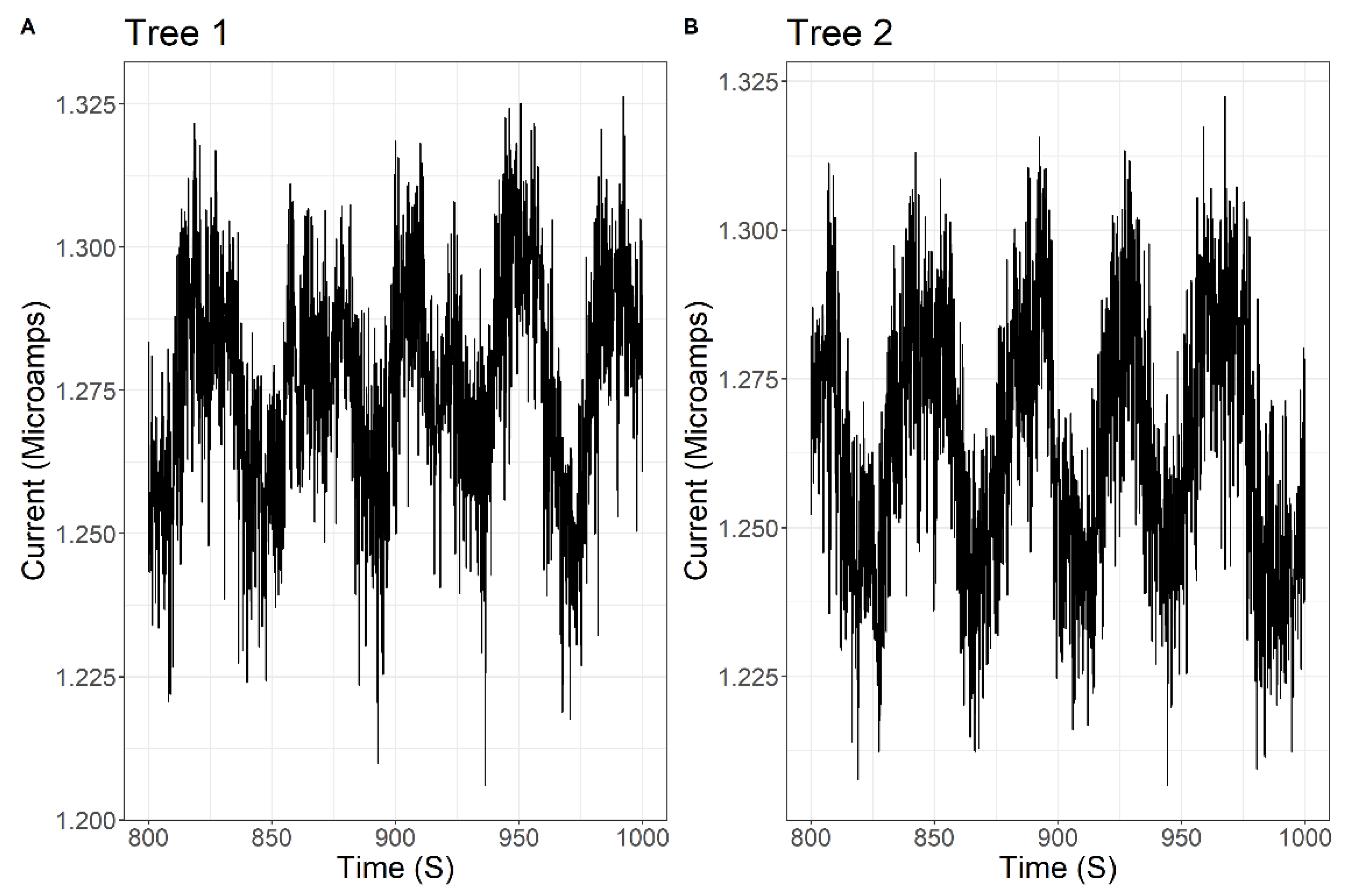
2.2. Data Processing
3. Results
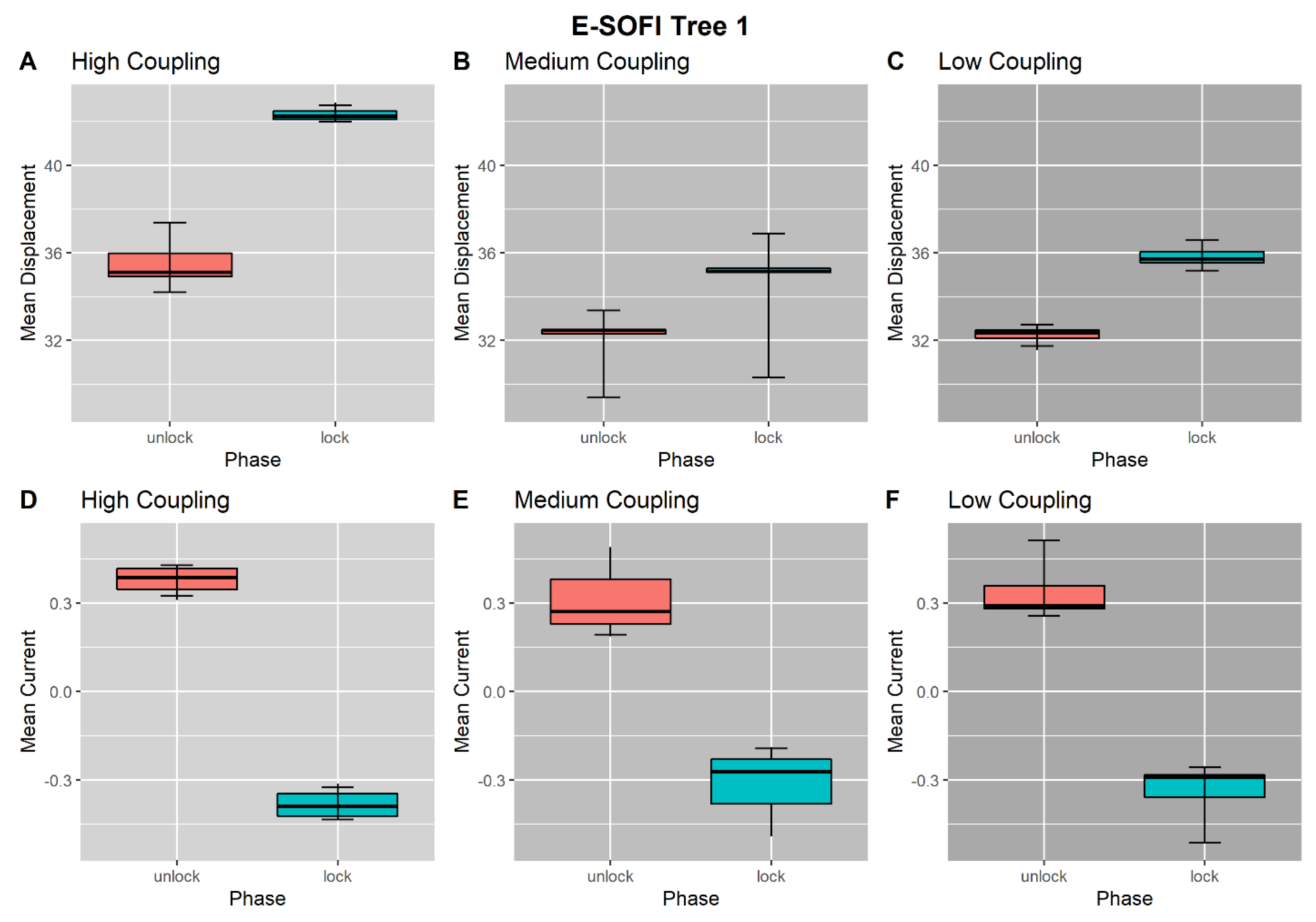
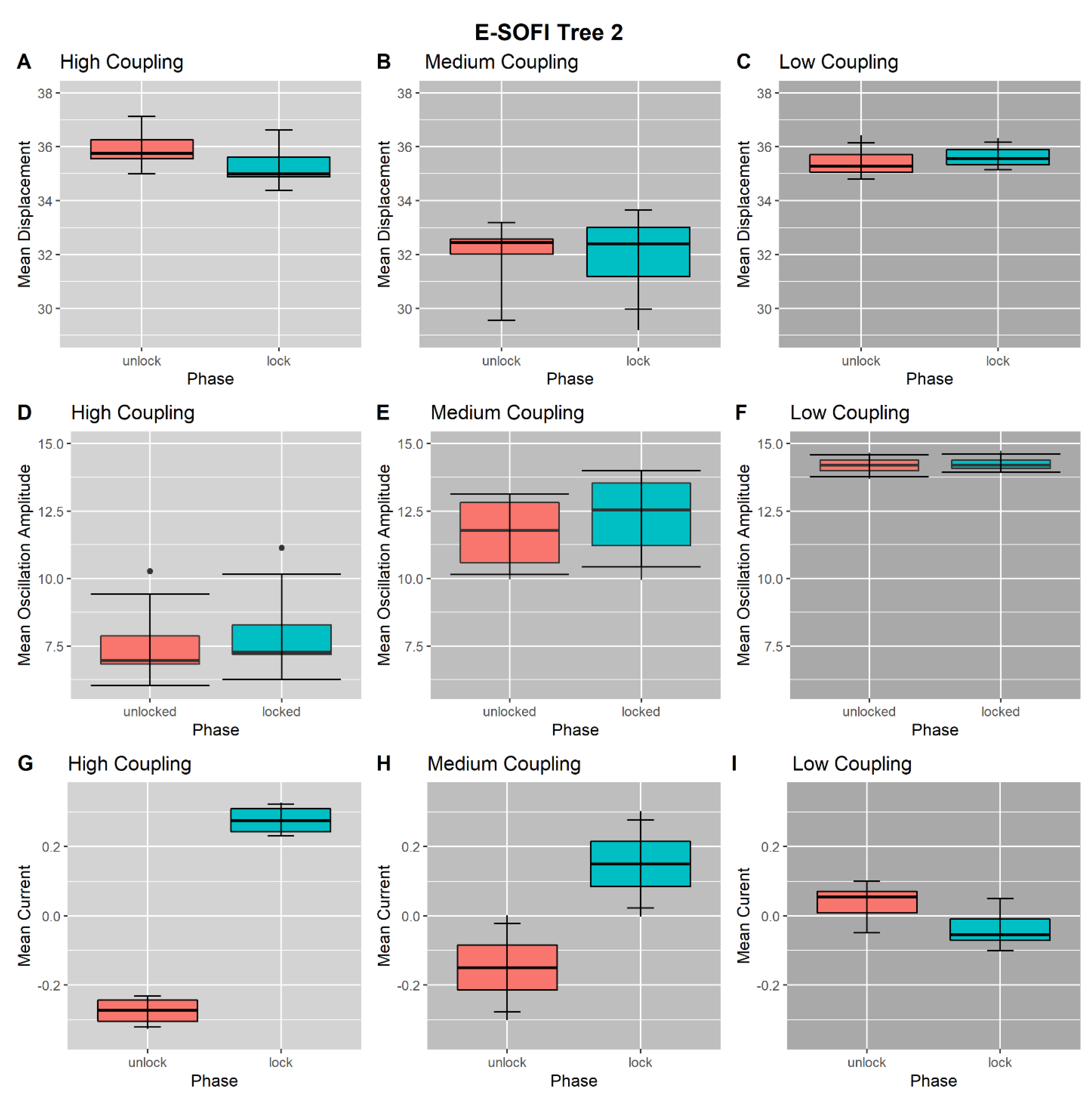
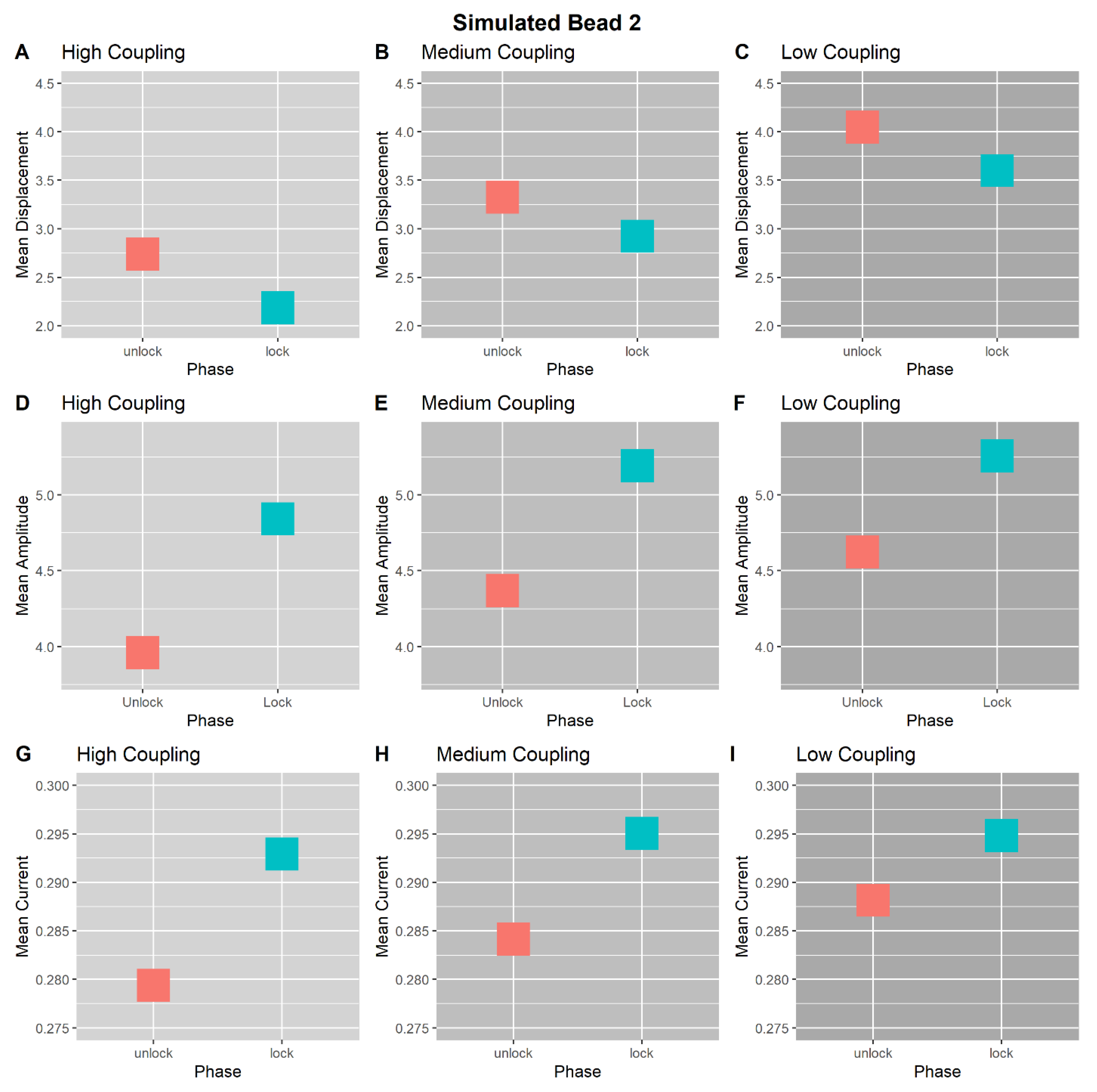
3.1. High-Coupling Condition
3.2. Medium-Coupling Condition
3.3. Low-Coupling Condition
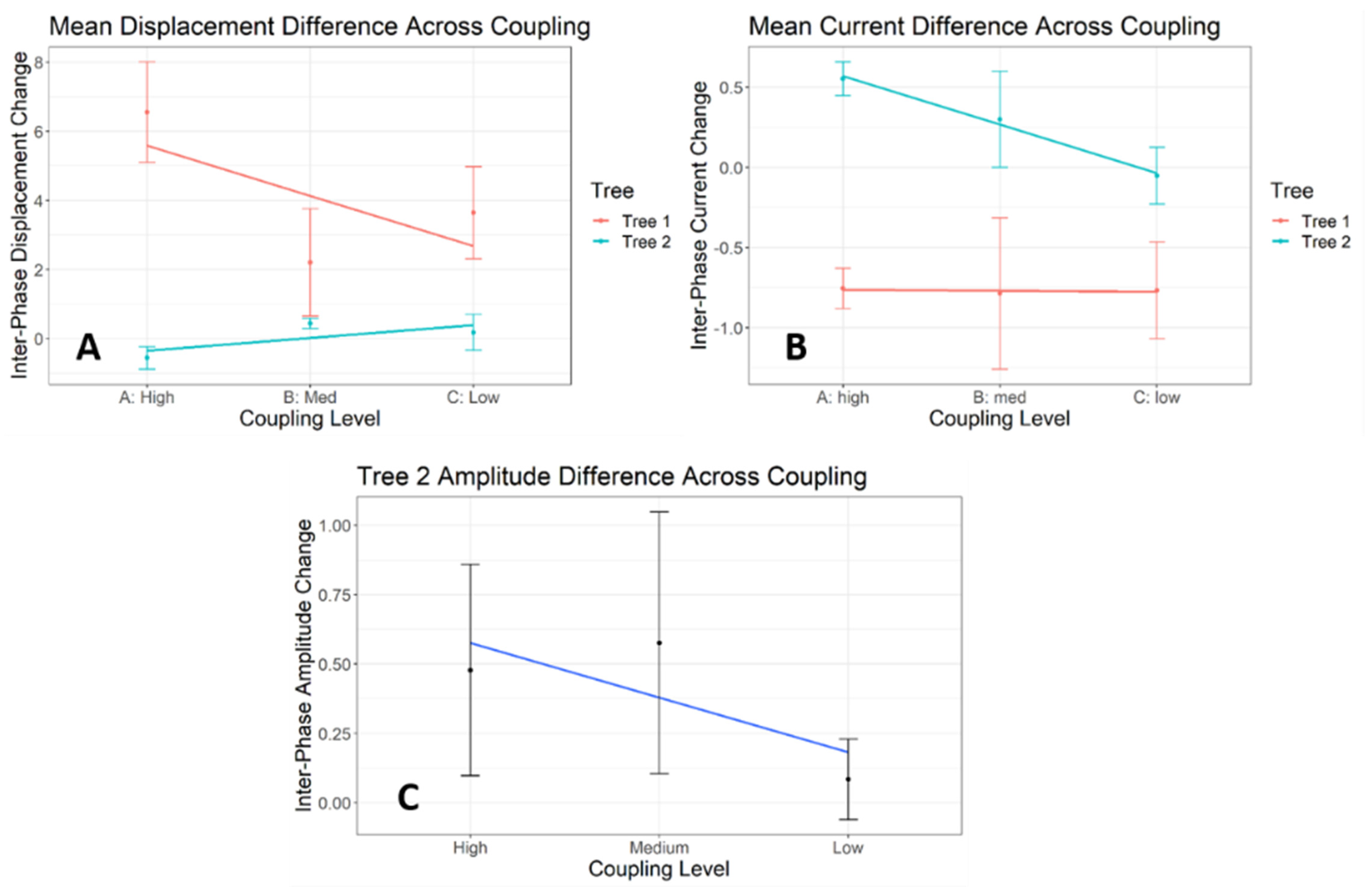
3.4. Coupling-Condition Effects on Inter-Phase Changes
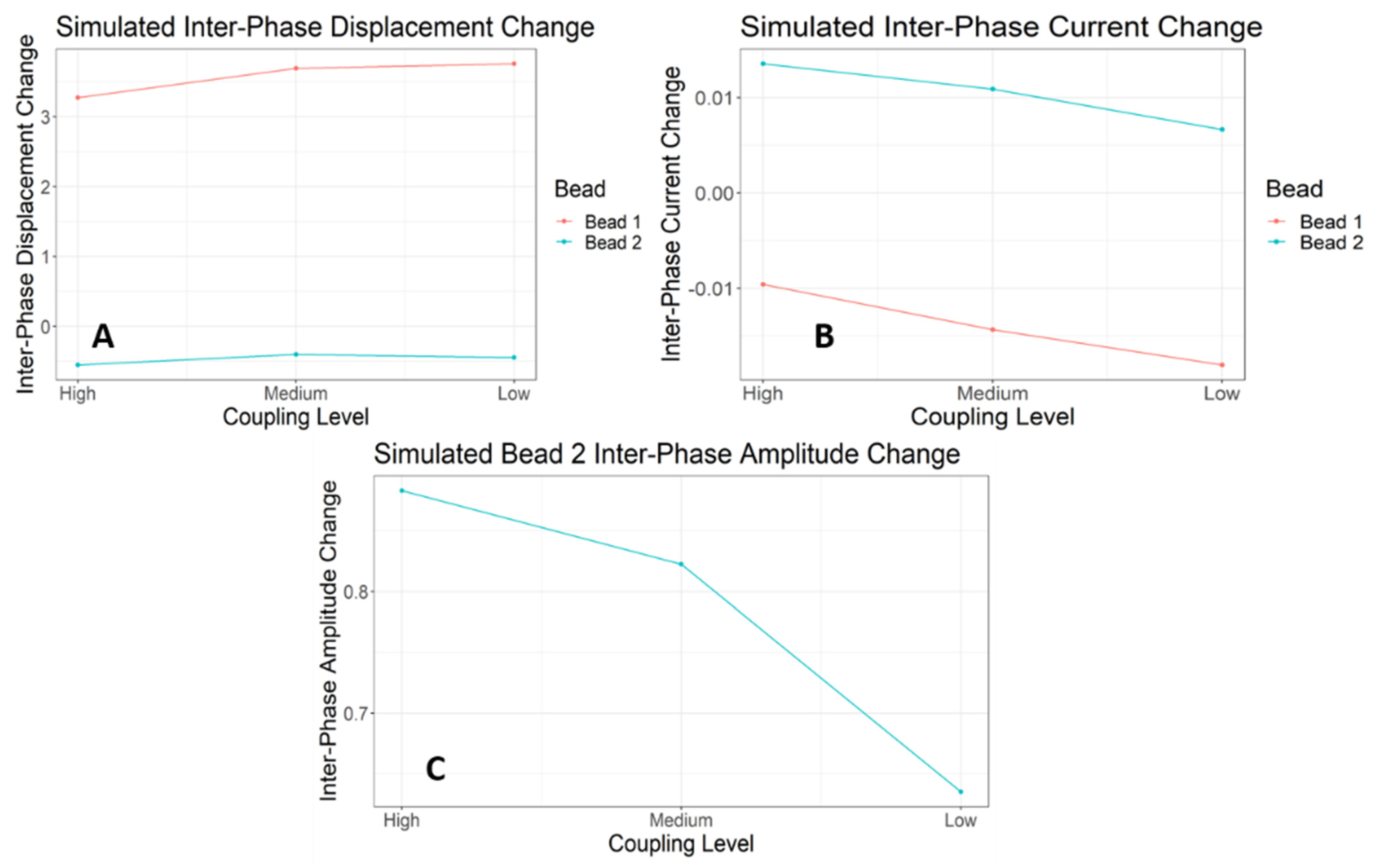
3.5. Simulations of the E-SOFI Dynamics
3.6. Simulation Results
4. Discussion
4.1. Interpreting the Results
4.2. Entailments of a Thermodynamic Account
4.3. Implications for Biological Coordination
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bernstein, N. Coordination and Regulation of Movements; Pergamon Press: Oxford, UK, 1967. [Google Scholar]
- Latash, M.L. The Bernstein problem: How does the central nervous system make its choices? In Dexterity and Its Development; Latash, M.L., Turvey, M.T., Eds.; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 1996; pp. 277–304. [Google Scholar]
- Nakagaki, T.; Yamada, H.; Toth, A. Intelligence: Maze-solving by an amoeboid organism. Nature 2000, 407, 470. [Google Scholar] [CrossRef] [PubMed]
- Seeley, T.D.; Camazine, S.; Sneyd, J. Collective decision-making in honey bees: How colonies choose among nectar sources. Behav. Ecol. Sociobiol. 1991, 28, 277–290. [Google Scholar] [CrossRef]
- Riley, M.A.; Richardson, R.J.; Shockley, K.; Ramenzoni, V.C. Interpersonal synergies. Front. Psychol. 2011, 2, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxton, A.; Dale, R.; Richardson, D.C. Social coordination of verbal and nonverbal behaviours. In Interpersonal Coordination and Performance in Social Systems; Passos, P., Davids, K., Chow, J.Y., Eds.; Routledge: London, UK, 2016; pp. 259–273. [Google Scholar]
- Hove, M.J.; Risen, J.L. It’s all in the timing: Interpersonal synchrony increases affiliation. Soc. Cogn. 2009, 27, 949–960. [Google Scholar] [CrossRef] [Green Version]
- Tunçgenç, B.; Cohen, E. Movement synchrony forges social bonds across group divides. Front. Psychol. 2016, 7, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiltermuth, S.S.; Heath, C. Synchrony and cooperation. Psychol. Sci. 2009, 20, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kelso, J.A.S.; Tuller, B.; Vatikiotis-Bateson, E.; Fowler, C.A. Functionally specific articulatory cooperation following jaw perturbations during speech: Evidence for coordinative structures. J. Exp. Psychol. Hum. Percept. Perform. 1984, 10, 812–832. [Google Scholar] [CrossRef]
- Turvey, M.T. Action and perception at the level of synergies. Hum. Mov. Sci. 2007, 26, 657–697. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, M.H.; Farley, C.T.; Full, R.J.; Koehl, M.A.R.; Kram, R.; Lehman, S. How animals move: An integrative view. Am. Assoc. Adv. Sci. 2000, 288, 100–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turvey, M.T.; Shaw, R.E.; Mace, W.M. Issues in the theory of action: Degrees of freedom, coordinative structures and coalitions. In Attention and Performance VII; Requin, J., Ed.; Lawrence Erlbaum Associates: Hillsdale, NJ, USA, 1978; pp. 557–595. [Google Scholar]
- Li, Z.; Latash, M.L.; Zatsiorsky, V.M. Force sharing among fingers as a model of the redundancy problem. Exp. Brain Res. 1998, 119, 278–286. [Google Scholar] [CrossRef]
- Black, D.P.; Riley, M.A.; McCord, C.K. Synergies in intra- and interpersonal interlimb rhythmic coordination. Hum. Kinet. 2007, 11, 348–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, R.J.; Burger, M. Oscillations and Traveling Waves in Chemical Systems; John Wiley & Sons: New York, NY, USA, 1985. [Google Scholar]
- Nicolis, G. Physics of far-from equilibrium systems and self-organization. In New Physics; Davies, P., Ed.; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Prigogine, I.; Nicolis, G. On symmetry-breaking instabilities in dissipative systems. J. Chem. Phys. 1967, 46, 3542–3550. [Google Scholar] [CrossRef]
- Dixon, J.A.; Kondepudi, D.; Davis, T.J. End-directedness and context in nonliving dissipative structures. In Contextuality from Quantum Physics to Psychology; World Scientific: Singapore, 2016; pp. 185–208. [Google Scholar]
- Kondepudi, D. Self-organization, entropy production, and physical intelligence. Ecol. Psychol. 2012, 24, 33–45. [Google Scholar] [CrossRef]
- Kondepudi, D.; Kay, B.; Dixon, J.A. End-directed evolution and the emergence of energy-seeking behavior in a complex system. Phys. Rev. E 2015, 91, 050902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ting, L.H.; Macpherson, J.M. A limited set of muscle synergies for force control during a postural task. J. Neurophysiol. 2005, 93, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Ramenzoni, V.C. Effects of joint task performance on interpersonal postural coordination. Ph.D. Thesis, University of Cincinnati, Cincinnati, OH, USA, 2008. [Google Scholar]
- Haken, H. The Science of Structure: Synergetics; Deutsche Verlags-Anstalt: Stuttgart, Germany, 1981. [Google Scholar]
- Kaplan, M.L.; Kaplan, N.R. The self-organization of human psychological functioning. Behav. Sci. 1991, 36, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Kondepudi, D.; Prigogine, I. Modern Thermodynamics: From Heat Engines to Dissipative Structures; John Wiley & Sons Ltd.: West Sussex, UK, 1998. [Google Scholar]
- Gallistel, C.R. Coordinate transformations in the genesis of directed action. In Cognitive Science; Bly, B.M., Rumelhart, D.E., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 1–42. [Google Scholar]
- Kugler, P.N.; Kelso, J.A.S.; Turvey, M.T. On the concept of coordinative structures as dissipative structures: I. theoretical lines of convergence. In Tutorials in Motor Behavior; Stelmach, G.E., Requin, J., Eds.; North-Holland Publishing Co.: New York, NY, USA, 1980. [Google Scholar]
- Skene, K.R. Life’s a gas: A thermodynamic theory of biological evolution. Entropy 2015, 17, 5522–5548. [Google Scholar] [CrossRef] [Green Version]
- Swenson, R.; Turvey, M.T. Thermodynamic Reasons for Perception—Action Cycles. Ecol. Psychol. 1991, 3, 317–348. [Google Scholar] [CrossRef]
- De Bari, B.; Dixon, J.A.; Kondepudi, D.; Kay, B. Oscillatory dynamics of an electrically driven dissipative structure. PLoS ONE 2019, 14, e0217305. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.K.; Hubler, A.H. Formation and structure of ramified charge transportation networks in an electromechanical system. Proc. Natl. Acad. Sci. USA 2005, 102, 536–540. [Google Scholar] [CrossRef] [Green Version]
- Martyushev, L.M.; Seleznev, V.D. Maximum entropy production principle in physics, chemistry, and biology. Phys. Rep. 2006, 1, 1–45. [Google Scholar] [CrossRef]
- Paltridge, G.W. Global dynamics and climate—A system of minimum entropy exchange. Q. J. R. Meteorol. Soc. 1975, 101, 475–484. [Google Scholar] [CrossRef]
- Ozawa, H.; Ohmura, A.; Lorenz, R.D.; Pujol, T. The second law of thermodynamics and the global climate system: A review of the maximum entropy production principle. Rev. Geophys. 2003, 41. [Google Scholar] [CrossRef] [Green Version]
- Chung, B.J.; Ortega, B.; Vaidya, A. Entropy production in a fluid-solid system far from thermodynamic equilibrium. Eur. Phys. J. E 2017, 40, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Endres, R.G. Entropy production selects nonequilibrium states in multistable systems. Sci. Rep. 2018, 7, 14437. [Google Scholar] [CrossRef] [PubMed]
- Zupanovic, P.; Brumen, M.; Jagodic, M.; Juretic, D. Bacterial chemotaxis and entropy production. Philos. Trans. R. Soc. B 2010, 365, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Vallino, J.J.; Huber, J.A. Using maximum entropy production to describe microbial biogeochemistry over time and space in a meromictic pond. Front. Environ. Sci. 2018, 6, 307. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.J.; Kay, B.A.; Kondepudi, D.; Dixon, J.A. Spontaneous interentity coordination in a dissipative structure. Ecol. Psychol. 2016, 28, 23–36. [Google Scholar] [CrossRef]
- Mathis, A.; Mamidanna, P.; Cury, K.M.; Abe, T.; Murthy, V.N.; Mathis, M.W.; Bethge, M. DeepLabCut: Markerless pose estimation of user-defined body parts with deep learning. Nat. Neurosci. 2018, 21, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Pikovsky, A.; Rosenblum, M.; Kurths, K. Synchronization: A Universal Concept in Nonlinear Sciences; Cambridge University Press: New York, NY, USA, 2001. [Google Scholar]
- De Bari, B.; Kondepudi, D.; Kay, B.A.; Dixon, J.A. Collective dissipative structures, force-flow reciprocity, and the foundations of perception-action mutuality. Ecol. Psychol. 2020. [Google Scholar] [CrossRef]
- Marsh, K.; Johnston, L.; Richardson, M.J.; Schmidt, R.C. Toward a radically embodied, embedded social psychology. Eur. J. Soc. Psychol. 2009, 39. [Google Scholar] [CrossRef]
- Marsh, K.; Richardson, M.J.; Baron, R.M.; Schmidt, R.C. Contrasting approaches to perceiving and acting with others. Ecol. Psychol. 2010, 18, 1–38. [Google Scholar] [CrossRef]
- Schmidt, R.C. Scaffolds for social meaning. Ecol. Psychol. 2007, 19, 137–151. [Google Scholar]
- Schmidt, R.C.; Carello, C.; Turvey, M.T. Phase transitions and critical fluctuations in the visual coordination of rhythmic movements between people. J. Exp. Psychol. Hum. Percept. Perform. 1990, 16, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Latash, M.L.; Shim, J.K.; Smilga, A.V.; Zatsiorsky, V.M. A Central Back-Coupling Hypothesis on the Organization of Motor Synergies: A Physical Metaphor and a Neural Model. Biol. Cybern. 2005, 92, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santello, M.; Soechting, J.F. Force synergies for multifingered grasping. Exp. Brain Res. 2000, 133, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Ramenzoni, V.C.; Riley, M.A.; Shockley, K.; Baker, A.A. Interpersonal and intrapersonal coordinative modes for joint and single task performance. Hum. Mov. Sci. 2012, 31, 1253–1267. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| s | High Coupling | Medium Coupling | Low Coupling |
|---|---|---|---|
| Bead 1 Constraints | xc = (−3, −1) | xc = (−4, −2) | xc = (−5, −3) |
| Bead 2 Constraints | xc = (1, 3) | xc = (2, 4) | xc = (3, 5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Bari, B.; Paxton, A.; Kondepudi, D.K.; Kay, B.A.; Dixon, J.A. Functional Interdependence in Coupled Dissipative Structures: Physical Foundations of Biological Coordination. Entropy 2021, 23, 614. https://doi.org/10.3390/e23050614
De Bari B, Paxton A, Kondepudi DK, Kay BA, Dixon JA. Functional Interdependence in Coupled Dissipative Structures: Physical Foundations of Biological Coordination. Entropy. 2021; 23(5):614. https://doi.org/10.3390/e23050614
Chicago/Turabian StyleDe Bari, Benjamin, Alexandra Paxton, Dilip K. Kondepudi, Bruce A. Kay, and James A. Dixon. 2021. "Functional Interdependence in Coupled Dissipative Structures: Physical Foundations of Biological Coordination" Entropy 23, no. 5: 614. https://doi.org/10.3390/e23050614
APA StyleDe Bari, B., Paxton, A., Kondepudi, D. K., Kay, B. A., & Dixon, J. A. (2021). Functional Interdependence in Coupled Dissipative Structures: Physical Foundations of Biological Coordination. Entropy, 23(5), 614. https://doi.org/10.3390/e23050614