Biosynthesis and Genetic Regulation of Proanthocyanidins in Plants

Abstract
:
Introduction
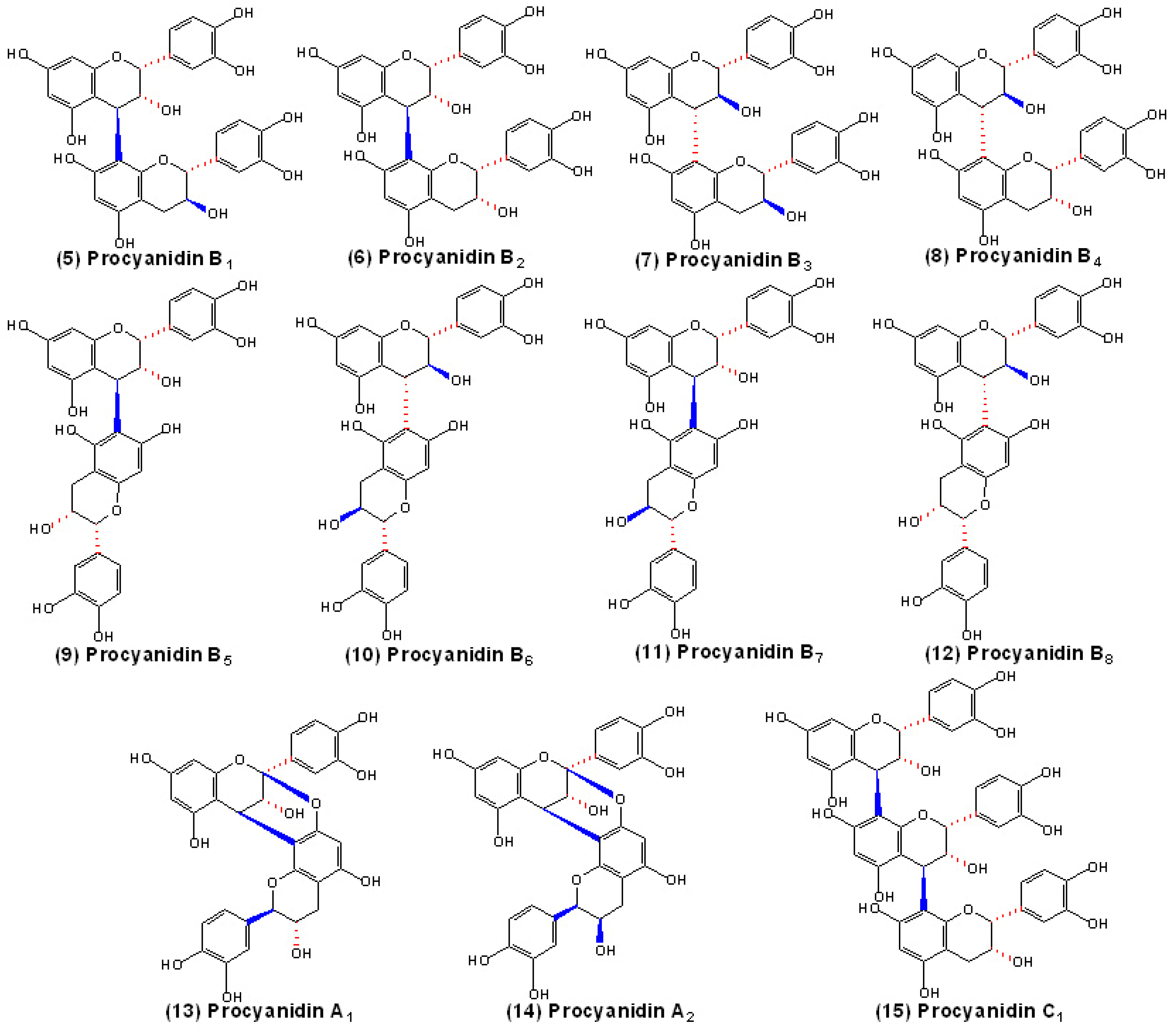
Structures of proanthocyanidins


General biosynthetic pathway of flavonoids

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure proteins | Plant species | ||
|---|---|---|---|
| Arabidopsis | Maize | Barley | |
| CHS | tt4 | c2 | |
| CHI | tt5 | chi | ant30 |
| F3’H | tt7 | pr1 | ant1, ant2, ant 5 |
| F3H | tt6 | fht1 | ant17, ant22 |
| DFR | tt3 | a1 | ant18 |
| ANS | ans/tt18 | a2 | |
| ANR | ban | ||
| Transporter | tt12 | ||

Transport factors of proanthocyanidin precursors
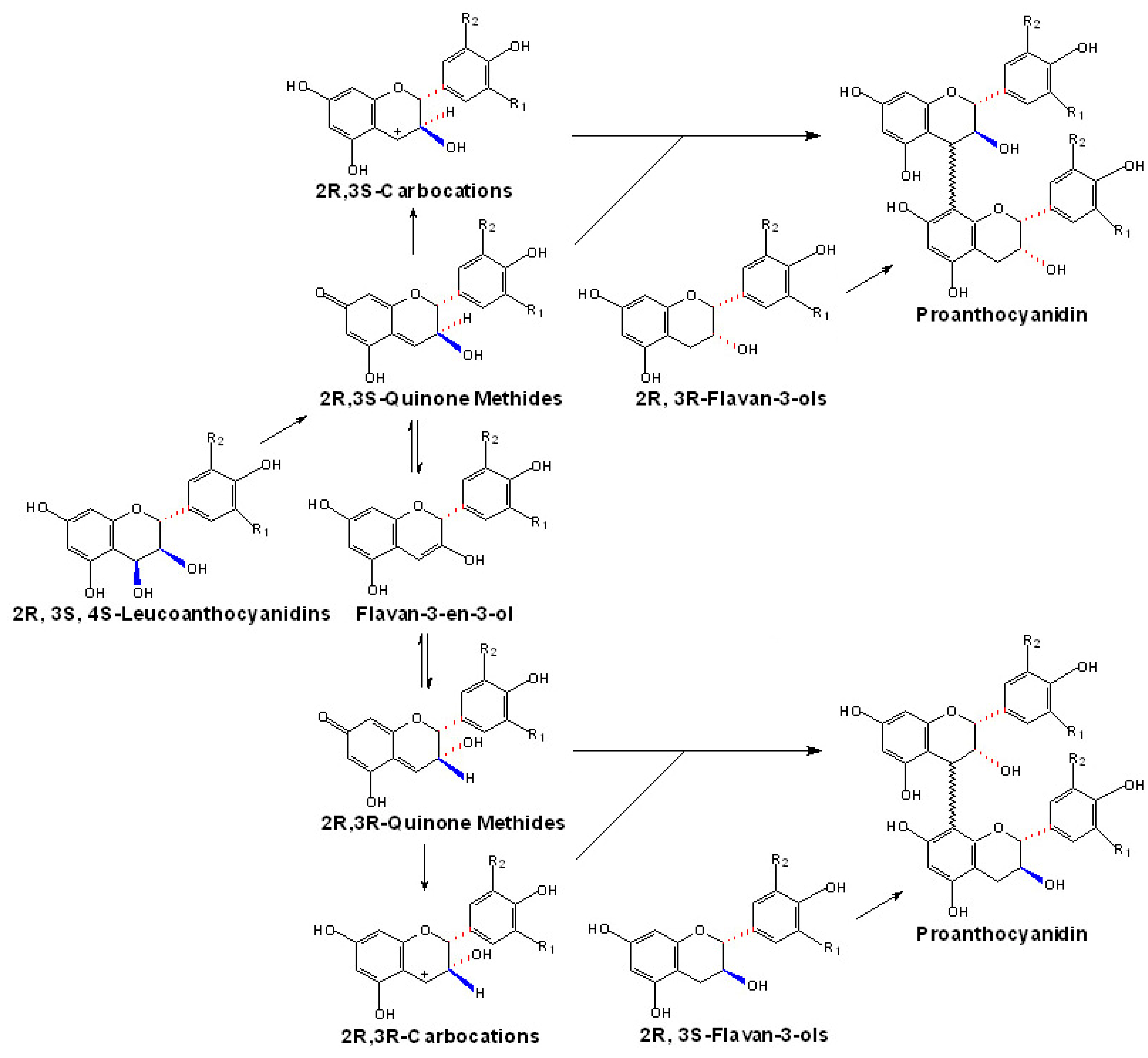
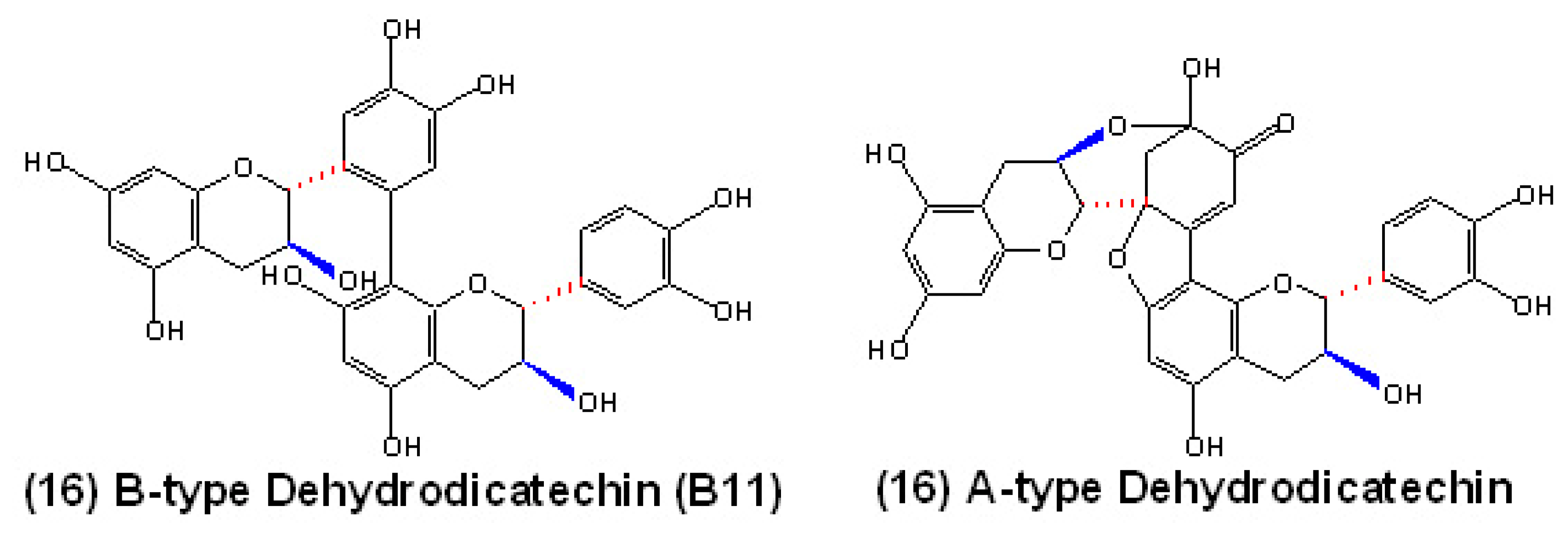
Potential polymerization mechanisms for proanthocyanidins



Regulation of proanthocyanidins biosynthesis
| Families | Genes | Locus | Gene regulated |
|---|---|---|---|
| MYC | TT8 | At4g09820 | BAN, DFR |
| GL3/EGL3 | At5g41315/At1g63650 | DFR | |
| MYB | TT2 | At5g35550 | BAN, DFR, ANS, TT12, AHA10, TT8, TTG2 |
| PAP1/PAP2 | At1g56650/At1g66390 | PAL,CHS, DFR, ANS, TT8 | |
| WD40 | TTG1 | At5g24520 | BAN, DFR, TT8, TTG2 |
| WRKY | TTG2 | At2g37260 | BAN, TT12 |
| MADS | TT16 | At5g23260 | BAN, TT2 |
| WIP | TT1 | At1g34790 | BAN |

Genetic manipulation of proanthocyanidin biosynthesis
Discussion and Conclusions
Acknowledgements
References and Notes
- Dixon, R.A.; Xie, D.Y.; Sharma, S.B. Proanthocyanidins - a final frontier in flavonoid research? New Phytol. 2005, 165, 9–28. [Google Scholar] [CrossRef]
- Bruyne, T.D.; Pieters, L.; Deelstra, H.; Vlietinck, A. Condensed vegetable tannins: Biodiversity in structure and biological activities. Biochem. Syst. Ecol. 1999, 27, 445–459. [Google Scholar] [CrossRef]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: a colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Feeny, P. Seasonal changes in oak leaf tannins and nutrients as a cause of spring feeding by winter moth caterpillars. Ecology 1970, 51, 565–581. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: from molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and tannin-like compounds – nature, occurrence, dietary intake and effects on nutrition and health. J. Sci. Food Agric. 2000, 80, 1094–1117. [Google Scholar] [CrossRef]
- Peleg, H.; Gacon, K.; Schlich, P.; Noble, A.C. Bitterness and astringency of flavan-3-ol monomers, dimers and trimers. J. Sci. Food Agric. 1999, 79, 1123–1128. [Google Scholar] [CrossRef]
- Prior, R.L.; Gu, L. Occurrence and biological significance of proanthocyanidins in the American diet. Phytochemistry 2005, 66, 2264–2280. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, B.; Wang, J.S.; Liu, Y.; Yu, L.M.; Jiang, Y.M. Immunomodulatory and anticancer activities of flavonoids extracted from litchi (Litchi chinensis Sonn.) pericarp. Int. Immunopharmacol. 2007, 7, 162–166. [Google Scholar] [CrossRef]
- Rao, L.J.M.; Yada, H.; Ono, H.; Ohnishi-Kameyama, M.; Yoshida, M. Occurrence of antioxidant and radical scavenging proanthocyanidins from the Indian minor spice nagkesar (Mammea longifolia planch and triana syn). Bioorg. Med. Chem. 2004, 12, 31–36. [Google Scholar] [CrossRef]
- Subarnas, A.; Wagner, H. Analgesic and anti-inflammatory activity of the proanthocyanidin shellegueain A from Polypodium feei METT. Phytomedicine 2000, 7, 401–405. [Google Scholar] [CrossRef]
- Sato, M.; Maulik, G.; Ray, P.S.; Bagchi, D.; Das, D.K. Cardioprotective effects of grape seed proanthocyanidin against ischemic reperfusion injury. J. Mol. Cell. Cardiol. 1999, 31, 1289–1297. [Google Scholar] [CrossRef]
- Sano, T.; Oda, E.; Yamashita, T.; Naemura, A.; Ijiri, Y.; Yamakoshi, J.; Yamamoto, J. Anti-thrombotic effect of proanthocyanidin, a purified ingredient of grape seed. Thromb. Res. 2005, 115, 115–121. [Google Scholar] [CrossRef]
- Sharma, S.D.; Meeran, S.M.; Katiyar, S.K. Dietary grape seed proanthocyanidins inhibit UVB-induced oxidative stress and activation of mitogenactivated protein kinases and nuclear factor-кB signaling in vivo SKH-1 hairless mice. Mol. Cancer. Ther. 2007, 6, 995–1005. [Google Scholar] [CrossRef]
- Yamakoshi, J.; Saito, M.; Kataoka, S.; Kikuchi, M. Safety evaluation of proanthocyanidin-rich extract from grape seeds. Food Chem. Toxicol. 2002, 40, 599–607. [Google Scholar] [CrossRef]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Zhang, Z.; Beecher, G.; Holden, J.; Haytowitz, D.; Prior, R.L. Liquid chromatographic/electrospray ionization mass spectrometric studies of proanthocyanidins in foods. J. Mass Spectrom. 2003, 38, 1272–1280. [Google Scholar] [CrossRef]
- Tarascou, I.; Barathieu, K.; Simon, C.; Ducasse, M.A.; André, Y.; Fouquet, E.; Dufourc, E.J.; Freitas, V.; Laguerre, M.; Pianet, I. A 3D structural and conformational study of procyanidin dimers in water and hydro-alcoholic media as viewed by NMR and molecular modeling. Magn. Reson. Chem. 2006, 44, 868–880. [Google Scholar] [CrossRef]
- Slade, D.; Ferreira, D.; Marais, J.P.J. Circular dichroism, a powerful tool for the assessment of absolute configuration of flavonoids. Phytochemistry 2005, 66, 2177–2215. [Google Scholar] [CrossRef]
- Matthews, S.; Mila, I.; Scalbert, A.; Pollet, B.; Lapierre, C.; Hervé du Penhoat, C.L.M.; Rolando, C.; Donnelly, D.M.X. Method for estimation of proanthocyanidins based on their acid depolymerization in the presence of nucleophiles. J. Agric. Food Chem. 1997, 45, 1195–1201. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Jones, G.P. Analysis of proanthocyanidin cleavage products following acid-catalysis in the presence of excess phloroglucinol. J. Agric. Food Chem. 2001, 49, 1740–1746. [Google Scholar] [CrossRef]
- Marles, M.A.S.; Ray, H.; Gruber, M.Y. New perspectives on proanthocyanidin biochemistry and molecular regulation. Phytochemistry 2003, 64, 367–383. [Google Scholar] [CrossRef]
- Xie, D.Y.; Dixon, R.A. Proanthocyanidin biosynthesis – still more questions than answers? Phytochemistry 2005, 66, 2127–2144. [Google Scholar] [CrossRef]
- Stafford, H.A. Proanthocyanidins and the lignin connection. Phytochemistry 1988, 27, 1–6. [Google Scholar] [CrossRef]
- Schofield, P.; Mbugua, D.M.; Pell, A.N. Analysis of condensed tannins: a review. Anim. Feed Sci. Tech. 2001, 91, 21–40. [Google Scholar] [CrossRef]
- Kelm, M.A.; Hammerstone, J.F.; Schmitz, H.H. Identification and quantitation of flavanols and proanthocyanidins in foods: How good are the datas? Clin. Dev. Immunol. 2005, 12, 35–41. [Google Scholar] [CrossRef]
- Tian, L.; Pang, Y.; Dixon, R.A. Biosynthesis and genetic engineering of proanthocyanidins and (iso)flavonoids. Phytochem. Rev. 2007, 1–21. [Google Scholar]
- Buzzini, P.; Turchetti, B.; Ieri, F.; Goretti, M.; Branda, E.; Mulinacci, N.; Romani, A. Catechins and proanthocyanidins: naturally occurring O-heterocycles with antimicrobial activity. TopHeterocycl. Chem. 2007, 10, 239–263. [Google Scholar]
- Rasmussen, S.E.; Frederiksen, H.; Krogholm, K.S.; Poulsen, L. Dietary proanthocyanidins: occurrence, dietary intake, bioavailability, and protection against cardiovascular disease. Mol. Nutr. Food Res. 2005, 49, 159–174. [Google Scholar] [CrossRef]
- Ferreira, D.; Li, X.C. Oligomeric proanthocyanidins: naturally occurring O-heterocycles. Nat. Prod. Rep. 2000, 17, 193–212. [Google Scholar] [CrossRef]
- Ferreira, D.; Slade, D. Oligomeric proanthocyanidins: naturally occurring O-heterocycles. Nat. Prod. Rep. 2002, 19, 517–541. [Google Scholar] [CrossRef]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Beecher, G.; Holden, J.; Haytowitz, D.; Prior, R.L. Screening of foods containing proanthocyanidins and their structural characterization using LC-MS/MS and thiolytic degradation. J. Agric. Food Chem. 2003, 51, 7513–7521. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P. Plant Phenolics; Hafner Publishing Company: New York, 1972; p. 254. [Google Scholar]
- Geiss, F.; Heinrich, M.; Hunkler, D.; Rimpler, H. Proanthocyanidins with (+)-epicatechin units from Byrsonima crassifolia bark. Phytochemistry 1995, 39, 635–643. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Kolodzieij, H.; Treutter, D. Procyanidin trimers possessing a doubly linked structure from Aesculus hippocastanum. Phytochemistry 1995, 38, 499–504. [Google Scholar] [CrossRef]
- Ferreira, D.; Bekker, R. Oligomeric proanthocyanidins: naturally occurring O-heterocycles. Nat. Prod. Rep. 1996, 13, 411–433. [Google Scholar] [CrossRef]
- Holton, T.A.; Brugliera, F.; Lester, D.R.; Tanaka, Y.; Hyland, C.D.; Menting, J.G.T.; Lu, C.Y.; Farcy, E.; Stevenson, T.W.; Cornish, E.C. Cloning and expression of cytochrome P450 genes controlling flower colour. Nature 1993, 366, 276–279. [Google Scholar] [CrossRef]
- Ayabe, S.; Akashi, T. Cytochrome P450s in flavonoid metabolism. Phytochem. Rev. 2006, 5, 271–282. [Google Scholar] [CrossRef]
- Malan, J.C.S.; Young, D.A.; Steynberg, J.P.; Ferreira, D. Oligomeric flavanoids. Part 10. Structure and synthesis of the first tetrahydropyrano[3,2-g]chromenes related to (4,6)-bis-(-)-fisetinidol profisetinidins. J. Chem. Soc., Perkin Trans. 1 1990, 227–234. [Google Scholar]
- Steynberg, P.J.; Steynberg, J.P.; Bezuidenhoudt, B.C.B.; Ferreira, D. Cleavage of the interflavanyl bond in 5-deoxy (A ring) proanthocyanidins. J. Chem. Soc., Chem. Commun. 1994, 31–32. [Google Scholar]
- Thompson, R.S.; Jacques, D.; Haslam, E.; Tanner, R.J.N. Plant proanthocyanidins -Part 1- Introduction: the isolation, structure and distribution in nature of plant procyanidins. J. Chem. Soc., Perkin Trans. 1 1982, 1387–1399. [Google Scholar]
- Hemingway, R.W.; Foo, L.Y.; Porter, L.J. Linkage isomerism in trimeric and polymeric 2,3-cis-procyanidins. J. Chem. Soc., Perkin Trans. 1 1982, 1209–1216. [Google Scholar] [CrossRef]
- Weinges, K.; Freundenberg, K. Condensed proanthocyanidins from cranberries and cola nuts. Chem. Commun. 1965, 2, 220–222. [Google Scholar]
- Guyot, S.; Marnet, N.; Drilleau, J. Thiolysis-HPLC characterization of apple procyanidins covering a large range of polymerization states. J. Agric. Food Chem. 2001, 49, 14–20. [Google Scholar] [CrossRef]
- Takahata, Y.; Ohnishi-Kameyama, M.; Furuta, S.; Takahashi, M.; Suda, I. Highly polymerized procyanidins in brown soybean seed coat with a high radical-scavenging activity. J. Agric. FoodChem. 2001, 49, 5843–5847. [Google Scholar] [CrossRef]
- Ferreira, D.; Guyot, S.; Marnet, N.; Delgadillo, I.; Renard, C.M.; Coimbra, M.A. Composition of phenolic compounds in a Portuguese pear (Pyrus communis L. var. S. Bartolomeu) and changes after sun-drying. J. Agric. Food Chem. 2002, 50, 4537–4544. [Google Scholar] [CrossRef]
- Prieur, C.; Rigaud, J.; Cheynier, V.; Moutounet, M. Oligomeric and polymeric procyanidins from grape seeds. Phytochemistry 1994, 36, 781–784. [Google Scholar] [CrossRef]
- Cheynier, V. Polyphenols in foods are more complex than often thought. Am. J. Clin. Nutr. 2005, 81, 223S–229S. [Google Scholar]
- Passos, C.P.; Cardoso, S.M.; Domingues, M.R.M.; Domingues, P.; Silva, C.M.; Coimbra, M.A. Evidence for galloylated type-A procyanidins in grape seeds. Food Chem. 2007, 105, 1457–1467. [Google Scholar] [CrossRef]
- Forrest, G.I.; Bendall, D.S. The distribution of polyphenols in the tea plant (Camellia sinensis L.). Biochem J. 1969, 113, 741–755. [Google Scholar]
- Lee, C.Y.; Jaworski, A. Phenolic compounds in white grapes grown in New York. Am. J. Enol. Vitic. 1987, 38, 277–281. [Google Scholar]
- Lee, C.Y.; Jaworski, A. Identification of some phenolics in white grapes. Am. J. Enol. Vitic. 1990, 41, 87–89. [Google Scholar]
- Achmadi, S.; Syahbirin, G.; Choong, E.T.; Hemingway, R.W. Catechin-3-O-rhamnoside chain extender units in polymeric procyanidins from mangrove bark. Phytochemistry 1994, 35, 217–219. [Google Scholar]
- Iida, N.; Inatomi, Y.; Murata, H.; Inada, A.; Murata, J.; Lang, F.A.; Matsuura, N.; Nakanishi, T. A new flavone xyloside and two new flavan-3-ol glucosides from Juniperus communis var. depressa. Chem. Biodivers. 2007, 4, 32–42. [Google Scholar] [CrossRef]
- Fletcher, A.C.; Porter, L.J.; Haslam, E.; Gupta, R.K. Plant proanthocyanidins. Part 3. Conformational and configurational studies of natural procyanidins. J. Chem. Soc., Perkin Trans. 1 1977, 1628–1637. [Google Scholar]
- Zucker, W.V. Tannins: does structure determine function? An ecological perspective. Am. Nat. 1983, 121, 335–365. [Google Scholar]
- Sarni-Manchado, P.; Cheynier, V.; Moutounet, M. nteractions of grape seed tannins with salivary proteins. J. Agric. Food Chem. 1999, 47, 42–47. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Powell, K.J. Polyphenol interactions with aluminium(III) and iron(III): their possible involvement in the podzolization process. Aust. J. Chem. 1985, 38, 879–888. [Google Scholar] [CrossRef]
- Weber, H.A.; Hodges, A.E.; Guthrie, J.R.; O’Brien, B.M.; Robaugh, D.; Clark, A.P.; Harris, R.K.; AlgaierL, J.W.; Smith, C.S. Comparison of proanthocyanidins in commercial antioxidants: grape seed and pine bark extracts. J. Agric. Food Chem. 2007, 55, 148–156. [Google Scholar] [CrossRef]
- Saito, K.; Yamazaki, M. Tansley review no. 138 Biochemistry and molecular biology of the late-stage of biosynthesis of anthocyanin: lessons from Perilla frutescens as a model plant. New Phyto. 2002, 155, 9–23. [Google Scholar] [CrossRef]
- Springob, K.; Nakajima, J.; Yamazaki, M.; Saito, K. Recent advances in the biosynthesis and accumulation of anthocyanins. Nat. Prod. Rep. 2003, 20, 288–303. [Google Scholar] [CrossRef]
- Abrahams, S.; Tanner, G.J.; Larkin, P.J.; Ashton, A.R. Identification and biochemical characterization of mutants in proanthocyanidin pathway in Arabidopsis. Plant Physiol 2002, 130, 561–576. [Google Scholar] [CrossRef]
- Cone, K.C. Anthocyanin synthesis in maize aleurone tissue. Plant Cell Monogr. 2007, 8, 121–139. [Google Scholar] [CrossRef]
- Jende-Strid, B. Genetic control of flavonoid biosynthesis in barley. Hereditas 1993, 119, 187–204. [Google Scholar] [CrossRef]
- Kreuzaler, F.; Hahlbrock, K. Enzymatic synthesis of aromatic compounds in higher plants. Formation of naringenin (5,7,4'-trihydroxyflavanone) from p-coumaroyl coenzyme A and malonyl coenzyme A. FEBS Lett. 1972, 28, 69–72. [Google Scholar] [CrossRef]
- Saslowsky, D.; Winkel-Shirley, B. Localization of flavonoid enzymes in Arabidopsis roots. Plant J. 2001, 27, 37–48. [Google Scholar] [CrossRef]
- Tian, L.; Wan, S.B.; Pan, Q.H.; Zheng, Y.J.; Huang, W.D. A novel plastid localization of chalcone synthase in developing grape berry. Plant Sci. 2008, 175, 431–436. [Google Scholar] [CrossRef]
- Cain, C.C.; Saslowsky, D.E.; Walker, R.A.; Shirley, B.W. Expression of chalcone synthase and chalcone isomerase proteins in Arabidopsis seedlings. Plant Mol. Biol. 1997, 35, 377–381. [Google Scholar] [CrossRef]
- Bednar, R.A.; Hadcock, J.R. Purification and Characterizationo of Chalcone Isomerase from Soybeans. J. Biol. Chem. 1988, 263, 9582–9588. [Google Scholar]
- Jez, J.M.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structure and mechanism of the evolutionarily unique plant enzyme chalcone isomerase. Nat. Struct. Biol. 2000, 7, 786–791. [Google Scholar] [CrossRef]
- Jez, J.M.; Noel, J.P. Reaction mechanism of chalcone isomerase. J. Biol.Chem. 2002, 277, 1361–1369. [Google Scholar] [CrossRef]
- Britsch, L.; Dedio, J.; Saedler, H.; Forkmann, G. Molecular characterization of flavanone 3β-hydroxylases. Consensus sequence, comparison with related enzymes and the role of conserved histidine residues. Eur. J. Biochem. 1993, 217, 745–754. [Google Scholar] [CrossRef]
- Lukačin, R.; Urbanke, C.; Gröning, I.; Matern, U. The monomeric polypeptide comprises the functional flavanone 3β-hydroxylase from Petunia hybrida. FEBS Lett. 2000, 467, 353–358. [Google Scholar] [CrossRef]
- Martens, S.; Teeri, T.; Forkmann, G. Heterologous expression of dihydroflavonol 4-reductases from various plants. FEBS Lett. 2002, 531, 453–458. [Google Scholar] [CrossRef]
- Petit, P.; Granier, T.; d’Estaintot, B.L.; Manigand, C.; Bathany, K.; Schmitter, J.M.; Lauvergeat, V.; Hamdi, S.; Gallois, B. Crystal structure of grape dihydroflavonol 4-reductase, a key enzyme in flavonoid biosynthesis. J. Mol. Biol. 2007, 368, 1345–1357. [Google Scholar] [CrossRef]
- Nakajima, J.; Tanaka, Y.; Yakushiji, H.; Saito, K. Reaction mechanism from leucoanthocyanidin to anthocyanidin 3-glucoside, a key reaction for coloring in anthocyanin biosynthesis. J. Biol. Chem. 2001, 276, 25797–25803. [Google Scholar] [CrossRef]
- Pelletier, M.K.; Murrell, J.R.; Shirley, B.W. Characterization of flavonol synthase and leucoanthocyanidin dioxygenase genes in Arabidopsis. Further evidence for differential regulation of “Early” and “Late” genes. Plant Physiol. 1997, 11, 1437–1445. [Google Scholar]
- Saito, K.; Kobayashi, M.; Gong, Z; Tanaka, Y.; Yamazaki, M. Direct evidence for anthocyanidin synthase as a 2-oxoglutarate-dependent oxygenase molecular cloning and functional expression of cDNA from a red forma of Perilla frutescens. Plant J. 1999, 17, 181–189. [Google Scholar] [CrossRef]
- Xie, D.Y.; Sharma, S.B.; Paiva, N.L.; Ferreira, D.; Dixon, R.A. Role of anthocyanidin reductase, encoded by BANYULS in plant flavonoid biosynthesis. Science 2003, 299, 396–399. [Google Scholar] [CrossRef]
- Xie, D.Y.; Sharma, S.B.; Dixon, R.A. Anthocyanidin reductases from Medicago truncatula and Arabidopsis thaliana. Arch. Biochem. Biophys. 2004, 422, 91–102. [Google Scholar] [CrossRef]
- Welford, R.W.D.; Turnbull, J.J.; Claridge, T.D.W.; Prescott, A.G.; Schofield, C.J. Evidence for oxidation at C-3 of the flavonoid C-ring during anthocyanin biosynthesis. Chem. Commun. 2001, 1828–1829. [Google Scholar]
- Wellmann, F.; Griesser, M.; Schwab, W.; Martens, S.; Eisenreich, W.; Matern, U.; Lukačin, R. Anthocyanidin synthase from Gerbera hybrida catalyzes the conversion of (+)-catechin to cyanidin and a novel procyanidin. FEBS Lett. 2006, 580, 1642–1648. [Google Scholar] [CrossRef]
- Tanner, G.J.; Francki, K.T.; Abrahams, S.; Watson, J.M.; Larkin, P.J.; Ashton, A.R. Proanthocyanidin Biosynthesis in Plants. Proanthocyanidin biosynthesis in plants. Purification of legume leucoanthocyanidin reductase and molecular cloning of its cDNA. J. Biol Chem. 2003, 278, 31647–31656. [Google Scholar]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef]
- Pang, Y.; Peel, G.J.; Wright, E.; Wang, Z.; Dixon, R.A. Early steps in proanthocyanidin biosynthesis in the model legume Medicago truncatula. Plant Physiol. 2007, 145, 601–615. [Google Scholar] [CrossRef]
- Ford, C.M.; Boss, P.K.; Høj, P.B. Cloning and characterization of Vitis vinifera UDP-glucose:flavonoid 3-O-glucosyltransferase, a homologue of the enzyme encoded by the maize Bronze-1 locus that may primarily serve to glucosylate anthocyanidins in vivo. J. Biol Chem. 1998, 273, 9224–9233. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, B.G.; Ko, J.H.; Lee, Y.; Hur, H.G.; Lim, Y.; Ahn, J.H. Molecular cloning, expression, and characterization of a flavonoid glycosyltransferase from Arabidopsis thalian. Plant Sci. 2006, 170, 897–903. [Google Scholar] [CrossRef]
- Baxter, I.R.; Young, J.C.; Armstrong, G.; Foster, N.; Bogenschutz, N.; Cordova, T.; Peer, W.A.; Hazen, S.P.; Murphy, A.S.; Harper, J.F. A plasma membrane H+-ATPase is required for the formation of proanthocyanidins in the seed coat endothelium of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2005, 102, 2649–2654. [Google Scholar] [CrossRef]
- Debeaujon, I.; Peeters, A.J.M.; Léon-Kloosterziel, K.M.; Koornneef, M. The TRANSPARENT TESTA12 gene of Arabidopsis encodes a multidrug secondary transporter-like protein required for flavonoid sequestration in vacuoles of seed coat endothelium. Plant Cell 2001, 13, 853–871. [Google Scholar] [CrossRef]
- Kitamura, S.; Shikazono, N.; Tanaka, A. TRANSPARENT TESTA19 is involved in the accumulation of both anthocyanins and proanthocyanidins in Arabidopsis. Plant J. 2004, 37, 104–114. [Google Scholar] [CrossRef]
- Marinova, K.; Pourcel, L.; Weder, B.; Schwarz, M.; Barron, D.; Routaboul, J.M.; Debeaujon, I.; Klein, M. The Arabidopsis MATE transporter TT12 acts as a vacuolar Flavonoid/H+-Antiporter active in proanthocyanidin-accumulating cells of the seed coat. Plant Cell 2007, 19, 2023–2038. [Google Scholar] [CrossRef]
- Marrs, K.A.; Alfenlto, M.R.; Lloyd, A.M.; Walbot, V. A glutathione S-transferase involved in vacuolar transfer encoded by the maize gene Bronze-2. Nature 1995, 375, 397–400. [Google Scholar] [CrossRef]
- Mueller, L.A.; Goodman, C.D.; Silady, R.A.; Walbot, V. AN9, a petunia glutathione S-transferase required for anthocyanin sequestration, is a flavonoid-binding protein. Plant Physiol 2000, 123, 1561–2000. [Google Scholar] [CrossRef]
- Ageorges, A.; Fernandez, L.; Vialet, S.; Merdinoglu, D.; Terrier, N.; Romieu, C. Four specific isogenes of the anthocyanin metabolic pathway are systematically co-expressed with the red colour of grape berries. Plant Sci. 2006, 170, 372–383. [Google Scholar] [CrossRef]
- Braidot, E.; Petrussa, E.; Bertolini, A.; Peresson, C.; Ermacora, P.; Loi, N.; Terdoslavich, M.; Passamonti, S.; Macrì, F.; Vianello, A. Evidence for a putative flavonoid translocator similar to mammalian bilitranslocase in grape berries (Vitis vinifera L.) during ripening. Planta 2008, 228, 203–213. [Google Scholar] [CrossRef]
- Botha, J.J.; Ferreira, D.; Roux, D.G. Synthesis of condensed tannins. Part 4. A direct biomimetic approach to [4,6]- and [4,8]- biflavonoids. J. Chem. Soc., Perkin Trans. 1 1981, 1235–1245. [Google Scholar] [CrossRef]
- Delcour, J.A.; Ferreira, D.; Roux, D.G. Synthesis of condensed tannins. Part 9. The condensation sequence of leucocyanidin with (+)-catechin and with the resultant procyanidins. J. Chem. Soc., Perkin Trans. 1 1983, 1711–1717. [Google Scholar] [CrossRef]
- Porter, L.J. The Flavonoids. In Flavans and Proanthocyanidins; Harborne, J.B., Ed.; Chapman & Hall: London, UK, 1988; pp. 21–62. [Google Scholar]
- Porter, L.J. The Flavonoids: Advances in Research Since 1986. In Flavans and Proanthocyanidins; Harborne, J.B., Ed.; Chapman& Hall: London, UK, 1993; pp. 23–55. [Google Scholar]
- Hemingway, R.W.; Laks, P.E. Condensed tannins: a proposed route to 2R,3R-(2,3-cis)-proanthocyanidins. J. Chem. Soc., Chem. Commun. 1985, 746–747. [Google Scholar] [CrossRef]
- Creasey, L.L.; Swain, T. Structure of condensed tannins. Nature 1965, 208, 151–153. [Google Scholar] [CrossRef]
- Vaughn, K.C.; Lax, A.R.; Duke, S.O. Polyphenol oxidase: the chloroplast enzyme with no established function. Physiol. Plant. 1988, 72, 659–665. [Google Scholar] [CrossRef]
- Ono, E.; Hatayama, M.; Isono, Y.; Sato, T.; Watanabe, R.; Yonekura-Sakakibara, K.; Fukuchi-Mizutani, M.; Tanaka, Y.; Kusumi, T.; Nishino, T.; Nakayama, T. Localization of a flavonoid biosynthetic polyphenol oxidase in vacuoles. Plant J. 2006, 45, 133–143. [Google Scholar] [CrossRef]
- Guyot, S.; Vercauteren, J.; Cheynier, V. Structural determination of colourless and yellow dimers resulting from (+)-catechin coupling catalyzed by grape polyphenoloxidase. Phytochemistry 1996, 42, 1279–1288. [Google Scholar] [CrossRef]
- Oszmianski, J.; Lee, C.Y. Enzymatic oxidative reaction of catechin and chlorogenic acid in a model system. J. Agric. Food Chem. 1990, 38, 1202–1204. [Google Scholar] [CrossRef]
- Oszmianski, J.; Cheynier, V.; Moutounet, M. Iron-catalyzed oxidation of (+)-catechin in model systems. J. Agric. Food Chem. 1996, 44, 1712–1715. [Google Scholar] [CrossRef]
- Sun, W.; Miller, J.M. Tandem mass spectrometry of the B-type procyanidins in wine and B-type dehydrodicatechins in an autoxidation mixture of (+)-catechin and (-)-epicatechin. J. Mass Spectrom. 2003, 38, 438–446. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 Encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef]
- Vidal, S.; Cartalade, D.; Souquet, J.M.; Fulcrand, H.; Cheynier, V. Changes in proanthocyanidin chain length in winelike model solutions. J. Agric. Food Chem. 2002, 50, 2261–2266. [Google Scholar] [CrossRef]
- Remy-Tanneau, S.; Guernevé, C.L.; Meudec, E.; Cheynier, V. Characterization of a colorless anthocyanin-flavan-3-ol dimer containing both carbon-carbon and ether interflavanoid linkages by NMR and mass spectrometry. J. Agric. Food Chem. 2003, 51, 3592–3597. [Google Scholar] [CrossRef]
- Yokotsuka, K.; Singleton, V.L. Disappearance of anthocyanins as grape juice is prepared and oxidized with PPO and PPO substrates. Am. J. Enol. Vitic. 1997, 48, 13–25. [Google Scholar]
- Escribano-Bailon, T.; Dangles, O.; Brouillard, R. Coupling reactions between flavylium ions and catechin. Phytochemistry 1996, 41, 1583–1592. [Google Scholar] [CrossRef]
- Hemingway, R.W.; Laks, P.E. Condensed tannins: a proposed route to 2R,3R-(2,3-cis)-proanthocyanidins. J. Chem. Soc., Chem. Commun. 1985, 746–747. [Google Scholar] [CrossRef]
- Nesi, N.; Debeaujon, I.; Jond, C.; Pelletier, G.; Caboche, M.; Lepiniec, L. The TT8 gene encodes a basic helix-loop-helix domain protein required for expression of DFR and BAN genes in Arabidopsis Siliques. Plant Cell 2000, 12, 1863–1878. [Google Scholar] [CrossRef]
- Ramsay, N.A.; Walker, A.R.; Mooney, M.; Gray, J.C. Two basic-helix-loop-helix genes (MYC-146 and GL3) from Arabidopsis can activate anthocyanin biosynthesis in a white-flowered Matthiola incana mutant. Plant Mol. Biol. 2003, 52, 679–688. [Google Scholar] [CrossRef]
- Nesi, N.; Jond, C.; Debeaujon, I.; Caboche, M.; Lepiniec, L. The Arabidopsis TT2 gene encodes an R2R3 MYB domain protein that acts as a key determinant for proanthocyanidin accumulation in developing seed. Plant Cell 2001, 13, 2099–2114. [Google Scholar]
- Borevitz, J.; Xia, Y.; Blount, J.W.; Dixon, R.A.; Lamb, C. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant Cell 2000, 12, 2383–2393. [Google Scholar] [CrossRef]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The TRANSPARENT TESTA GLABRA1 locus, which regulates trichome differentiation and anthocyanin biosynthesis in Arabidopsis, encodes a WD40 repeat protein. Plant Cell 1999, 11, 1337–1349. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. TRANSPARENT TESTA GLABRA2, a trichome and seed coat development gene of Arabidopsis, encodes a WRKY transcription factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef]
- Ishida, T.; Hattori, S.; Sano, R.; Inoue, K.; Shirano, Y.; Hayashi, H.; Shibata, D.; Sato, S.; Kato, T.; Tabata, S.; Okada, K.; Wadaa, T. Arabidopsis TRANSPARENT TESTA GLABRA2 is directly regulated by R2R3 MYB transcription factors and is involved in regulation of GLABRA2 transcription in epidermal differentiation. Plant Cell 2007, 19, 2531–2543. [Google Scholar] [CrossRef]
- Nesi, N.; Debeaujon, I.; Jond, C.; Stewart, A.J.; Jenkins, G.I.; Caboche, M.; Lepiniec, L. The TRANSPARENT TESTA16 locus encodes the Arabidopsis bisister MADS domain protein and is required for proper development and pigmentation of the seed coat. Plant Cell 2002, 14, 2463–2479. [Google Scholar] [CrossRef]
- Lalusin, A.G.; Nishita, K.; Kim, S.H.; Ohta, M.; Fujimura, T. A new MADS-box gene (IbMADS10) from sweet potato (Ipomoea batatas (L.) Lam) is involved in the accumulation of anthocyanin. Mol. Gen. Genomics. 2006, 275, 44–54. [Google Scholar] [CrossRef]
- Sagasser, M.; Lu, G.H.; Hahlbrock, K.; Weisshaar, B.A. A. thaliana TRANSPARENT TESTA 1 is involved in seed coat development and defines the WIP subfamily of plant zinc finger proteins. Gene. Dev. 2001, 16, 138–149. [Google Scholar]
- Kubo, H.; Kishi, M.; Goto, K. Expression analysis of ANTHOCYANINLESS2 gene in Arabidopsis. Plant Sci. 2008. [Google Scholar] [CrossRef]
- Skirycz, A.; Jozefczuk, S.; Stobiecki, M.; Muth, D.; Zanor, M.I.; Witt, I.; Mueller-Roeber, B. Transcription factor AtDOF4;2 affects phenylpropanoid metabolism in Arabidopsis thaliana. NewPhytol. 2007, 175, 425–438. [Google Scholar]
- Zhang, F.; Gonzalez, A.; Zhao, M.; Payne, C.T.; Lloyd, A. A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development 2003, 130, 4859–4869. [Google Scholar] [CrossRef]
- Baudry, A.; Caboche, M.; Lepiniec, L. TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant J. 2006, 46, 768–779. [Google Scholar] [CrossRef]
- Dubos, C.; Le Gourrierec, J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J.-M.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. 2008, 55, 940–953. [Google Scholar] [CrossRef]
- Zimmermann, I.M.; Heim, M.A.; Weisshaar, B.; Uhrig, J.F. Comprehensive identification of Arabidopsis thaliana MYB transcription factors interacting with R/B-like BHLH proteins. Plant J. 2004, 40, 22–34. [Google Scholar] [CrossRef]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Tonelli, C.; Weisshaar, B.; Martin, C. Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef]
- Chen, Z.-H.; Nimmo, G.A.; Jenkins, G.I.; Nimmo, H.G. BHLH32 modulates several biochemical and morphological processes that respond to Pi starvation in Arabidopsis. Biochem. J. 2007, 405, 191–198. [Google Scholar]
- Broun, P. Transcriptional control of flavonoid biosynthesis: a complex network of conserved regulators involved in multiple aspects of differentiation in Arabidopsis. Curr. Opin. Plant Biol. 2005, 8, 272–279. [Google Scholar] [CrossRef]
- Endt, D.V.; Kijne, J.W.; Memelink, J. Transcription factors controlling plant secondary metabolism: what regulates the regulators? Phytochemistry 2002, 61, 107–114. [Google Scholar] [CrossRef]
- Devic, M.; Guilleminot, J.; Debeaujon, I.; Bechtold, N.; Bensaude, E.; Koornneef, M.; Pelletier, G.; Delseny, M. The BANYULS gene encodes a DFR-like protein and is a marker of early seed coat development. Plant J. 1999, 19, 387–398. [Google Scholar] [CrossRef]
- Debeaujon, I.; Nesi, N.; Perez, P.; Devic, M.; Grandjean, O.; Caboche, M.; Lepiniec, L. Proanthocyanidin-accumulating cells in Arabidopsis testa: regulation of differentiation and role in seed development. Plant Cell 2003, 15, 2514–2531. [Google Scholar] [CrossRef]
- Cominelli, E.; Gusmaroli, G.; Allegra, D.; Galbiati, M.; Wade, H.K.; Jenkins, G.I.; Tonelli, C. Expression analysis of anthocyanin regulatory genes in response to different light qualities in Arabidopsis thaliana. J. Plant Physiol. 2008, 165, 886–894. [Google Scholar] [CrossRef]
- Peltonen, P.A.; Vapaavuori, E.; Julkunen-Tiitto, R. Accumulation of phenolic compounds in birch leaves is changed by elevated carbon dioxide and ozone. Global Change Biol. 2005, 11, 1305–1324. [Google Scholar] [CrossRef]
- Mori, K.; Sugaya, S.; Gemma, H. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic. 2005, 105, 319–330. [Google Scholar] [CrossRef]
- Ubi, B.E.; Honda, C.; Bessho, H.; Kondo, S.; Wada, M.; Kobayashi, S.; Moriguchi, T. Expression analysis of anthocyanin biosynthetic genes in apple skin: effect of UV-B and temperature. Plant Sci. 2006, 170, 571–578. [Google Scholar] [CrossRef]
- Jeong, S.T.; Goto-Yamamoto, N.; Kobayashi, S.; Esaka, M. Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skins. Plant Sci. 2004, 167, 247–252. [Google Scholar] [CrossRef]
- El-Kereamy, A.; Chervin, C.; Roustan, J.P.; Cheynier, V.; Souquet, J.M.; Moutounet, M.; Raynal, J.; Ford, C.; Latché, A.; Pech, J.C.; Bouzayen, M. Exogenous ethylene stimulates the long-term expression of genes related to anthocyanin biosynthesis in grape berries. Physiol. Plant. 2003, 119, 175–182. [Google Scholar] [CrossRef]
- Punyasiri, P.A..; Tanner, G.J.; Abeysinghe, I.S.B.; Kumar, V.; Campbell, P.M.; Pradeepa, N.H.L. Exobasidium vexans infection of Camellia sinensis increased 2,3-cis isomerisation and gallate esterification of proanthocyanidins. Phytochemistry 2004, 65, 2987–2994. [Google Scholar] [CrossRef]
- Kortekamp, A. Expression analysis of defence-related genes in grapevine leaves after inoculation with a host and a non-host pathogen. Plant Physiol. Biochem. 2006, 44, 58–67. [Google Scholar] [CrossRef]
- Hirner, A.A.; Veit, S.; Seitz, H.U. Regulation of anthocyanin biosynthesis in UV-A-irradiated cell cultures of carrot and in organs of intact carrot plants. Plant Sci. 2001, 161, 315–322. [Google Scholar] [CrossRef]
- Ryan, K.G.; Swinny, E.E.; Markham, K.R.; Winefield, C. Flavonoid gene expression and UV photoprotection in transgenic and mutant Petunia leaves. Phytochemistry 2002, 59, 23–32. [Google Scholar] [CrossRef]
- Lea, U.S.; Slimestad, R.; Smedvig, P.; Lillo, C. Nitrogen deficiency enhances expression of specific MYB and bHLH transcription factors and accumulation of end products in the flavonoid pathway. Planta 2007, 225, 1245–1253. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Matthews, M.A.; Gaspero, G.D.; Gambetta, G.A. Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta 2007, 227, 101–112. [Google Scholar] [CrossRef]
- Taylor, L.P.; Briggs, W.R. Genetic Regulation and Photocontrol of Anthocyanin Accumulation in Maize Seedlings. Plant Cell 1990, 2, 115–127. [Google Scholar] [CrossRef]
- Hartmann, U.; Sagasser, M.; Mehrtens, F.; Stracke, R.; Weisshaar, B. Differential combinatorial interactions of cis-acting elements recognized by R2R3-MYB, BZIP, and BHLH factors control light-responsive and tissue-specific activation of phenylpropanoid biosynthesis genes. Plant Mol. Biol. 2005, 57, 155–171. [Google Scholar] [CrossRef]
- Carey, C.C.; Strahle, J.T.; Selinger, D.A.; Chandler, V.L. Mutations in the pale aleurone color1 regulatory gene of the Zea mays anthocyanin pathway have distinct phenotypes relative to the functionally similar TRANSPARENT TESTA GLABRA1 gene in Arabidopsis thaliana. Plant Cell 2004, 16, 450–464. [Google Scholar] [CrossRef]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Association of VvmybA1 gene expression with anthocyanin production in grape (Vitis vinifera) skin-color mutants. J. Japan. Soc. Hort. Sci. 2005, 74, 196–203. [Google Scholar] [CrossRef]
- Walker, A.R.; Lee, E.; Bogs, J.; McDavid, D.A.J.; Thomas, M.R.; Robinson, S.P. White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. 2007, 49, 772–785. [Google Scholar] [CrossRef]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb-related genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis. Planta 2002, 215, 924–933. [Google Scholar] [CrossRef]
- Deluc, L.; Barrieu, F.; Marchive, C.; Lauvergeat, V.; Decendit, A.; Richard, T.; Carde, J.P.; Mérillon, J.M.; Hamdi, S. Characterization of a grapevine R2R3-MYB transcription factor that regulates the phenylpropanoid pathway. Plant Physiol. 2006, 140, 499–511. [Google Scholar] [CrossRef]
- Goplen, B.P.; Howarth, R.E.; Sarkar, S.K.; Lesins, K. A search for condensed tannins in annual and perennial species of Medicago, Trigonella, and Onobrychis. Crop Sci. 1980, 20, 801–804. [Google Scholar] [CrossRef]
- Lees, G.L. Condensed tannins in some forage legumes: their role in the prevention of ruminant pasture bloat. Basic Life Sci. 1992, 59, 915–934. [Google Scholar]
- Robbins, C.T.; Mole, S.; Hagerman, A.E.; Hanley, T.A. Role of tannins in defending plants against ruminants: reduction in dry matter digestion? Ecologl. 1987, 68, 1606–1615. [Google Scholar] [CrossRef]
- Sarwar, G.; Bell, J.M.; Sharby, T.F.; Jones, J.D. Nutritional evaluation of meals and meal fractions derived from rape and mustard seed. Can. J. Anim. Sci. 1981, 61, 719–733. [Google Scholar] [CrossRef]
- Ray, H.; Yu, M.; Auser, P.; Blahut-Beatty, L.; Mckersie, B; Bowley, S.; Westcott, N.; Coulman, B.; Lloyd, A.; Gruber, M.Y. Expression of anthocyanins and proanthocyanidins after transformation of alfalfa with maize Lc. Plant Physiology 2003, 132, 1448–1463. [Google Scholar] [CrossRef]
- Wang, Y.; Frutos, P.; Gruber, M.Y.; Ray, H.; McAllister, T.A. In vitro ruminal digestion of anthocyanidin-containing alfalfa transformed with the maize Lc regulatory gene. Can. J. Anim. Sci. 2006, 86, 1119–1130. [Google Scholar]
- Carron, T.R.; Robbins, M.P.; Morris, P. Genetic modification of condensed tannin biosynthesis in Lotus corniculatus. 1. Heterologous antisense dihydroflavonol reductase down-regulates tannin accumulation in ‘hairy root’ cultures. Theor. Appl. Genet. 1994, 87, 1006–1015. [Google Scholar]
- Bavage, A.D.; Davies, I.G.; Robbins, M.P.; Morris, P. Expression of an Antirrhinum dihydroflavonol reductase gene results inchanges in condensed tannin structure and accumulation in root cultures of Lotus corniculatus (bird’s foot trefoil). Plant Mol. Biol. 1997, 35, 443–458. [Google Scholar] [CrossRef]
- Robbins, M.P.; Paolocci, F.; Hughes, J.W.; Turchetti, V.; Allison, G.; Arcioni, S.; Morris, P.; Damiani, F. Sn, a maize bHLH gene, modulates anthocyanin and condensed tannin pathways in Lotus corniculatus. J. Exp. Bot. 2003, 54, 239–248. [Google Scholar] [CrossRef]
- Paolocci, F.; Bovone, T.; Tosti, N.; Arcioni, S.; Damiani, F. Light and an exogenous transcription factor qualitatively and quantitatively affect the biosynthetic pathway of condensed tannins in Lotus corniculatus leaves. J. Exp. Bot. 2005, 56, 1093–1103. [Google Scholar] [CrossRef]
- Paolocci, F.; Robbins, M.P.; Madeo, L.; Arcioni, S.; Martens, S.; Damiani, F. Ectopic expression of a basic helix-loop-helix gene transactivates parallel pathways of proanthocyanidin biosynthesis. Structure, expression, analysis, and genetic control of leucoanthocyanidin 4-reductase and anthocyanidin reductase genes in Lotus corniculatus. Plant Physiol. 2007, 143, 504–516. [Google Scholar]
- Sharma, S.B.; Dixon, R.A. Metabolic engineering of proanthocyanidins by ectopic expression of transcription factors in Arabidopsis thaliana. Plant J. 2005, 44, 62–75. [Google Scholar] [CrossRef]
- Xie, D.Y.; Sharma, S.B.; Wright, E.; Wang, Z.Y.; Dixon, R.A. Metabolic engineering of proanthocyanidins through co-expression of anthocyanidin reductase and the PAP1 MYB transcription factor. Plant J. 2006, 45, 895–907. [Google Scholar] [CrossRef]
- Abrahams, S.; Tanner, G.J.; Larkin, P.J.; Ashton, A.R. Identification and biochemical characterization of mutants in proanthocyanidin pathway in Arabidopsis. Plant Physiol. 2002, 130, 561–576. [Google Scholar] [CrossRef]
- Tian, L.; Kong, W.F.; Pan, Q.H.; Zhan, J.C.; Wen, P.F.; Chen, J.Y.; Wan, S.B.; Huang, W.D. Expression of the chalcone synthase gene from grape and preparation of an anti-CHS antibody. Protein Expres. Purif. 2006, 50, 223–228. [Google Scholar] [CrossRef]
- Hunt, A.G.; Xu, R.; Addepalli, B.; Rao, S.; Forbes, K.P.; Meeks, L.R.; Xing, D.; Mo, M.; Zhao, H.; Bandyopadhyay, A.; Dampanaboina, L.; Marion, A.; Von Lanken, C.; Li, Q.Q. Arabidopsis mRNA polyadenylation machinery: Comprehensive analysis of protein-protein interactions and gene expression profiling. BMC Genomic. 2008, 9. Article number 220. [Google Scholar]
- Sample Availability: Not available.
© 2008 by the authors. Licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
He, F.; Pan, Q.-H.; Shi, Y.; Duan, C.-Q. Biosynthesis and Genetic Regulation of Proanthocyanidins in Plants. Molecules 2008, 13, 2674-2703. https://doi.org/10.3390/molecules13102674
He F, Pan Q-H, Shi Y, Duan C-Q. Biosynthesis and Genetic Regulation of Proanthocyanidins in Plants. Molecules. 2008; 13(10):2674-2703. https://doi.org/10.3390/molecules13102674
Chicago/Turabian StyleHe, Fei, Qiu-Hong Pan, Ying Shi, and Chang-Qing Duan. 2008. "Biosynthesis and Genetic Regulation of Proanthocyanidins in Plants" Molecules 13, no. 10: 2674-2703. https://doi.org/10.3390/molecules13102674
APA StyleHe, F., Pan, Q. -H., Shi, Y., & Duan, C. -Q. (2008). Biosynthesis and Genetic Regulation of Proanthocyanidins in Plants. Molecules, 13(10), 2674-2703. https://doi.org/10.3390/molecules13102674